The Acipenseriformes (Chondrostei) represent a group of primitive, ray-finned fish, comprising Acipenseridae (sturgeons) and Polyodontidae (paddlefishes) (Bemis et al., 1997; Birstein et al., 1997). They are often referred to as ‘living fossils’ of the actinopterygian lineage and have distinctive morphologic and/or developmental attributes (Billard and Lecointre, 2001). Furthermore, some sturgeon species exhibit repeated rounds of genome duplication, resulting in diversification into different ploidy levels among sturgeon species (Blacklidge and Bidwell, 1993; Kim et al., 2005). Based on these unique and interesting characteristics, this extant primitive fish group is regarded as a useful model for evolutionary genetics and genomic studies (Cho et al., 2007; Kim et al., 2009; Akbarzadeh et al., 2011).
In addition to the interest in their evolutionary aspects, sturgeons have long been recognized as very valuable natural resources for fisheries in Northern Hemisphere countries, particularly with regard to caviar production. However, over the last few decades, wild sturgeon stocks have faced considerable threats to their natural habitats as a result of various anthropogenic and industrial activities (Pikitch et al., 2005). Currently, all extant sturgeon species are considered to be ‘critically endangered’ and are included in the Convention on International Trade in Endangered Species (CITES) list, which applies to global trading. Thus, sustainable, aquaculture-based production has become mandatory for the commercial exploitation of sturgeons and their products (Karpinsky, 2010; Webb and Doroshov, 2011).
In Korea, the introduction of sturgeon species into the aquaculture domain was first noted in 1999 for the Siberian sturgeon
>
Broodfish and hormonal artificial spawning
The first experiment for the morphologic staging of embryonic development was performed in March 2011, and this experiment was replicated in March 2012. The broodfish used for artificial spawning and gamete collection were individuals that were produced in 2003. The broodfish were maintained at ambient temperature (range, 12 to 20℃) at Dinoville Aquafarm Inc. (Hamyang, Korea). Fish were sexed by external gonad biopsy at 6 years of age. Out of six (in 2011) or four (in 2012) female candidates, four (average body weight [BW], 14 ± 2.6 kg) and three individuals (13 ± 4.6 kg BW) were selected for hormonal induction of spawning in 2011 and 2012, respectively. When necessary, polarization index (PI) observations using catheterized oocytes were conducted (PI scores < 0.1) (Van Eenennaam et al., 1996). In addition, four males each for 2011 (7 ± 2.7 kg BW) and 2012 (7 ± 3.1 kg BW) were selected based on the presence of milt, which was obtained by either palpation of the abdomen or catheterization using a silicon tube-connected syringe into the genital duct. The selected females were administered the primary intramuscular injection of the luteinizing hormone-releasing hormone analogue des- Gly10, [D-Ala6] LH-RH ethylamide (LHRHa; Syndel Laboratories Ltd., Qualicum Beach, BC, Canada) at 10-15 μg/kg BW. At 12 h after the primary injection, each female was injected again with LHRHa at 90-135 μg/kg BW as a resolving dose. The male broodfish were injected with LHRHa at 100 μg/kg BW. The temperature of the water during the induced spawning was adjusted to 15.0 ± 0.5℃ in both years. After injection, the fish were incubated in individual spawning tanks until a low number of ovulated eggs was released, which usually occurred around 36 h after the second injection of hormone.
>
Artificial insemination and egg incubation
When ovulation followed by spawning was identified, milt was collected from the males using a silicon tube-connected aspirator and stored at 2-4℃ until used. After milt collection, the eggs were removed from the spawned females by hand stripping and Caesarean section. The eggs were mixed with milt (diluted 1:200 with water) for 150 s, and then rinsed with fresh water. To remove the adhesiveness of the fertilized eggs, the eggs were mixed continuously with Fuller’s earth (Sigma- Aldrich, St. Louis, MO, USA) and washed with fresh water several times for 30 min. The fertilized eggs were placed in McDonald incubation jars until hatched. Dead embryos were removed at 6-h intervals. The water temperature was maintained at 18.0 ± 0.5℃ until hatching.
