Ulmi Pumilae Cortex (UPC) is a deciduous tree with uneven pinnate leaves and is classified as a subfamily of Ulmuceae and contains many pharmacologically active constituents. The aim of this study was to investigate the effects of UPC on the growth and survival of AGS cells, the most common human gastric adenocarcinoma cell lines.
The AGS cells were treated with varying concentrations of UPC. Analyses of the sub G1, caspase-3 activity, and mitochondrial depolarization were conducted to determine whether AGS cell death occured by apoptosis. Furthermore, to identify the role of the transient receptor potential melastatin (TRPM) 7 channels in AGS cell growth and survival, we used human embryonic kidney (HEK) 293 cells overexpressed with TRPM7 channels.
The addition of UPC to a culture medium inhibited AGS cell growth and survival. Experimental results showed that the sub G1, caspase-3 activity, and mitochondrial depolarization were increased. Furthermore, TRPM7 channel overexpression in HEK 293 cells exacerbated UPC-induced cell death.
These findings indicate that UPC inhibits the growth and survival of gastric cancer cells due to a blockade of the TRPM7 channel activity. Therefore, UPC is a potential drug for treatment of gastric cancer, and TRPM7 channels may play an important role in survival in cases of gastric cancer.
Interest in alternative therapies and the therapeutic use of natural products, especially those derived from plants has been growing. This interest in drugs of plant origin has several reasons; namely, conventional medicine can be inefficient (side effects and ineffective therapy), and abusive, and incorrect use of synthetic drugs can result in side effects and other problems [1].
Gastric and breast cancer are leading causes of cancer-related mortality in Korea. In previous studies, we suggested that human gastric adenocarcinoma cells expressed the transient receptor potential melastatin 7 (TRPM7) channel, which is essential for cell survival and is a potential target for pharmacological gastric cancer treatment [5]. TRPM7 is a member of the large TRP channel superfamily that is expressed in nearly every tissue and cell type [6-8]. Many reports suggest that activated TRPM7 channels contribute to a number of physiological and pathophysiological processes [9-11]. However, the role of the TRPM7 channel in the survival of gastric cancer cells after incubation with UPC is unknown. In this study, we examined the effects of UPC and the role of TRPM7 channels in UPC-inhibited apoptosis of AGS cells, the most common human gastric adenocarcinoma cell lines.
The UPC used in this study was purchased from the Korea Research Institute of Bioscience and Biotechnology (KRIBB).
The AGS cell lines were that were used established at the Cancer Research Center, Seoul National University College of Medicine, Korea. The cell lines were propagated in RPMI-1640 medium (Gibco-BRL) supplemented with 10% heat-inactivated fetal bovine serum and 20-μg/mL penicillin and streptomycin in an atmosphere of 5% CO2 at 37℃ .
In order to investigate whether the cell cycle of AGS cells was redistributed, a flow cytometric analysis was used with propidium iodine (PI) stain [12, 13]. 1 x 106 cells were placed in an e-tube. 700 ㎕ of an ice-cold fixation buffer (ethyl alcohol) was slowly added with vortexing. Tubes were sealed with parafilm and incubated at 4℃ overnight. Samples were spun for 3 min at 106 g at 4℃ , and the supernatant was aspirated and discarded. The cell pellet was resuspended in 200 ㎕ of PI staining solution (PI [5 mg/mL] 2 ㎕ and RNase 2 ㎕ in PBS 196 ㎕) at 20817 g for 5 s. After 30 min in the dark at room temperature, samples were analyzed in a fluorescence activated cell sorter (FACScan; Becton-Dickinson, Moutain View, CA, USA) at λ= 488 nm by using Cell-Quest software (Becton-Dickinson). The DNA content distribution of normally growing cells was characterized by using two peaks, the G1/G0 and the G2/M phases. The G1/G0 phase comprises the normal functioning and resting state of the cell cycle with the most diploid DNA content while the DNA content in the G2/M phase is more than diploid. Cells in the sub-G1 phase have the least DNA content in the cell cycle distribution; this is termed hypodiploid. The hypoploid DNA content represents the DNA fragmentation [14].
