Thisexperiment was conducted to investigate the effects of the addition of wintering waterfowl’s feces prior to planting rice on nutrient dynamics, rice growth, and decomposition of rice straws in a controlled mesocosm. Waterfowl’s feces and rice straws were placed on paddy soils in a mesocosm and the water level maintained at 5 cm. The amounts of supplied feces were 0 (control), 222, and 444 g/m2. While the addition of feces showed no immediate effects, nutrients in the surface water increased in the month following treatment. Nutrients increased to a greater degree in the treatments with more feces added. Simultaneously, the decomposition of rice straws was promoted, indicating that more nutrients would be made available over time. The rice showed high productivity in the period during which nutrient level was increased, when rice needs more nutrients for the tillering stage. Therefore, the wintering waterfowl’s feces could be associated with increased productivity during the growing season of rice through promoting nutrient supply and rapid decomposition of rice residue.
A large colony of birds can have a profound effect on the environment, and a large-scale bird migration has significance in nutrient cycling. In particular, bird feces provide an outside nutrient source to environments in which they dwell (Manny et al. 1994, Tomassen et al. 2005, Chaichana et al. 2010). Subsequently, continuous inhabitation of birds can have an important impact on the plant community in their habitats (Ishida 1997, Nam et al. 2008).
Many migratory waterfowl inhabit wetland areas and form large flocks. The environment and plants around the wetlands they inhabit can be influenced by their excretion (Manny et al. 1994). South Korea is home to many wintering habitats of migratory waterfowl (Lee et al. 2000, 2001). In particular, the Han River estuary in winter 2008 recorded about 100,000 birds of 53 different species, which were dominated by waterfowl such as the white-fronted goose and bean goose (Kang et al. 2008). Many species of wintering bird are observed in rice paddy fields near the estuary. The paddy fields in winter supply wintering waterfowl with a foraging and resting place. They stay form around October to March. As large flocks of waterfowl stay in the paddy fields in winter, the addition of their feces can be influential on the nutrient status for the growth of rice in the following spring.
However, there is a paucity of studies analyzing the impacts that the feces of wintering waterfowl have on rice growth in the locations where they forage and live during the winter months. Although there are studies on waterfowl’s feces, the research has been conducted in lakes and ponds of deep water (Unckless and Makarewicz 2007, Nakamura et al. 2010), which is a very different environment from paddy fields, wetlands composed of shallow water. Other studies on paddy fields looked at the effect of added feces during the growth period (Isobe et al. 2005, Chengfang et al. 2008). However, the wintering waterfowl’s feces are added to the paddy fields in the Han River estuary only during the early season before rice growth. Thus, we have on information about the effect of feces supplied by wintering waterfowl on the environment of paddy fields and on rice growth during the growing season.
The addition of fecal matter can cause a change in the decomposition rate of the previous season’s plant matter, thereby increasing the amount of available nutrients to new plants over time (Kelly and Henderson 1978, Elwood et al. 1981). Plant residues are an important source of nutrients in the soil, indicating that decomposition of rice straws could be an influential factor to nutrient cycling in paddy fields. In particular, the synchronization of nutrient release from plant residues and uptake by plants is an important matter affecting the growth of rice plants (Villegas-Pangga et al. 2000, Rejmankova 2011).
The purpose of this study was to investigate the effects of waterfowl’s feces, added to paddy fields before rice growth, on the nutrient status of the paddy field, rice growth, and the decomposition of rice straws during the period of rice plant growth. A greenhouse experiment was designed to simulate the fecal additions by waterfowl before rice growth in a variable-controlled environment.
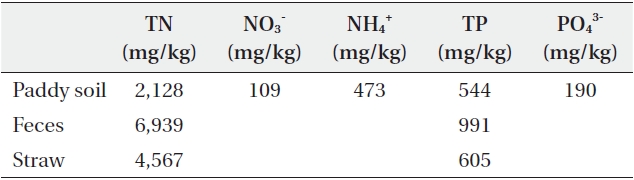
[Table 1.] Analysis of paddy soil, feces and straw used in the experiment
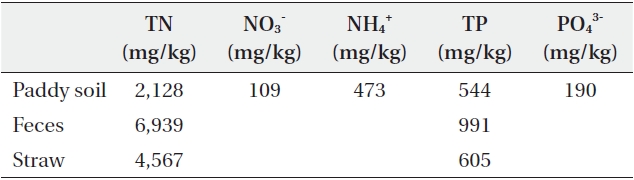
Analysis of paddy soil, feces and straw used in the experiment
[Table 2.] Input amount of waterfowl feces
Input amount of waterfowl feces
The experiment was conducted in a greenhouse at Seoul National University from 27 May 2009 to 13 October 2009. Maximum and minimum temperatures fluctuated between 38℃ and 15℃ and between 21℃ and 8℃, respectively (Fig. 1). Plastic pots (50 × 36 × 30 cm) were used for growing rice and filled with 25.41 kg of soil, collected from harvested paddy fields and sieved with 2 mm mesh, to a depth of 10 cm (Table 1). The water level was maintained with tap water at 5 cm above the soil surface.