>
Sampling of biological specimens for developmental staging
In 2011, the development of embryos from two mating pairs was examined. Based on a modified version of the developmental stages defined for Russian sturgeon,
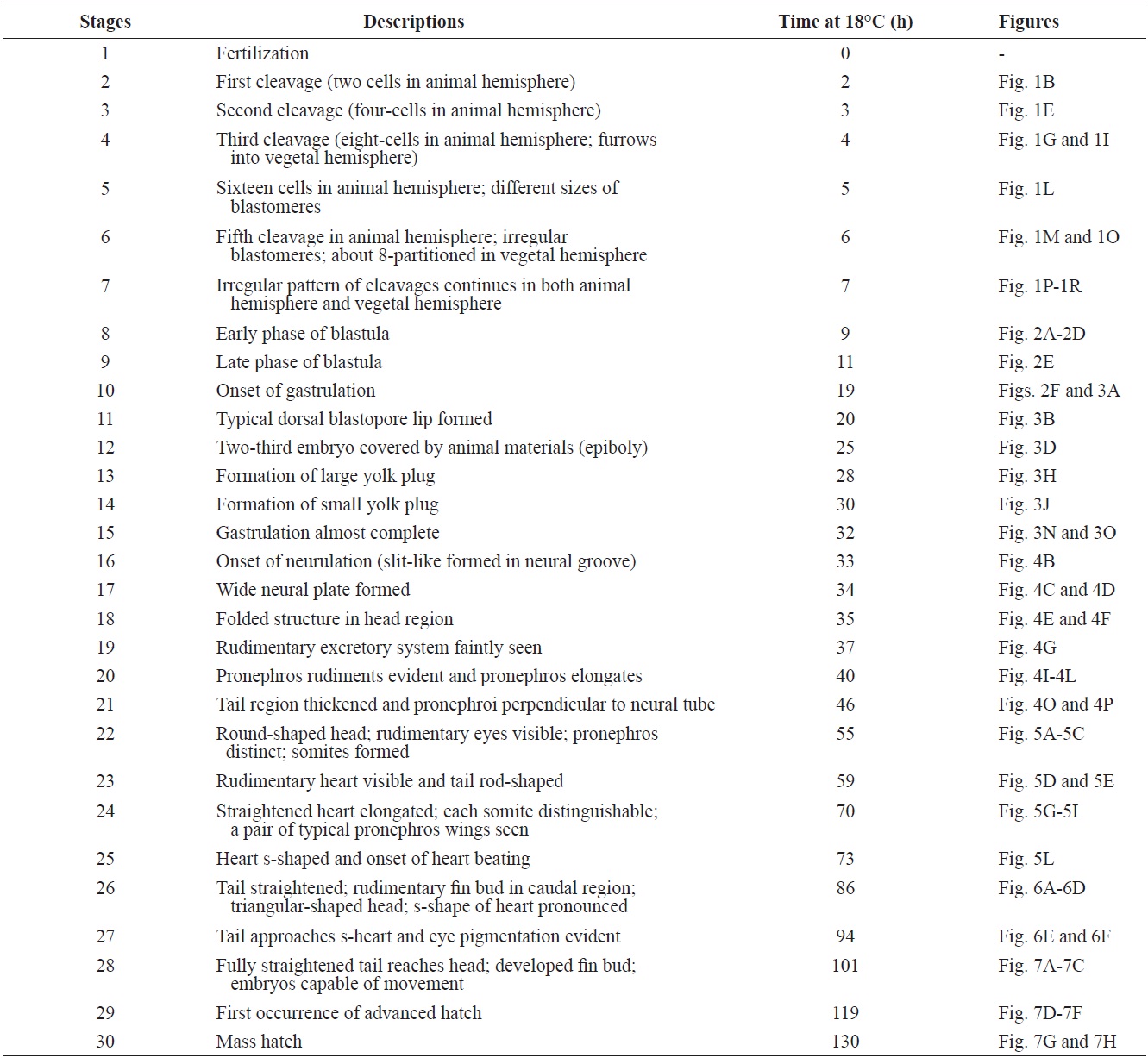
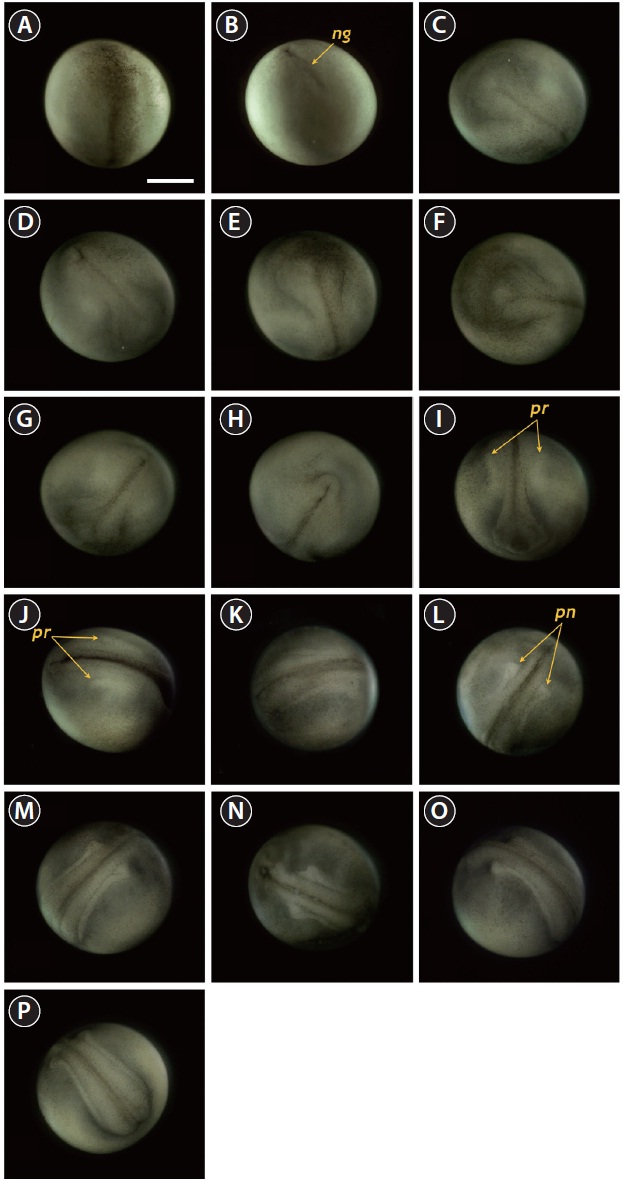
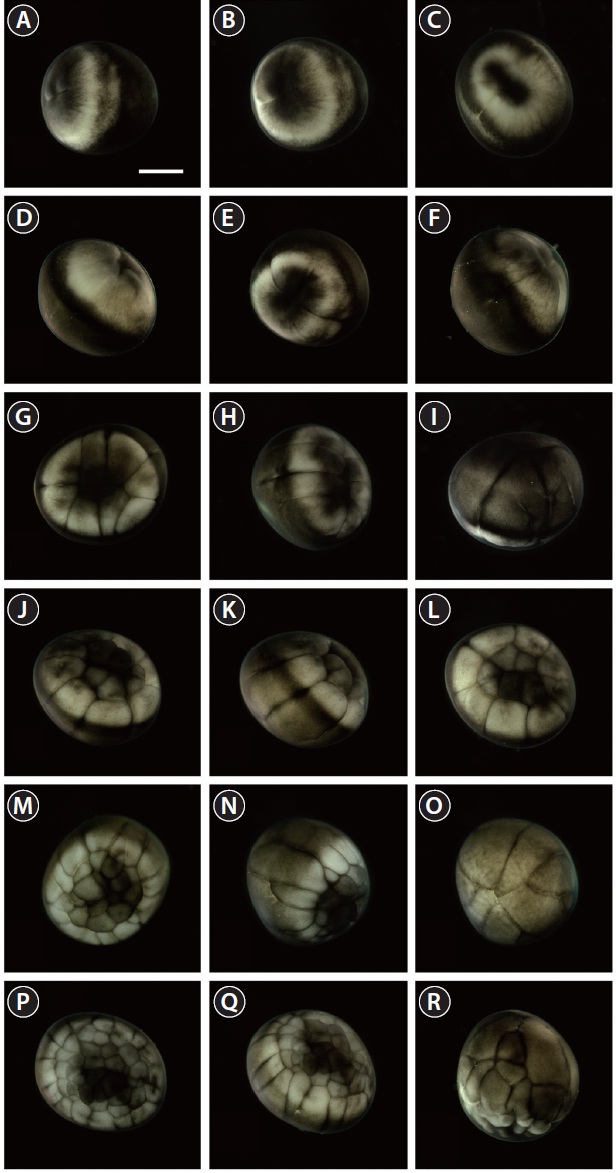
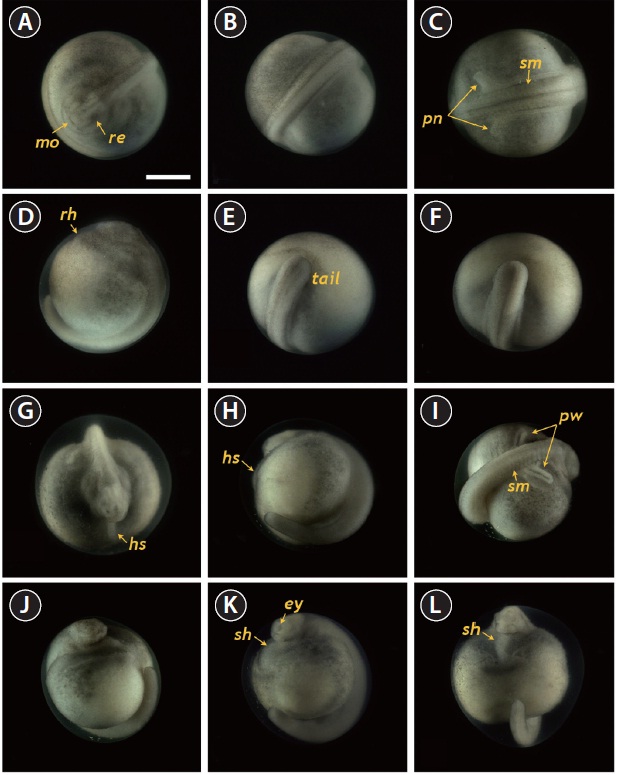
The developmental staging results are listed in Table 1. Upon fertilization, a black pigment developed at the animal pore, and the first cleavage furrow appeared at the animal pore at 2 h post-fertilization (HPF) (Fig. 1A). This cleavage furrow was limited to the animal hemisphere (Fig. 1B and 1C). Following the