2.4. MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5-
diphenyltetrazolium bromide) assay
Cell viability was assessed by using a MTT assay. The AGS cells were seeded into each well of 12-well culture plates and then cultured in Roswell Park Memorial Institute medium (RPMI)-1640 supplemented with other reagents for 72 h. After incubation, 100 ㎕ of MTT solution (5 mg/mL in phosphate-buffered saline (PBS)) was added to each well, and the plates were incubated at 37 ℃ for 4 h. After the supernatant had been removed and shaken with 200 ㎕ of dimethyl sulfoxide (Jersey Lab Supply, Livingston, NJ, USA) for 30 min, the absorbance was measured at 570 nm. All experiments were repeated at least 3 times.
Caspase-3 assay kits (Cellular Activity Assay Kit Plus) were purchased from BioMol (Plymouth, PA, USA). After experimental treatment, cells were centrifuged (10000 g, 4 ℃ , 10 min) and washed with PBS. Cells were re-suspended in an ice-cold cell lysis buffer and incubated on ice for 10 min. Sample were centrifuged at 10000 g (4 ℃ , 10 min), and the supernatant was removed. Supernatant samples (10 ㎕ ) were incubated with a 50 ㎕ of substrate (400-μM Ac- DEVD-pNA) in a 40-㎕ of buffer at 37 ℃ . The absorbance at 405 nm was read at several time points. The pNA concentrations in the samples were extrapolated from a standard created using the absorbances of sequential pNA concentrations.
2.6. Assessment of mitochondrial membrane depolarization
Mitochondrial membrane depolarization was evaluated using a JC-1 fluorescence probe according to the manufacturer's instructions (Santa Cruz). The AGS cells were labeled with 2-μM JC-1 for 30 min at 37 ℃ and then analyzed by using flow cytometry with 488-nm excitation and 530/30- or 585/42-nm bypass emission filters. The cells without red fluorescence were regarded as the cells manifesting mitochondrial membrane depolarization.
The whole-cell configuration of the patch-clamp technique experiment was performed at room temperature (22-25 ℃ ). The AGS cells were transferred into a small chamber on an inverted microscope stage (IX70, Olympus, Japan) and were constantly perfused with a solution containing 2.8-mmol/L KCl, 145- mmol/L NaCl, 2-mmol/L CaCl2, 10-mmol/L glucose, 1.2-mmol/L MgCl2, and 10-mmol/L 4-(2-hydroxyethyl)- 1-piperazineethanesulfonic acid (HEPES), adjusted to a pH of 7.4 with NaOH. The pipette solution contained 145-mmol/L Cs-glutamate, 8-mmol/L NaCl, 10- mmol/L Cs-2-bis(2-aminophenoxy)-ethane-N,N,N',N'- tetraacetic acid, and 10-mmol/L HEPES-CsOH adjusted to a pH of 7.2 with CsOH. Axopatch I-D (Axon Instruments, Foster City, CA, USA) was used to amplify the membrane currents and potentials. For data acquisition and the application of command pulses, pCLAMP software v.9.2 and a Digidata 1322A units (Axon Instruments) were used. Results were analyzed using pClamp and Origin software (Microcal Origin version 6.0).
2.8. TRPM7 expression in human embryonic kidney
293 cells
Human embryonic kidney (HEK)-293 cells were transfected with the Flag-murine long transient receptor potential channel 7 (LTRPC7)/pCDNA4-TO constructed and grown on glass coverslips in Dulbecco's modified Eagle medium supplemented with 10% fetal bovine serum, blasticidin (5 μg/mL), and zeocin (0.4 mg/mL). TRPM7 (LTRPC7) expression was induced by adding 1 μg/mL of tetracycline to the culture medium. Whole-cell patch-clamp experiments were performed at 21-25 ℃ with cells that had been grown on the glass coverslips.