Three treatments were studied: fecal additions of 0, 40, and 80 g per pot (Table 2). The addition of 40 and 80 g of fecal matter was calculated to result in a distribution of 222 and 444 g/m2, respectively. We used this amount based on the following data. Daily feces amounts of goose and duck are 32.76 g and 15.07 g, respectively (Manny et al. 1994) and they stay in paddy field for 7.9% and 6.7% of a day, respectively (Pae et al. 1995) from mid-August to mid-March (about 150 days). This makes 151 g to 388 g feces m-2 y-1. Unlike other studies in which experimental protocol included the continuous addition of feces throughout the growth period, we added the experimental amount of feces to the pots only at the beginning of the experiment. The feces for the experiment were collected from the paddy fields around the Han River in February, and total nitrogen (TN) and total phosphorus (TP) were analyzed immediately (Table 1). The feces were stored in a refrigerator at 4℃ and pulverized after air-drying. Each treatment had four replicates, and a total of twelve pots are arranged so that pots receiving the same treatment were not adjacent to one another.
Rice straws were air-dried at room temperature and the rice straws to be used in litter bags were dried at 60℃ for 48 h. The rice straws were then cut to a length of 5 cm and 5 g were placed into each nylon litter bag (5 cm × 5 cm) with 1 mm mesh. Eight litter bags were put into each pot, to which rice seedlings at the two-leaf stage were transplanted. After the harvest, the litter bags were collected and TN and TP were analyzed.
A sample of 100 mL of water was gathered from each pot every two weeks for the first month, then once a month for the duration of the study. The sampled water was filtered and stored at -20℃ until analysis of available nitrogen and phosphorus was performed. Nitrate (NO3-), ammonium (NH4+), and phosphate (PO43-) were analyzed by the hydrazine method (Kamphake et al. 1967), indophenol method (Liddicoat et al. 1975) and ascorbic acid reduction method (Solorzano 1969), respectively.
Soil samples were taken at the beginning and end of the experiment. TN and TP in the soil samples were analyzed at the National Instrumentation Center for Environmental Management at Seoul National University. Available nitrogen and phosphorus were extracted using KCl extraction and Truog’s method, respectively (Kim et al. 2004). Nitrate, ammonium and phosphate in the extracted solutions were analyzed using the same methods used for water sample analysis. To investigate whether a change in rice growth occurred, the number and height of shoots in each pot were measured and the average and total sum of shoot length calculated. After conducting the first measurement at the end of the first week, measurements were conducted every two weeks. At the end of the experiment, the rice was collected from each pot for measurement of total biomass. Shoots and roots were collected at the same time. Collected roots were washed with tap water and dried at 60℃ for 48 h, then weighed.
The litter bags, used to measure the decomposition rate of the rice straws, were collected at 0, 1, 3, 6, 10, 13, 17, and 21 weeks. Collected litter bags were carefully washed and dried at 60℃ for 48 h, weighed, and the weight loss ratio calculated.
