Approximately 60% of Iran is classified as arid and semi-arid land. Much of the country has a desert climate with an average annual precipitation of less than 300 mm (Mansoory 1992). Sadeghian et al. (2010) reported that milkweed (Calotropis procera L.) can adapt to arid land. This species is one of the most important deciduous tree desert species in south Iran. It is very apparent that the effects of human activities and livestock on dry land have severely limited tree growth in some regions. Cutting and grazing has drastically reduced the extent of many forests throughout the country, and sufficient consideration of plant’s success rate, especially seedling vigor, must be made prior to re-planting efforts (El-Moslimany 1986). Calotropis sp. produces lots of small seeds which are dispersed by wind, but has a low density per unit area because its germination stage is sensitive to drought (Bazrafkan 2011). Seed germination performs an important function in the regeneration of plant species, particularly under arid and unpredictable environmental conditions, most notably those of Mediterranean ecosystems (Gimenez- Benavides et al. 2005). Low seedling vigor, especially poor emergence and seedling establishment are regarded as major causes of these low densities (Close and Wilson 2002). Seed emergence, seedling establishment and survival are affected by light, climate and edaphic and species-specific factors (Bewley and Black 1994). Thus, any study of dry land restoration should ideally separate the effects of individual environmental factors and their interactions on demographic stages. In dry lands, emergence, seedling growth and survival are affected by light intensity (Khurana and Singh 2001). The reflex of emergence to light intensity depends on the individual species as well as the environment. Biomass production, relative growth rate, root/shoot ratio, specific leaf area, net assimilation rate, and leaf area ratio can be limited by either poor light availability (Rinco?n and Huante 1993) or low soil moisture, due to the removal of water from the dry forest understory by nurse trees (Rodriguez-Calcerrada et al. 2008). Partial shade protects soil moisture during seed emergence in dry forests. In dry lands, species emergence and early establishment must occur during the beginning of wet season when water is most plentiful (Khurana and Singh 2001). In dry lands, most precipitation occurs in winter and spring (Ginley 2008). When temperature rises quickly, water evaporates and rainfall may be extremely scarce during summer. Emergence can also be affected by water content of the growth medium. Soil water storage capacity depends on soil tissue and type. These characteristics positively affect the establishment of different plant species (Grange and Loach 1983, Newton et al. 1992). Perlite materials absorb water, and prevent the penetration of too much water to the lower layers of soil. Damizadeh (2004) used perlite as a growth medium for Capparis decidua. Yusef Saleh (2011) reported that sand mixed with perlite is the best growth medium for vegetative growth of Dahlia (Dahlia pinnata), Marigold (Tagetes erecta), Zinnia (Zinnia elegans) and Cosmos (Cosmos bipinnatus) plants. Abo-Rezq et al. (2009) used sand in growth media on selected plant species To facilitate the emergence. The general hypothesis of this study is that the combination of environmental factors and partial shade affects the early stages of the life cycle of a milkweed seed. This study aims to study the effects of growth medium and partial shade on early vigor of (Calotropis procera L.) under drought stress in mid-spring.
Mature seeds of milkweed (Calotropis procera L.) were collected from natural populations in the desert area in Sistan Balochestans province (60°N and 29°E), located in the elevations of 1,396 m above mean sea level (AMSL) in Southeast desert of Iran, and were stored in bags in a refrigerator (5℃) until the start of the experiment.
The split-split plot experimental design with three replications was carried out in a nursery. The main treatment plot was divided into two shade treatments (no shading and partial shading) and sub treatment plot1 included growth medium at four levels (G1 = clay [the soil type in the milkweed growth region], G2 = clay + sand, G3 = clay + perlite, G4 = clay + perlite + sand), and sub treatment plot 2 included drought (irrigation interval) at six levels (D1 = 2 [control], D2 = 4, D3 = 6, D4 = 8, D5 = per 10 and D6 = 12 days per for three month). Ten seeds were planted in each treatment area in a 1 liter plastic pot filled with the various soil type. The pots were watered according to the irrigation treatments. Vigor-related traits were the measured for the individual plants after two months. The traits were: emergence percentage, shoot length, root length, shoot dry weight, root dry weight, seedling dry weight and vigor index. Shoot and root dry weights were measured after drying for 24 h in an oven at 70℃ 2002). Vigor index was calculated for each treatment as follows (Abdul-Baki and Anderson 1973):
Vigor index = seedling length × emergence percentage
Data were checked for normality and were then analyzed using SAS ver. 9.1.3 (SAS Institue Inc., Cary, NC, USA). Treatment means were separated by Duncan’s multiple range test if the F-value of the treatments was significant at the 0.05 or 0.01 probability levels.