Data are expressed as means ± standard errors of the mean (SEM). Differences between the data were evaluated by using the student's
3.1. UPC-induced cell death in AGS cells
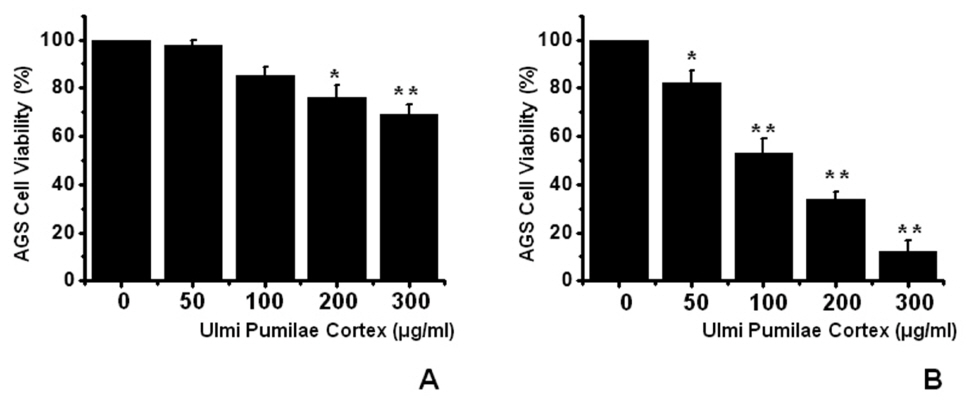
To ascertain whether UPC kills AGS cells, we performed MTT assays. The viable cell population was gradually reduced with increasing concentrations of UPC for 24 h (Fig 1)A), and with the IC50 value of 107 μg/ml for 72 h for AGS cells (Fig 1)B). Thus, our results demonstrate that UPC induces cell death in AGS cells.
3.2. UPC-triggered apoptosis in AGS cells
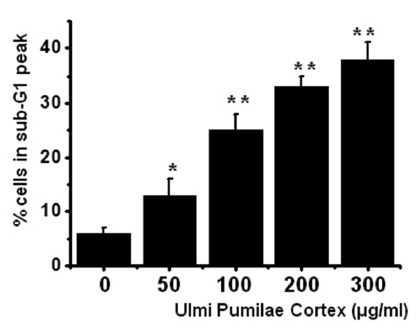
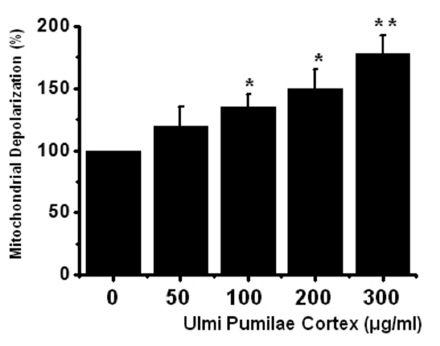
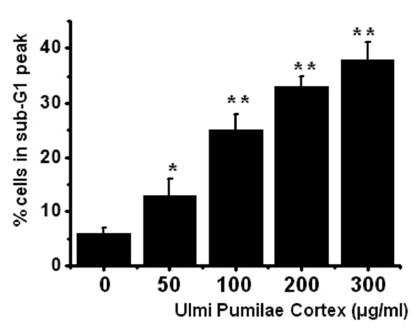
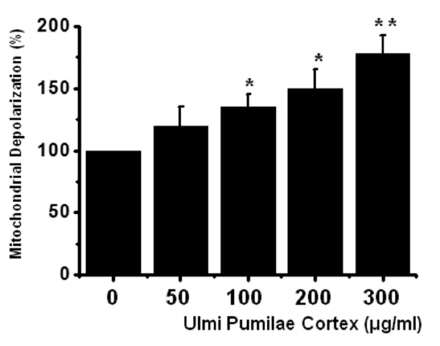
To determine whether AGS cell death occured by apoptosis, we conducted a sub-G1 analysis [15, 16]. In this protocol, cells were incubated with UPC and stained with a fluorescent DNA stain (PI). The action of endogenous endonucleases in apoptotic cells cleaves DNA into endonucleosomal fragments of typical size, which are extracted from the cells. The loss of DNA is detected by using a FACS analysis, as reduced nuclear staining in the apoptotic cells, which results in a novel (sub-G1) fluorescence peak to the left of the regular fluorescence peak. Flow cytometric analysis showed that the percentage of sub-G1 