A one-way ANOVA (
>
Available nutrients in the surface water
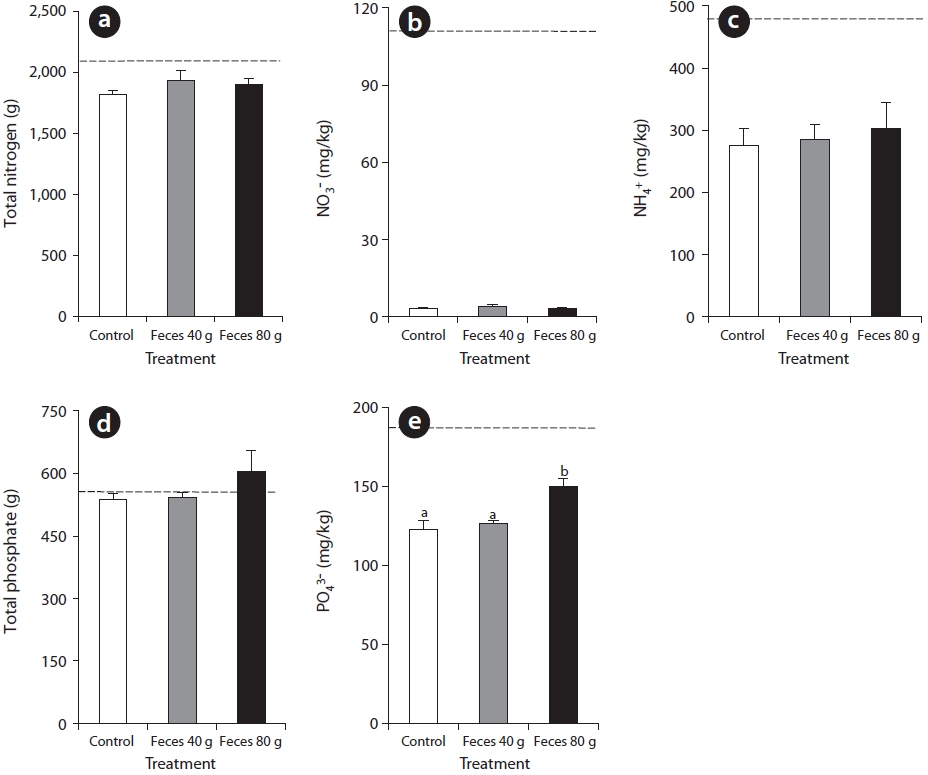
Nutrients in the surface water initially increased at a high rate, then appeared at decreasing rate over time with only a small quantity left at the completion of the study (Fig. 2). The plots containing 80 g of feces were found to contain a higher nutrient content compared with other plots during the period of nutrient increase. Nitrate increased rapidly at the fourth week and then largely decreased at the eighth week (Fig. 2a). Nitrate content in the plots containing 80 g of feces was significantly different from other plots. Ammonium content was highest in the second week under all experimental conditions without a significant difference among the different plots. Ammonium content in the plots containing 80 g of feces was significantly different from control at the fourth week, and ammonium content in the plots containing 40 g of feces was significantly different from that found in the control at the eighth week (Fig. 2b). Phosphate increased at the fourth week and immediately decreased, being significantly different in the plots containing 80 g of feces from all other plots in the study (Fig. 2c).
>
Available nutrients in the soil
Each nutrient in the soil decreased differentially by the end of the study (Fig. 3). The content of TN was 2,128 mg/kg at week 0. After harvest, the control plots, plots containing 40 g of feces, and plots containing 80 g of feces had 1,821 ± 32 mg/kg, 1,930 ± 83 mg/kg, and 1,904 ± 46 mg/kg, respectively (Fig. 3a). The initial nitrate content in all plots was 109 mg/kg. After the harvest, nitrate content in the control plots, plots containing 40 g of feces, and plots containing 80 g of feces was 3.3 ± 0.4 mg/kg, 4.1 ± 0.7 mg/kg, and 3.3 ± 0.4 mg/kg, respectively, indicating that the available nitrate was almost entirely exhausted (Fig. 3b). The initial content of ammonium in all plots was 473 mg/kg and decreased to 276 ± 27 mg/kg in the control, 285 ± 25 mg/kg in plots containing 40 g of feces, and 302 ± 43 mg/kg in the plots containing 80 g of feces, without a
significant difference in the amount of ammonium consumed among the plots (Fig. 3c). TP and phosphates in the soil showed a slight decrease over the period of the experiment. TP was 544 mg/kg at the beginning of the experiment. After the harvest, TP in the control plots and the plots containing 40g of feces was 537 ± 14 mg/kg and 543 ± 12 mg/kg, respectively. In contrast, the TP content in the plots containing 80 g of feces increased to 605 ± 49 mg/kg (Fig. 3d). The initial content of phosphates was 190 mg/kg at the beginning of the experiment. After the harvest, phosphate content in the control plots, plots containing 40 g of feces, and the plots containing 80 g of feces was 123 ± 6 mg/kg, 127 ± 2 mg/kg, and 150 ± 5 mg/kg, respectively (Fig. 3e).