first cleavage (1 h later), the second cleavage formed typical 4-cell (four equal-sized blastomeres) embryos (Fig. 1D and 1E). Although the second cleavage furrow did not divide completely the vegetal hemisphere, partial infiltration of the vegetal hemisphere was observed (Fig. 1F). At 4 HPF, 8-cell embryos (similarly sized blastomeres) were formed in the animal hemisphere through the third cleavage event (Fig. 1G and 1H). At that time-point, the cleavage infiltrated the entire vegetal hemisphere and the vegetal hemisphere
[Table 1.] Staging of embryonic development in Siberian sturgeon Acipenser baerii
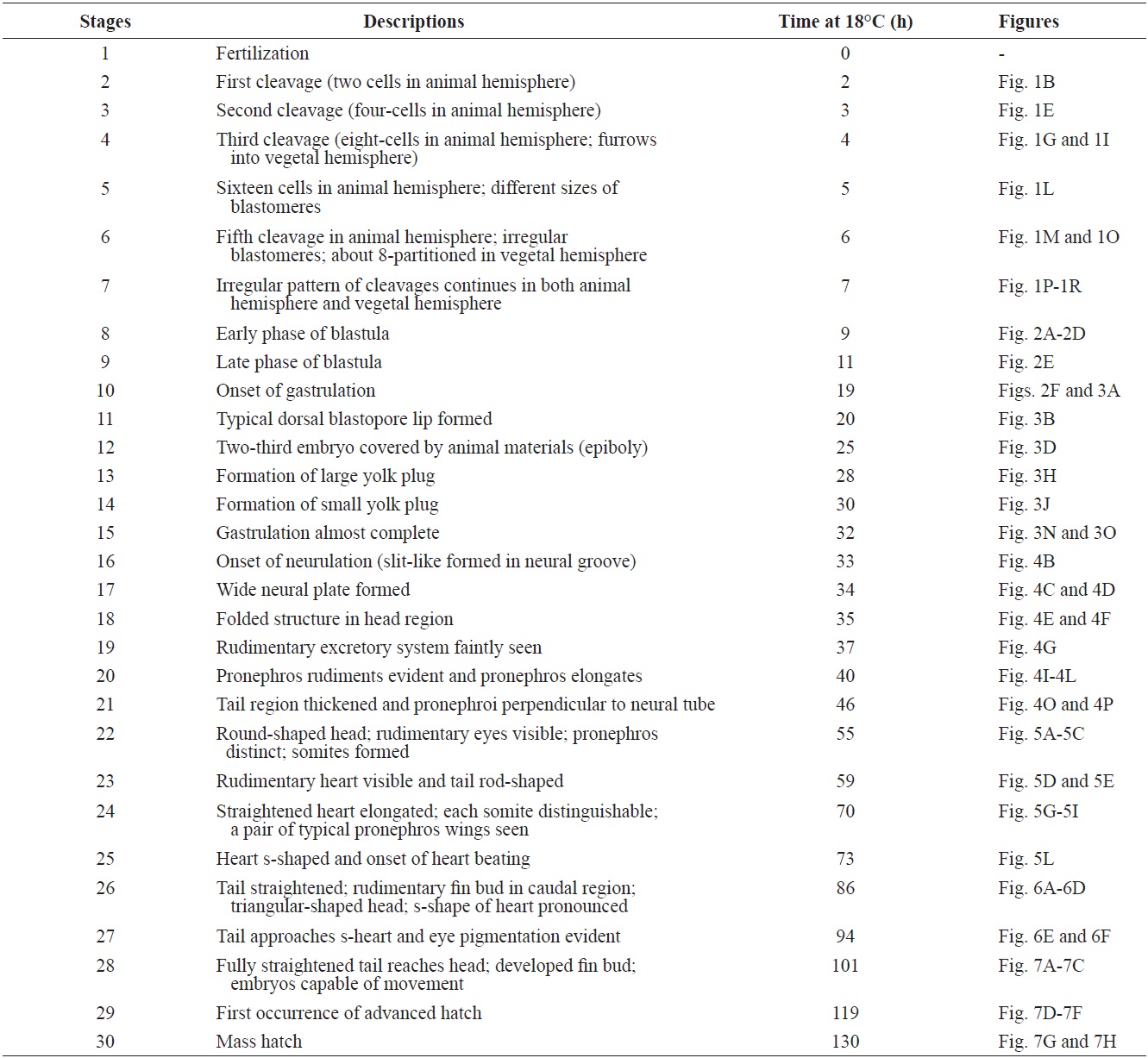
Staging of embryonic development in Siberian sturgeon Acipenser baerii
was completely divided into four similarly sized compartments (Fig. 1I). When the 16 blastomeres were formed in the animal hemisphere (at 5 HPF; fourth cleavage), one half (newly formed) of the 16 blastomeres in the center of the animal hemisphere were much smaller than the remaining half (earlier ones) (Fig. 1J-1L). The sizes and shapes of the blastomeres that resulted from subsequent cleavages in the animal hemisphere were noticeably dissimilar (Fig. 1M and 1N), and the furrows in the vegetal hemisphere were also formed in an irregular manner (Fig. 1O). This irregular pattern of division continued in both the animal (Fig. 1P and 1Q) and vegetal (Fig. 1R) hemispheres.