phase cells was markedly increased in the cells treated with UPC in a dosedependent manner in AGS cells (Fig 2) In addition, UPC elevated mitochondrial membrane depolarization, an early event of an intrinsic apoptosis signaling (Fig 3)
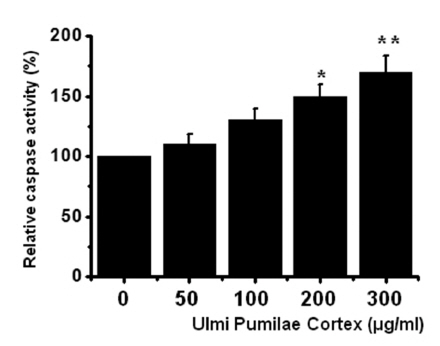
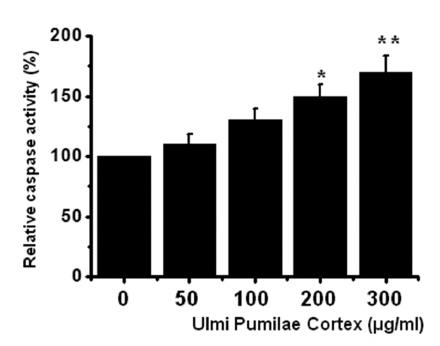
Thus, our findings suggest that UPC induces apoptosis via intrinsic apoptotic mechanism(s). Caspase-3 activation is one of the hallmarks of apoptotic cell death. We also measured the enzyme activity in AGS cells after UPC incubation. Using a synthetic substrate, we detected the caspase-3 activity in AGS cells. UPC increased the activity of caspase-3 (Fig 4)
3.3. Effects of UPC in TRPM7 currents in AGS and TRPM7
overexpressed HEK293 cells
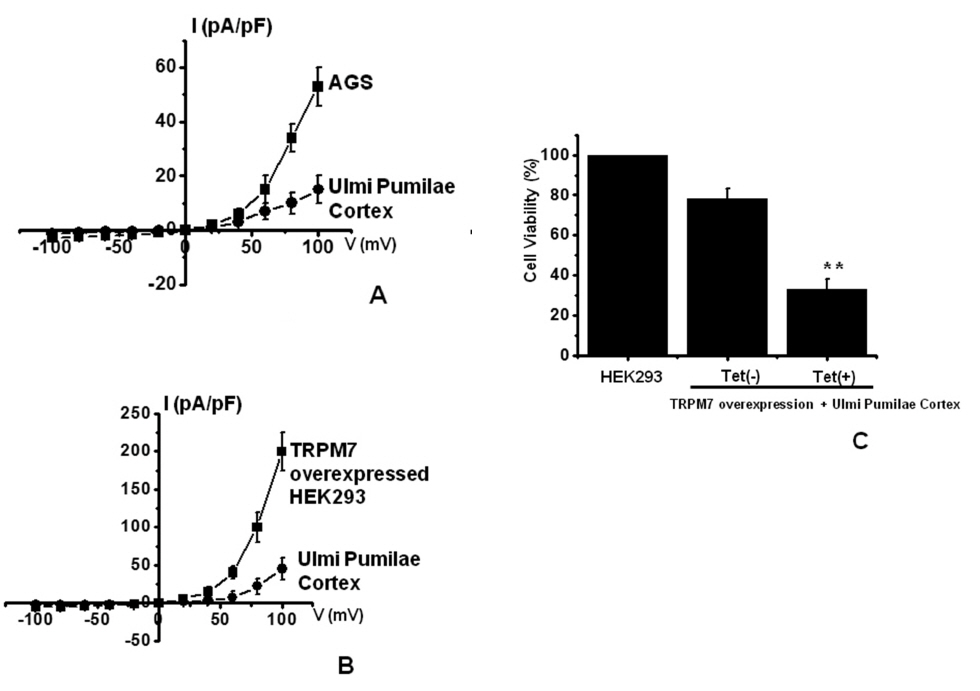
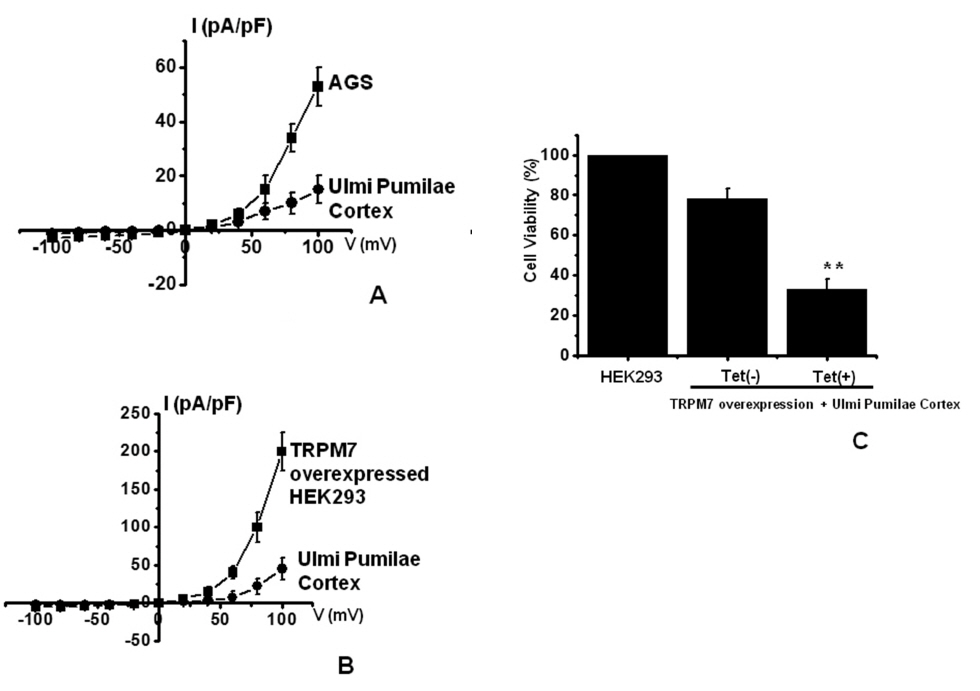
TRPM7 has been proposed to be required for cell survival on the basis of experiments on geneticallyengineered DT-40 B-cells [17]. Also, we recently suggested, as in previous reports, that AGS cells express the TRPM7 channel and that suppression of the TRPM7 channel induced cell death [5]. Therefore, we investigated whether UPC influences TRPM7 currents in AGS cells. To confirm the effect of UPC in TRPM7 currents, we investigated the effects of UPC in AGS cells by using patch-clamp techniques. We performed