>
Effect of different amount of feces on height and biomass of rice
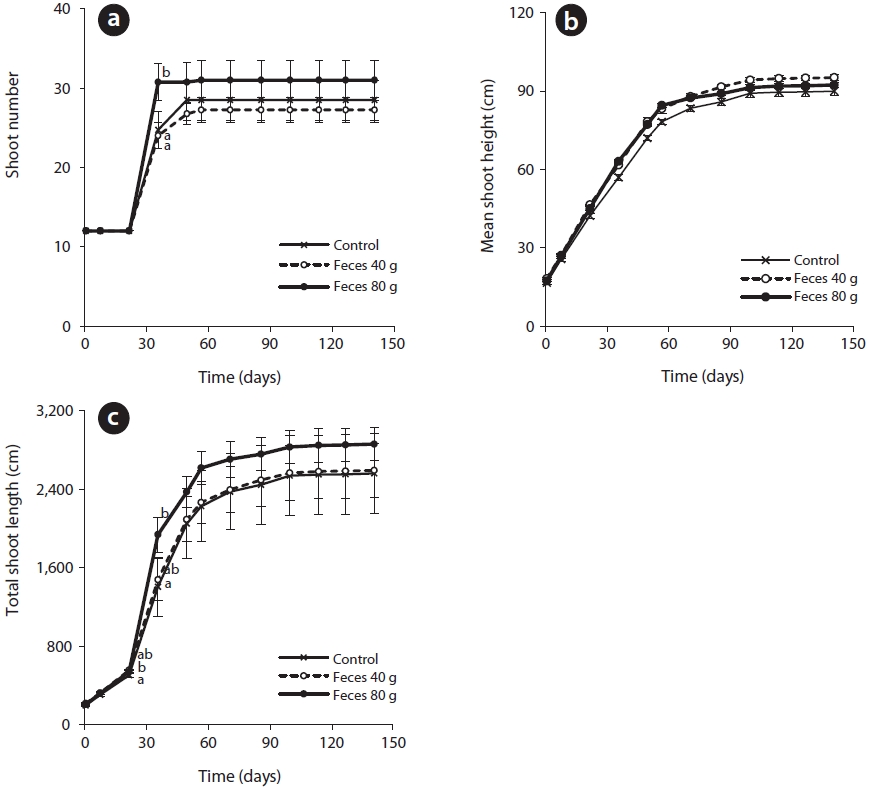
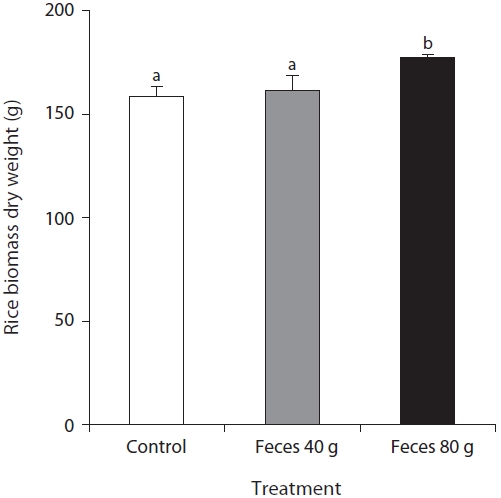
The rice showed a sharp increase in growth at the beginning of the tillering period at the fifth week of the experiment, and the total of the shoot height increased in the presence of added feces (Fig. 4). The shoot number began to differ among the experimental conditions at the tillering period. The plots containing 80 g of feces had the largest number of shoots, 31 ± 1 shoots. There was no significant difference between the control, 28 ± 3 shoots, and the plots containing 40 g of feces, 27 ± 2 shoots. The plots containing 40 g of feces had the highest average of shoot height, 95 ± 1 cm, followed by the shoot height in the plots containing 80 g of feces, 92± 1 cm, and the control, 90 ± 1 cm. The total of shoot height showed a significant difference between the plots with feces and without feces at the third week, and between the control and the plots containing 80 g of feces at the sixth week. Following the sixth week, although shoot height among the different plots were not statistically different, the average value showed an increasing trend in the presence of feces. At the experiment’s end, the average value of shoot height in the control plots, plots containing 40 g of feces, and plots containing 80 g of feces were 2,563 ± 203 cm, 2,671 ± 137 cm, 2,861 ± 83 cm, respectively. The biomass of rice showed an increasing trend in the presence of feces. The dried biomasses collected from the control plots and plots containing 40 g of feces were 159 ± 4.7 g and 162 ± 6.5 g, respectively. The plots containing 80 g of feces had a significantly different biomass, 177 ± 1.4 g, compared to both the control and plots containing 40 g of feces.