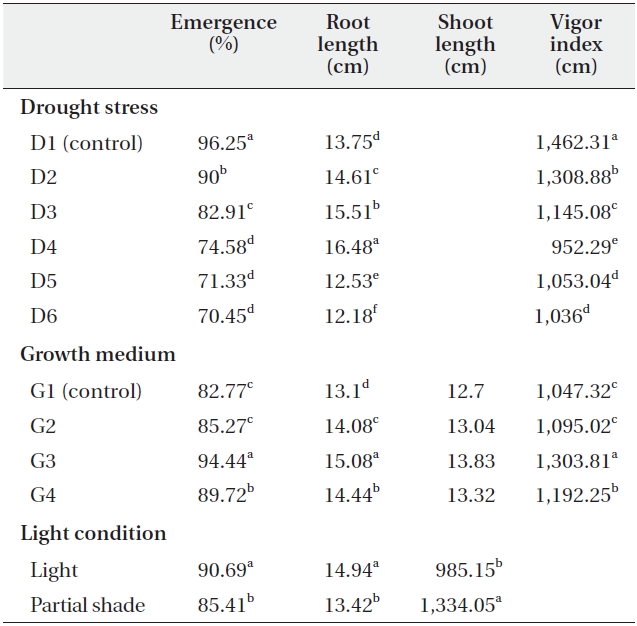
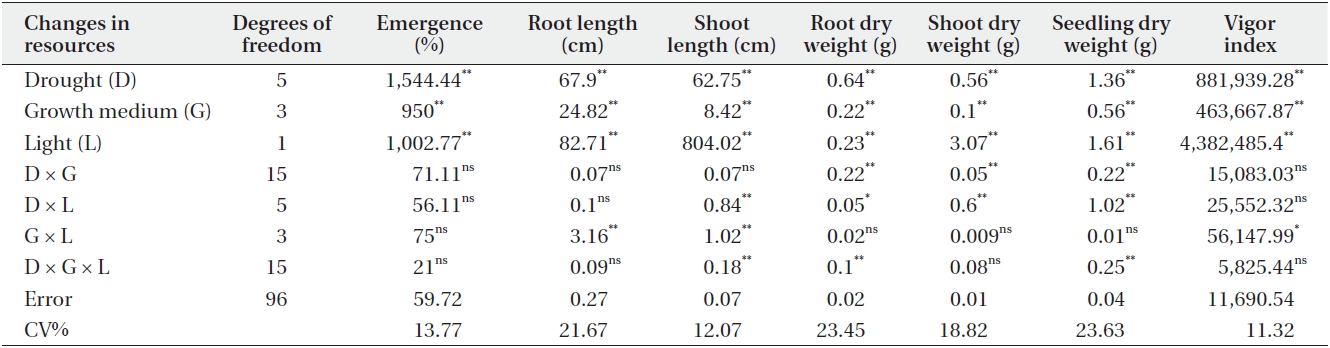
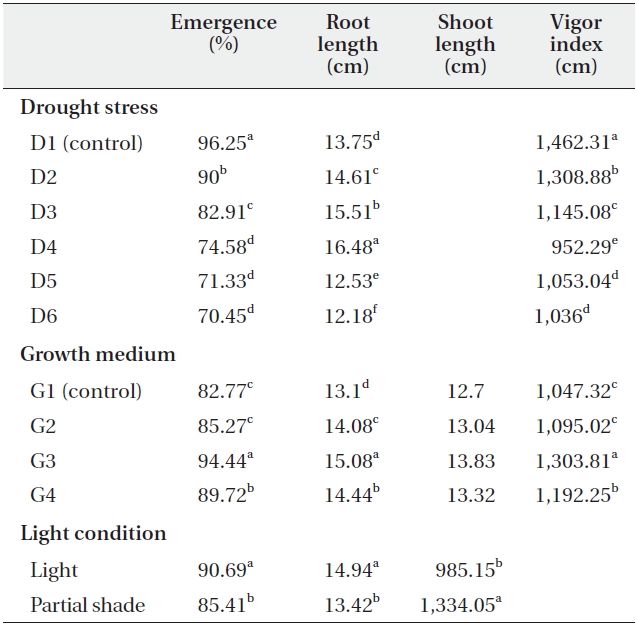
Drought stress and soil treatments had significant (P < 0.01) effects on emergence percentage (Table 1). The highest and lowest emergence percentage was obtained in D1 (control) and D6 irrigation interval respectively. The emergence percentage of C. procera L. decreased with increasing the irrigation intervals, with 96.25% emergence
in D1 (control) and 70.45% emergence in D6 irrigation interval treatment (Table 2). The growth medium had significant (P < 0.01) effects on emergence percentages, as 82.77% emerged in G1 (control) and 94.44% emerged in G3 (clay + perlite) (Table 2). Emergence percentage was significantly (P < 0.01) affected by light treatments also (Table 1). Partial shade treatment decreased the emergence percentage in all growth medium and irrigation treatments. The mean emergence percentages were 90.69% and 85.41% in light and partial shade, respectively (Table 2).
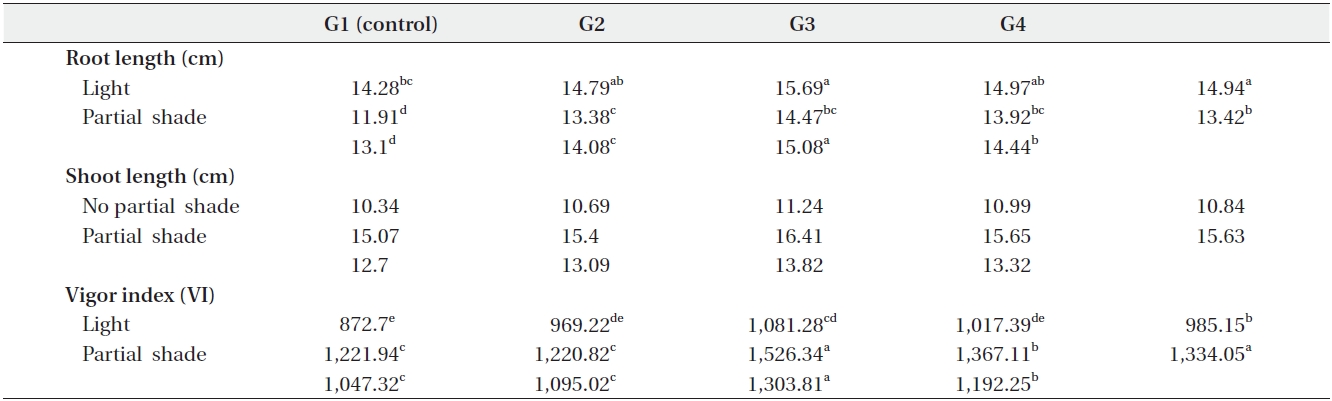
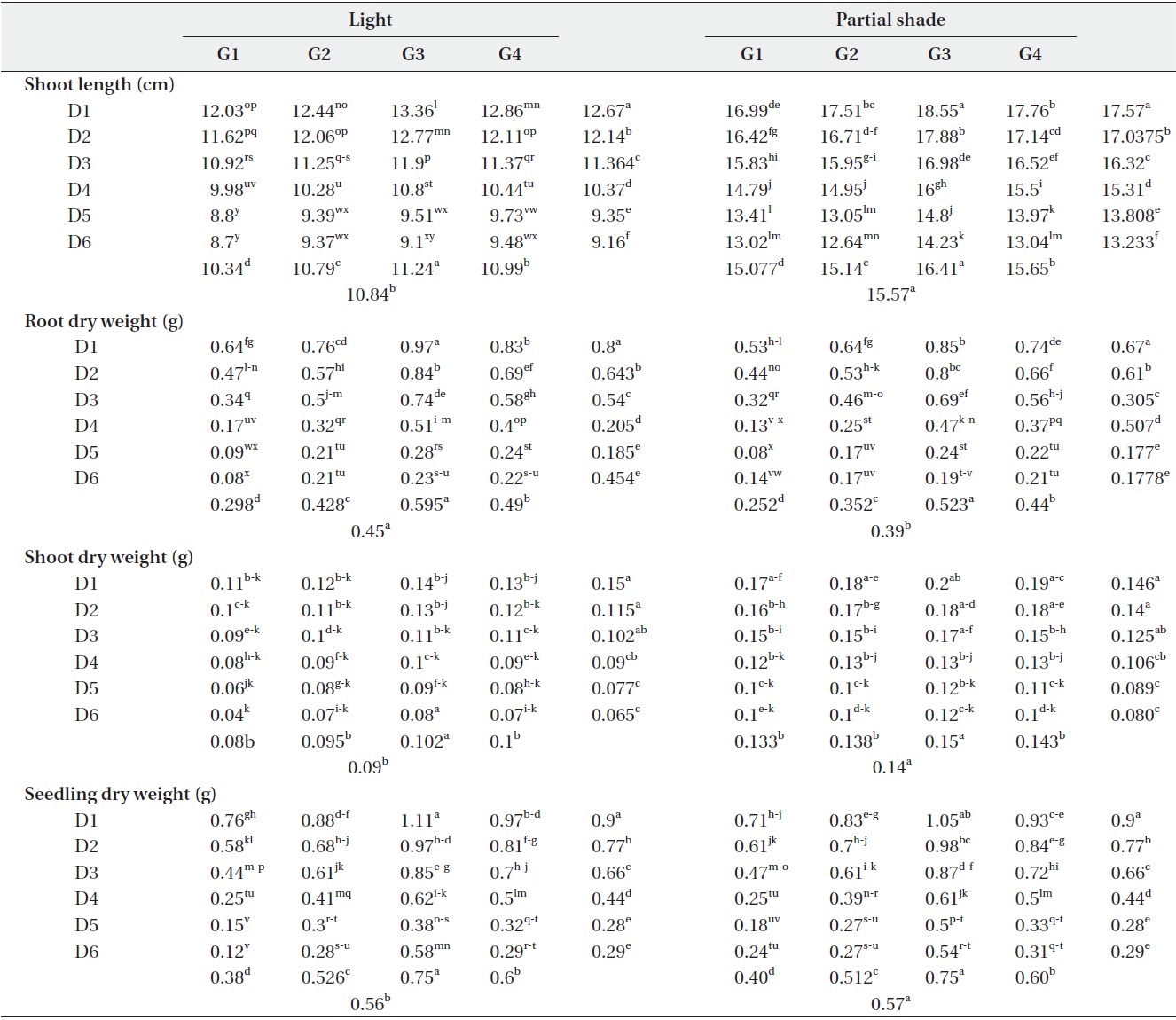
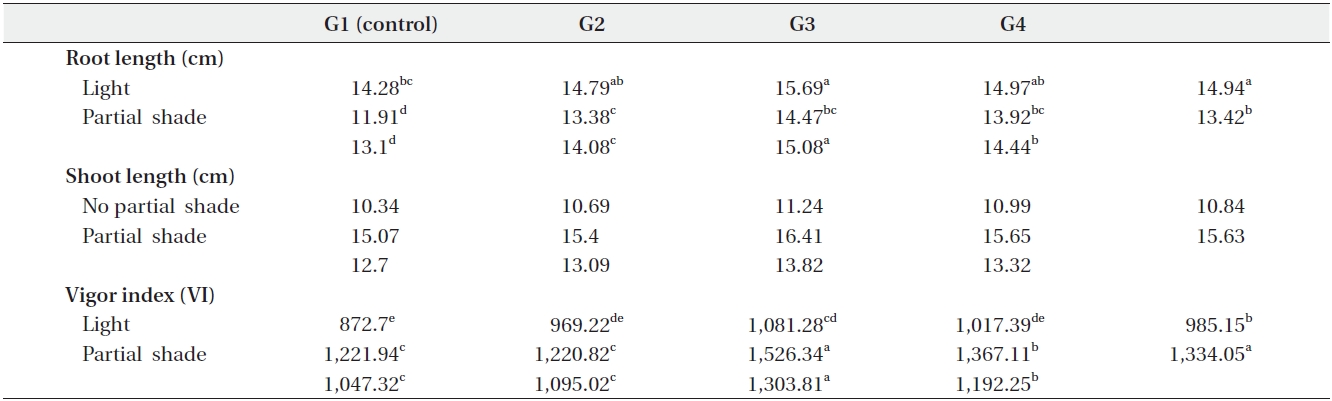