whole cell voltage-clamp recordings to investigate the effect of UPC on TRPM7-like currents in AGS cells. A voltage ramp from +100 mV to -100 mV evoked small inward currents at negative potentials whereas larger outward currents were evoked at positive potentials, showing that they were outward-rectifying cation currents (n = 4; (Fig 5A). However, in the presence of 300 μg/ml UPC, the amplitudes of these currents were inhibited outwardly by 70.4 ± 2.1% and inwardly by 85.3 ± 2.3% (n = 4; (Fig 5A). Also, similar results were obtained in HEK293 cells overexpressing TRPM7 (Fig.5B). To provide additional evidence that supports the contribution of the TRPM7 channel to UPC toxicity, we investigated the changing expression levels of the TRPM7 channel and its influences on UPC-mediated cell death. We used HEK293 cells with inducible TRPM7 channel expression [10, 11]. In the absence of induced TRPM7 channel expression [TRPM7(-) cells, Tet(-)], incubation of HEK293 cells with UPC induced cell death in the MTT assay (n = 5; (Fig 5C). However, when TRPM7 channel overexpression was induced by adding tetracycline [TRPM7 (+) cells, Tet (+)], incubation of HEK293 cells with UPC induced cell death at an increased rate in the MTT assay, which suggests that increased expression of TRPM7 channels leads to an increased rate of UPC-induced cell death.