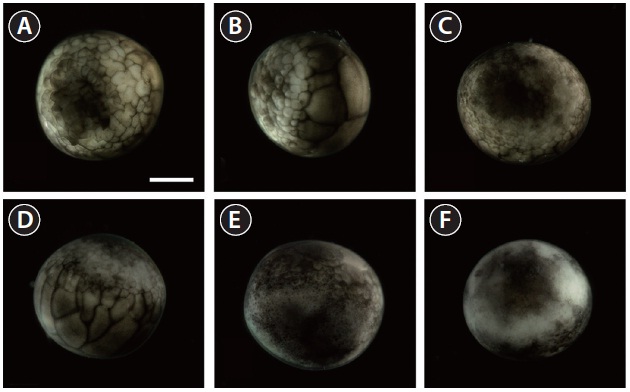
As cleavage continued, small blastomeres proliferated in the animal hemisphere (Fig. 2A), concomitant with irregular divisions in the vegetal hemisphere (Fig. 2B). The blastomeres in the animal hemisphere became uncountable (Fig. 2C), although large blastomeres were still formed in the vegetal
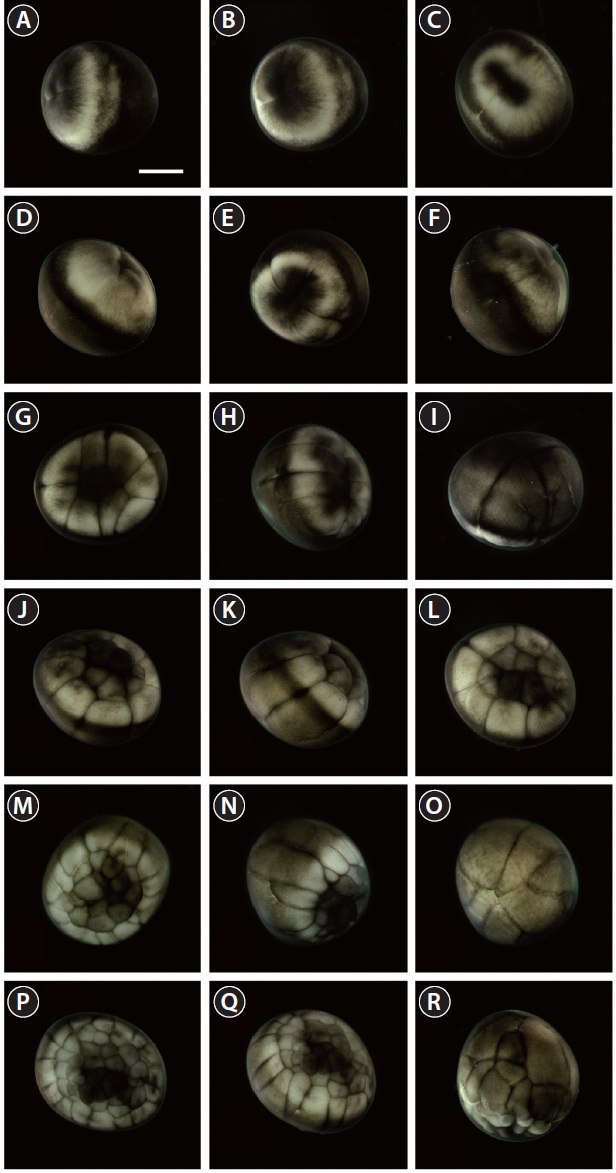
hemisphere (Fig. 2D). Cell division meant that the blastomeres became indistinguishable from one another in the animal hemisphere under the microscope at low-power magnification, and a cleavage cavity (blastocoel) began to form at the apex of the animal hemisphere (9 HPF). Meanwhile, in the vegetal hemisphere, relatively small blastomeres were located near the marginal zone (the equator), while larger blastomeres were evident in the vegetal apex region. At 11 HPF, the primordial cleavage cavity became larger and more evident in the animal hemisphere, whereas cell division continued to generate smaller blastomeres in the vegetal hemisphere (Fig. 2E). Nearing the completion of blastulation (18-19 HPF), the smooth appearance of the animal hemisphere changed to a milky white blastula roof (Fig. 2F).
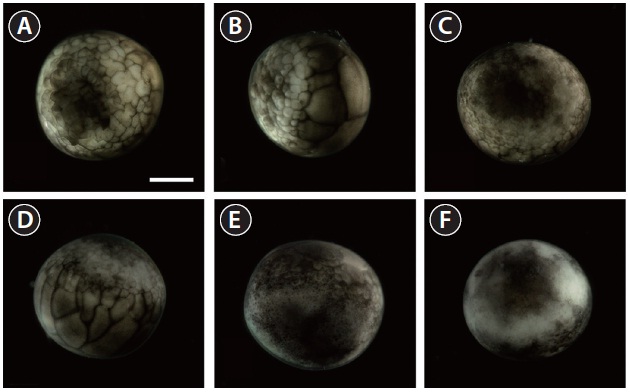
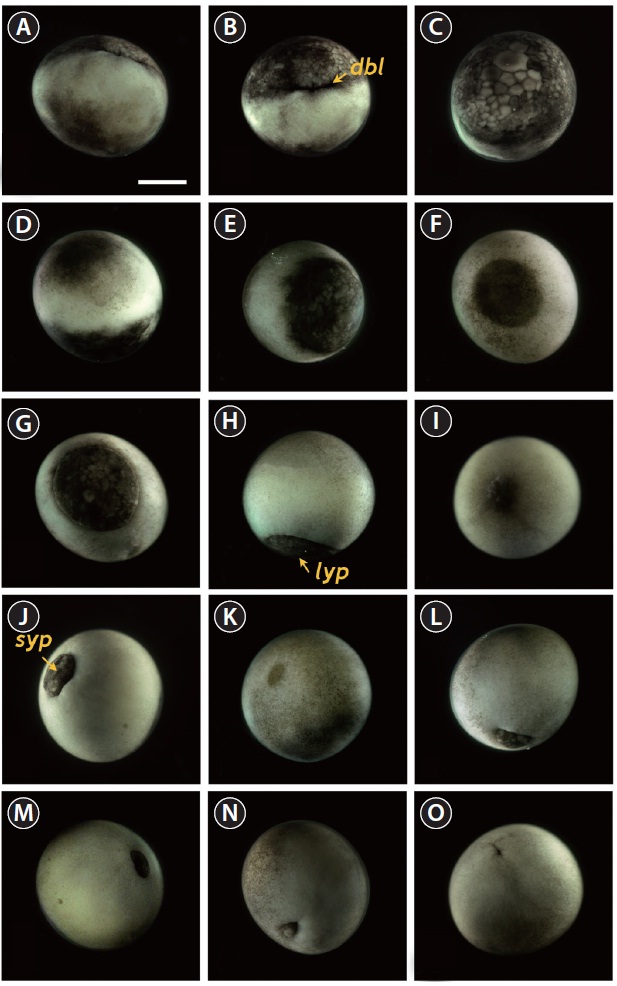
Upon the onset of gastrulation (19-20 HPF), a band was formed close to the equator, and this was followed by the formation of a “dorsal blastopore lip” between the animal and vegetal hemispheres (Fig. 3A and 3B). At this time-point, the vegetal hemisphere contained a number of divided blastomeres of differing sizes, whereby relatively large, countable (distinguishable) blastomeres were present in the region close to the apex and smaller (often uncountable) blastomeres were detected in the region close to the marginal zone (Fig. 3C). The blastula roof of the animal hemisphere enveloped progressively the vegetal hemisphere. At 25 