Shoot and root length were significantly (P < 0.01) affected by the interaction of growth medium and light (Table 1). For example, root length changed from 14.28cm in G1 (control) to 15.69 cm in G3 under light treatment. Also, it changed from 11.91 cm in G1 (control) to 14.47 cm in G3 under partial shade treatment (Table 3). In other words, root length increased by 1.41 cm (8.9%) and 2.26cm (15.6%) under light and partial shade treatments, respectively (Table 3). Shoot length was increased from 10.34 cm in G1 (control) to 11.24 cm under G3 light treatment. Moreover, it increased from 15.07 cm in G1 (control) to 16.41cm under G3 partial shade treatment (Table 3). In other words, root length increased 0.09 cm and 1.34 cm under light and partial shade treatments, respectively (Table 3). Interaction of drought stress and light treatments had significant (P < 0.01) effects on shoot length (Table 1). Shoot length decreased with increasing drought stress in light and partial shade. The shoot length changed from 12.67 cm in D1 (control) to 9.16 cm in D4 under light treatment. Also, it changed from 17.57 cm in D1 (control) to 13.23 cm in D4 under partial shade (Table 4).
Interaction of drought stress with growth medium as well as drought stress with light had significant (P < 0.01) effects on root and shoot dry weight (Table 1). Growth medium decreased drought stress effects on root and shoot dry weight more effectively under light treatments. Root and shoot dry weight changed from 0.64 g and 0.11 g to 0.97 g and 0.14 g in D1 and G3, respectively. Additionally, root and shoot dry weight changed from 0.08 g and 0.04 g to 0.23 g and 0.08 g in D6 and G3, respectively (Table 4). Growth medium decreased the effects of drought stress on root and shoot dry weight under the partial shade treatment. Root and shoot dry weight were increased from 0.53 g and 0.17g in D1 and G1 to 0.85 g and 0.2 g in D1 and G3. Also, root and shoot dry weight changed from 0.14 g and 0.1 g in D6 and G1 to 0.19 g and 0.12 g in D6 and G3 (Table 4).
Seedling dry weight was significantly (P < 0.01) affected by the interaction of drought stress with growth medium treatments (Table 1). Seedling dry weight decreased with the increase of drought stress, as the seedling dry weight changed from 0.76 g in G1, D1(control) to 0.12 g in G1, D6 (Table 4). Growth medium decreased the effects of drought stress, as seedling dry weight changed from 0.76 g in G1, D1 to 0.12 g in G1, D6. Seedling dry weight was 1.11 g in G3, D1 and 0.58 g in G3, D6 (Table 4). Seedling dry weight was significantly (P < 0.01) affected by the interaction of drought stress with light treatments (Table 1). Partial shade decreased the effects of drought stress. While seedling dry weight was 1.11 g in G3, D1 and 0.58 g in G3 D6 under light treatment, it was 1.05 g in G3, D1 and 0.4 g in G3, D6 under partial shade treatment (Table 4).
Vigor index was significantly (P < 0.01) affected by drought stress (Table 1). Vigor index decreased with increasing drought stress, as it was 1,462.31 in D1 (control) but just 1,036 in D6 (Table 2). Growth medium increased the vigor index from 1,047.32 in G1 to 1303.81 in G3 (Table 2).
There was significant (P < 0.01) difference in the vigor index between light treatments (Table1). Partial shade increased the vigor index, which was calculated to be 985.15 and 1,334.05 in no shade and partial shade respectively (Table 2).