Apoptosis is a selective process of physiological cell deletion that plays an important role in the balance between cell replication and cell death [1]. Because cancer chemotherapeutic as well as, chemopreventive, agents have recently been suggested to exert their pharmacological effects by triggering
Apoptosis is a selective process of physiological cell deletion that plays an important role in the balance between cell replication and cell death [1]. Because cancer chemotherapeutic as well as, chemopreventive, agents have recently been suggested to exert their pharmacological effects by triggering apoptotic cell death or cell cycle transition, the induction of apoptosis in tumor cells has become a predictor of tumor treatment response [1, 18-21]. Therefore, research on apoptosis is very important and significant.
The potential use of medicinal plants as a source of new drugs is still poorly explored. Of the estimated 500,000 plants species, only a small percentage has been investigated phytochemically and even a smaller percentage has been properly studied in terms of their pharmacological properties; in most cases, only pharmacological screening or preliminary studies have been carried out [1]. Plasma-membrane ion channels participate in cellular electrogenesis, electrical excitability, and virtually all basic cellular behaviors, including such crucial ones for maintaining tissue homeostasis as proliferation, differentiation, and apoptosis [22-25]. The involvement of ion channels in the regulation of cellular proliferation or apoptosis in vitro has been known since, at least, the late 1980s based on observations that classical blockers of these channels can influence the rate of cell death by prolonging or shortening cell survival. Ion channels are crucial to tumor development and cancer growth. The TRPM7 cation channel supports multiple cellular and physiological functions, including cellular Mg2+ homeostasis [26], cell viability and growth [17, 27], anoxic neuronal cell death [28], synaptic transmission [29], cell adhesion [30], and intestinal pacemaking [31]. Wykes et al. [32] suggested that TRPM7 channels were critical to human mast cell survival. Jiang et al. [10] suggested that activation of TRPM7 channels was involved in the growth and proliferation of human head and neck carcinoma cells. Abed et al. [33] proposed the importance of TRPM7 in the human osteoblast-like cell proliferation. Also, Guilbert et al. [34] suggested that TRPM7 was required for breast cancer cell proliferation. In a previous study, we suggested that TRPM7 channels played an important role in the growth and the survival of gastric cancer cells [5].
Therefore, TRPM6 might also have an important role in apoptosis, so we should investigate the involvement of TRPM6 in the future. Among the TRP families, TRPC, TRPV and TRPM are mainly related to the growth and progression in cancer cells. Depending on the type of cancer, regulation of TRP mRNA and protein expression has been changed. These ion channel changes are related with cell growth and apoptotic-induced cell death in cancer cells. Therefore, the regulations of ion channels in cancer cells are the most promising strategy, and considerable efforts should be done to fight cancer cells [37]. In line with these studies, our studies show that UPC induces apoptosis in human gastric adenocarcinoma cells, which may be due to a blocking of the TRPM7 channel activity.
UPC has been used as a constituent of oriental herb medicine prescriptions to produce a variety of physiologic or pharmacologic effects in various regions. However, only a few reports have described the effects of UPC on gastric cancer. In summary, UPC inhibited the growth and the survival of AGS cells. Sub G1 and caspase-3 activities were elevated, and the mitochondrial membrane depolarization potentials were increased. TRPM7 currents were inhibited by UPC. Furthermore, overexpression of TRPM7 channels in HEK 293 cells increases the rate of UPC-induced cell death. These findings indicate that UPC inhibits the growth and survival of gastric cancer and that the UPC-induced apoptosis is due to a blockade of TRPM7 channel activity. Therefore, UPC is a potential drug for the treatment of gastric cancer, and TRPM7 channels may play an important role in survival in cases of gastric cancer.

![Ulmi Pumilae Cortex (UPC) induces cell death in AGS cells. MTT (3-[4, 5-dimethylthiazol-2-yl]-2, 5-diphenyltetrazolium
bromide)based viability assay. The AGS cells were treated with increasing concentrations of UPC for 24 h (A) and for 72 h (B). Distilled water
was used as a vehicle. Cell viability is expressed as a value relative to that of the untreated cells which is set to 100%. The figures show mean
± SEM. *P < 0.05, **P < 0.01.](http://oak.go.kr/repository/journal/11870/DHOCBS_2013_v16n2_55_g001.jpg)