HPF, approximately two-thirds of the embryo were covered by blastoderm (Fig. 3D and 3E), and a darkly pigmented region was formed in a round shape at the surface of the animal pore in many, albeit not all, of the embryos (Fig. 3F). As gastrulation continued, epiboly covered more than two-thirds of the embryos, and the area of remaining vegetal material declined progressively (Fig. 3G). At 28 HPF, a large yolk plug was formed, and the darkly pigmented region at the animal pore decreased (Fig. 3H
and 3I). About 2 h later, the size of the yolk plug was further reduced to less than one-fifth the diameter of the embryo (i.e., small yolk plug formation) (Fig. 3J). The pigmented region at the surface of the animal pore was also markedly reduced in size and coloration (Fig. 3K). The size of the yolk plug gradually decreased (Fig. 3L) until the blastopore appeared only as a small circle at the apex of the vegetal pore (Fig. 3M and 3N). As gastrulation neared completion (32 HPF), the blastopore took on a slit-like appearance, which signaled the initiation of neurulation (Fig. 3O).
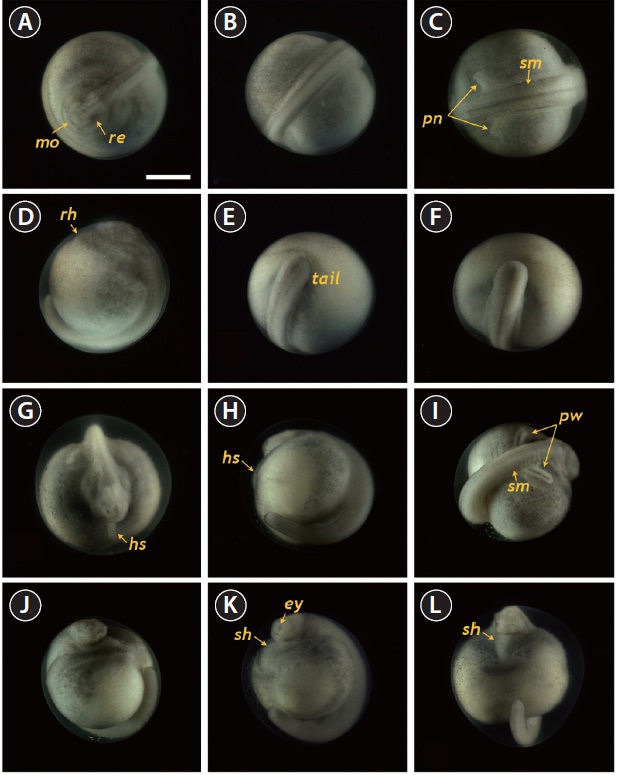
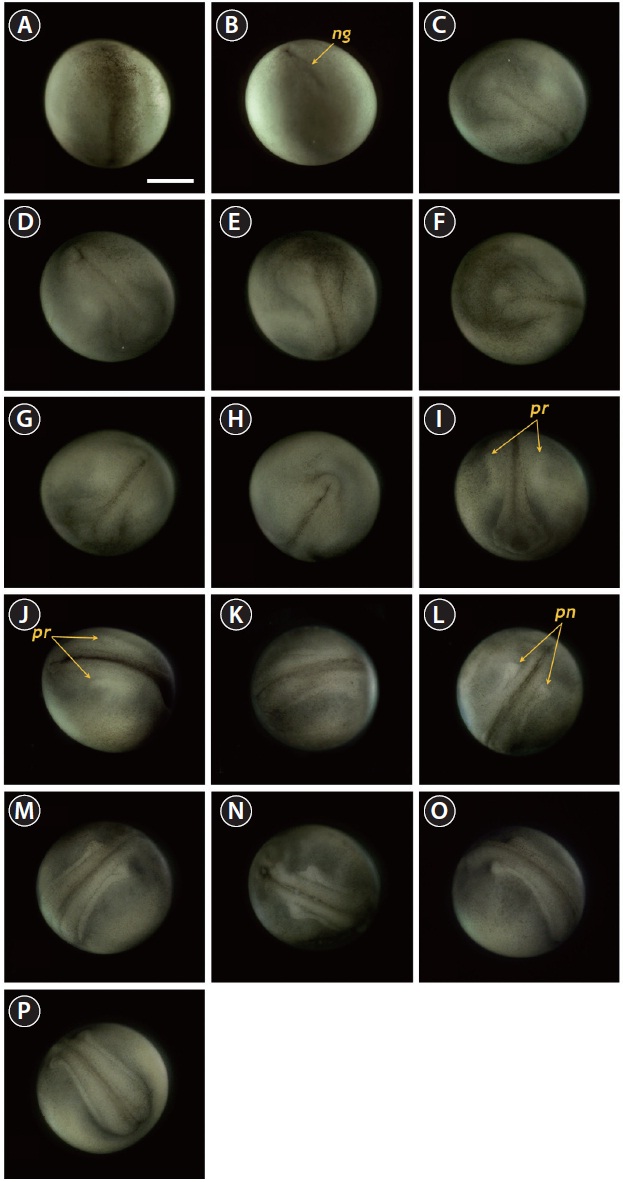
Neurulation began with the appearance of a slit-shaped
neural groove in the blastopore (Fig. 4A and 4B), followed by the formation of the neural plate at the dorsal surface with the folded structure in the head region (33 HPF) (Fig. 4C and 4D). With the progression of development (33-35 HPF), the neural plate in the head region widened and the neural folds rose and thickened (Fig. 4E and 4F). In the dorsal region, the neural groove was more evident and the excretory system rudiments were faintly visible parallel to the neural groove (37 HPF) (Fig. 4G). At that time-point, a folded shape was seen in the tail region (Fig. 4H). As neurulation continued, the folding