The results show that Calotropis procrea L. is sensitive to drought stress in the emergence stage. Drought stress was found to reduce emergence percentage, shoot and root length (Table 2), as well as shoot, root and seedling dry weight (Table 3). Bazrafkan (2011) reported that Calotropis procrea L. is sensitive to drought in the germination stage. Ghaedi et al. (2009) reported that seeds of Haloxylon aphyllum L. as a halophyte is sensitive to drought and salinity in germination stage. Ramazani et al. (2009) reported similar findings about (Capparis spinosa L.). Similar findings have also been reported for Atriplex griffithii var. stoksii (Khan and Rizvi 1994), Kalidium capsicum (Tobe et al. 2000), Calotropis procera L (Taghvaei et al. 2012). Drought stress decreases soil osmotic potential, which in turn, decreases the water uptake of seeds, thereby inhibiting their emergence (Katembe et al. 1998). Emergence percentage, shoot length and root length were significantly affected by light treatments (Table 1). Duchesne et al. (2000) reported the positive effect of partial shade on the early establishment of eastern white pine seedlings. Taghvaei (2010) reported that partial shade treatments had significant effects on emergence percentages and rates, shoot length, shoot dry weight, root dry weight, leaf area, and Chlorophyll content of Quercus brantii L. Partial shade could increase shoot length (Qian and Engelke 1999). Ray and Brown (1995) reported that emergence success in dry forest in the US Virgin Islands was approximately equal under conditions of partial- and no-shade. Growth medium had positive effects on all characteristics of C. procera L. seedlings. The results showed that emergence percentage, shoot and root length, vigor index, as well as shoot and root dry weight increased with the application of lighter-textured soil (Table 2). Similar results were reported for Salsola vermiculata (Guma and Padron-Mederos et al. 2010). Damizadeh (2004) reported that light textured sandy and sandy loam is the most appropriate growth medium for Capparis decidua seedling survival. It seems that it has positive effects on ventilation and drainage. Abo-Rezq et al. (2009) found out that a mixture of perlite with peatmoss and sand is the best growing medium for Chlorophytum comcosum and Lycopersicom esculentum cv. Using sand improves the physical properties of a growing media and improves drainage (Wilson and Stoffella 2006). CONCLUSION
Calotropis sp. is a species adapted to the deserts of southern Iran (Sadeghian et al. 2010). Calotropis sp. is sensitive to drought (Bazrafkan 2011) and salinity (Taghvaei et al. 2012). Climate change has increased periods of drought innmany regions of Iran, and additional anthropogenic influences, including farming practices, have reduced the density of Calotropis sp. in these areas, even though many seeds are produced. In Calotropis sp., root initiation and growth ceased at soil osmotic potentials between -0.4 and -0.6 MPa (Bazafkan 2011). Under these dry conditions, the germination and early establishment of many species, especially Calotropis sp., must occur during the wet season or at the end of winter, when water is more available. Evaporation from the top layer of soil in early spring decreases germination of species seeds. Partial shade is modified at the ground level with a reduction of temperature (Rincon and Huante 1993). Partial shade treatment seems to result in sufficient alteration of the microclimate for a significant improvement in emergence and establishment (Gehlhausen et al. 2000). In this experiment, partial shade treatment was found to have an influence on seedling emergence and seedling growth. Partial shade treatment increased shoot length and vigor index, but decreased emergence percentage. Partial shade protect soil moisture, and improves the field emergence of seeds (Chalton et al. 1986). Consequently, light quality becomes a limiting factor for their survival and growth (Crow 1988). Although low light as a dominant environmental factor limits seedling establishment under partial shade treatment, it may increase root growth. Root regeneration and root growth are very sensitive to soil moisture stress (Lopez-Barrera and Newton 2005). So, to increase germination, the growth medium which will retain rain water for a longer time must be considered. Sand+perlite media increased root characteristics, and this may imply that this medium had better water retention properties. The use of perlite allows plants to optimize water and oxygen holding, and could affect plant growth and the development of roots and shoots directly or indirectly, as perlite increases aeration and water holding capacity of soil (Leakey et al. 1990, Ofori et al. 1996). Therefore, to improve the establishment of milkweed in dry semi-arid regions, it is suggested that seeds be planted under a canopy of native plants as nurse plant. Because organic matter is derived from litter under the canopy of these plants, rain water will be retained with greater efficiency, thus improving germination and emergence percentages. The partial shade of nurse plants result in the improvement of the microclimate, which can be sufficient to maintain young seedlings against a variety of effects, including excess radiation and grazing in arid and semi-arid lands.