of the head region was slightly reorganized and a pair of pronephros rudiments became more evident as cords running parallel to the neural groove (40 HPF) (Fig. 4I and 4J). The neural folds rose significantly and the anterior part of the pronephros was visualized as a distinct form (Fig. 4K and 4L). Thereafter, the pronephros became elongated and thickened along with the risen neural folds (Fig. 4M and 4N). The tail region continued to thicken significantly as development progressed, although it had not yet detached from the yolk sac membrane. At this stage, the pronephroi were located almost perpendicular to the neural tube and the neural tube was mostly closed (46 HPF) (Fig. 4O and 4P).
>
Progress to the onset of heart beating
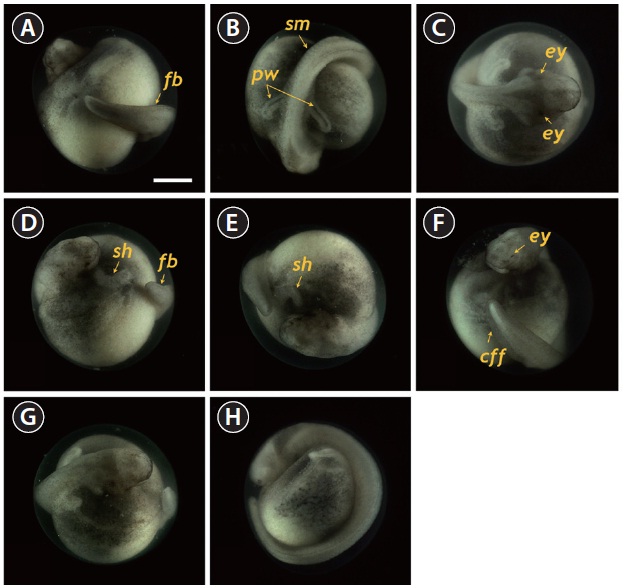
At 55 HPF, the lateral plates were fused to the prosencephalon (forebrain), and the head region appeared as a roundshaped object in the dorsal view, in which the rudimentary
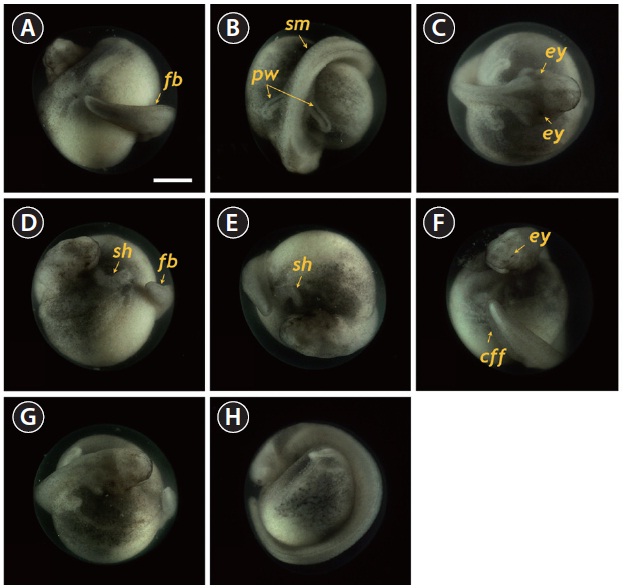
eyes and undeveloped mouth were easily visible (Fig. 5A). At this stage, the dorsal region had a rod-shaped appearance with developed somites, and the primary pronephros-wings were visible (Fig. 5B and 5C). A rudimentary heart formed in the embryo (59 HPF), although the head had not yet separated (Fig. 5D). In the caudal region, the flattened tail was transformed to a rod-shaped structure and the tail had begun to separate from the yolk sac (Fig. 5E and 5F). As development continued (60-70 HPF), the head region thickened and began to separate from the yolk, in which the heart was seen as a short straight tube (Fig. 5G). The tail continued to lengthen and pronounced somites were visible over the entire embryonic body (Fig. 5H and 5I). At this point, a pair of highly differentiated pronephros-wings could be observed (Fig. 5I). At 73 HPF, the heart became s-shaped (the so-called s-heart) and began to beat. The eyes were more distinct in this stage (Fig. 5J-5L).
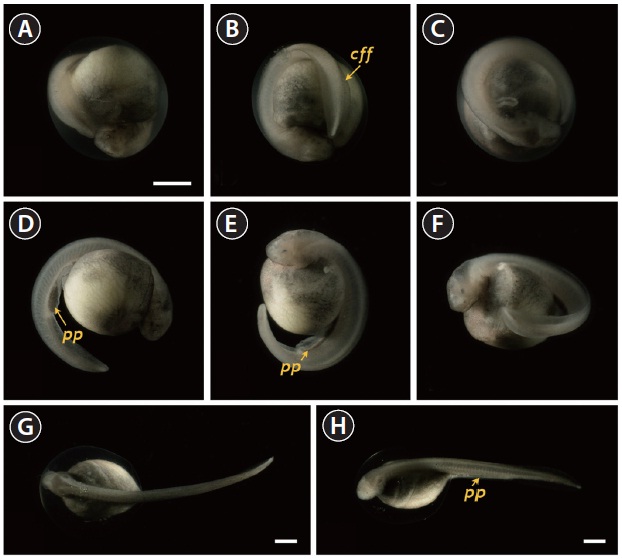
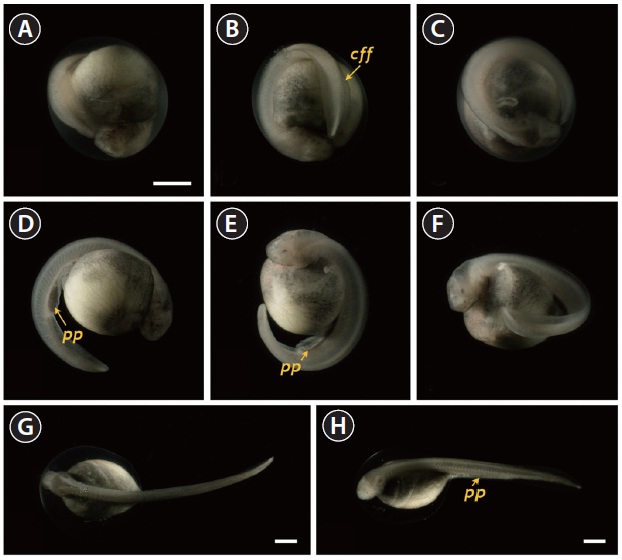
At 86 HPF, the round, rod-shaped structure of the tail straightened, and fin-bud rudiments were visible in the caudal region (Fig. 6A). Other morphologic features typical of the embryos at this stage were: separation of the anterior part of the head from the yolk sac; the triangular shape of the developed head; slightly pigmented eyes; and a more pronounced s-heart (Fig. 6B-6D). At 94 HPF, the tail reached the beating
s-heart and fin folds appeared in the caudal region (Fig. 6E- 6H). Blood circulation was easily observable, and olfactory organs were also seen. As the tail continued to lengthen, it approached the head (101 HPF). Eye caps were evident and caudal fin folds were easily distinguishable (Fig. 7A-7C). The embryos at this stage were capable of movement. From 119 HPF, several advanced embryos began to hatch, and mass hatching was observed at 130 HPF. The average hatching success based on duplicate examinations was 80.5%. Individuals from advanced hatches were often bent in shape (Fig. 7D-7F), whereas most of the mass-hatched prolarvae (
Unlike many teleostean embryos that show the meroblastic cleavages on the blastodisc exclusively in the animal hemisphere, sturgeon embryos display holoblastic cleavage (Conte et al., 1988). In general, the holoblastic cleavage pattern observed in the present study is in agreement with those previously defined for other Acipenseriform fishes (Dettlaff et al., 1993). In similarity with other sturgeon species, the Siberian sturgeon exhibits an uneven pattern of holoblastic cleavage in which the vegetal hemisphere is not completely divided with each cleavage furrow. This asymmetrical cleavage pattern in sturgeon embryos is different from the holoblastic cleavage seen in anuran embryos. This difference is attributable to the uneven distribution of yolk constituents between the animal and vegetal hemispheres in sturgeon embryos, a consequence of which is a reduction of the cleavage rate in the vegetal hemisphere, which contains more yolk content than the animal hemisphere (Colombo et al., 2007).
These differential rates of cleavages result in differently sized and irregular-shaped blastomeres in the animal and vegetal hemispheres at the beginning of blastulation. With the progression of blastulation, a marginal zone of transition between the animal and vegetal hemispheres is formed with intermediate-sized blastomeres. In this zone, a “dorsal lip” appears as a marker of the initiation of gastrulation in sturgeon embryos (Bolker, 1993). Following the primary internalization by involution, germ layers arise and the overall plan for the body architecture is programmed during gastrulation. This usually involves a complex set of morphogenetic movements of cellular progenitors, syncytial yolk cells, and an extra-embryonic sheath (enveloping layers) (Cooper and Virta, 2007; Shook and Keller, 2008). During gastrulation, the cell layers (
In the present study, a complete set of images of normal embryonic development was prepared, together with embryologic descriptions for 30 developmental stages of the Siberian sturgeon. Based on the results of this study, the effects of integral water temperature on the mitotic divisions and development of sturgeon might be examined in future studies. The results of the present study could be useful in hatchery management for the reproductive control of the Siberian sturgeon, and constitute a basis for future studies of developmental gene expression in this sturgeon species.