>
Decomposition rate of straw with different amount of feces
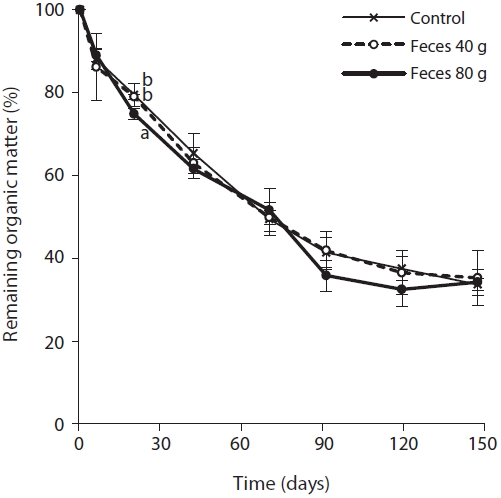
Although the decomposition of the rice straw showed a significant difference among the plots during the period from the first week to the sixth week, there was no significant difference in the final analysis. The decomposition rate in the plots containing 80 g of feces was significantly higher compared to the control and plots containing 40 g of feces at the third week. After the third week, the decomposition rates in the control and the plots containing 40 g of feces changed in a similar trend, and the plots containing 80 g of feces had a relatively higher rate of decomposition but without statistical significance.
We found that addition of waterfowl’s fecal matter had an effect on the water and soil chemistry, the decomposition rate of rice straws, and influenced plant growth. In particular, the effect was most distinct in the plots containing 80 g of feces. The content of nitrate and phosphate in the surface water increased rapidly at the fourth week in all treatments. During this time, the nutrient content in the surface water was different among the treatments, as well as, the decomposition rate of rice straws and the total sum of shoot height. However, from then on, as the contents of nitrate and phosphate decreased with rice growth, the difference among the treatments ceased to exist. However, the final biomass measurement from the plots containing 80 g of feces was significantly greater when compared with other treatments.
The fecal impacts appeared at about one month from addition in the present study (Fig. 2). The time over which nutrient content was increased in the surface water did not depend on the amount of feces in each treatment, but the quantities of nutrients present did. Natural environments influence the time during which feces begin to have an effect on nutrient content in bodies of water. Ganning and Wulff (1969) and Pettigrew et al. (1998) reported that the rapid dissolution of bird feces influenced nutrient content of a body of water over a short period. In contrast, Unckless and Makarewicz (2007) suggested that based on the results of a mesocosm experiment, the impact of bird feces would not become instantly evident and the additional nutrient input from the feces would settle quickly into the sediment. Nakamura et al. (2010) reported that the size of the wintering waterfowl population immediately influenced the TN content, but the TP content peaked one month after the wintering birds had left. Availablity of nutrients and nutrient content of surface water are subject to many variables such as biological consumption, absorption into sediments, decomposition, and mixing events (Unckless and Makarewicz 2007, Nakamura et al. 2010). In the present experiment in which the rice was grown in pots with soil from paddy fields, the contents of available nutrients in the surface water decreased at first then increased after one month.
Although Ganning and Wulff (1969) and Nakamura et al. (2010) argued that the nutrients from feces were primarily taken up by algae, their results were based on the experiment in rockpools and ponds, respectively. However, the increased nutrient content from fecal additions was likely used by the rice plants in the present study. Nutrient content decreased with rice growth, and the total sum of shoot height and shoot number increased with the nutrient amount available during the period over which the nutrient content increased (Figs. 2 and 4). These results support the idea that additional nutrient input from feces could be directly absorbed by rice plants and influence rice growth.
The tillering stage in the present study occurred around the same time that the nutrient content in the surface water was significantly different among the treatments (Figs. 2 and 4a). The tillering stage is important in that it increases the leaf area for photosynthesis and the ear number. Increased nutrient availability tends to stimulate the tillering of rice (Yoshida and Hayakawa 1970, Yoshida 1981). The present study also supports the idea that the nutrient increase from bird feces is closely connected with the increased number of rice shoots. That is, the plots containing 80 g of feces and subsequent statistically significant increase in nutrient content showed a relatively greater number of rice shoots than the control and plots containing 40 g of feces which did not exhibit a statistically different increase in nutrient content.
The addition of fecal matter on straw decomposition rate resulted in a timely contribution to the tillering phase of rice plants. At the fourth week, the decomposition rate, as well as the nitrate and phosphate present in the surface water, were different among treatments (Figs. 2 and 5). This supports the concept that the presence of additional nutrients due to the presence of feces tends to positively stimulate the decomposition of rice straws (Elwood et al. 1981). The rapid decomposition rate of rice straws indicates an increase in available nutrients. Therefore, the addition of fecal matter leads to an increase nutrient supply by stimulating the decomposition of fallen rice straws. In addition, the tillering stage of rice occurred at about
the same time as decomposition was stimulated, which might have worked in the rice plants’ favor during the tillering stage despite there being no difference in the rate of decomposition at the end of the experiment.
Although the shoot number was similar between the control and the plots containing 40 g of feces, the average of rice biomass and soil nutrients was higher in the plots containing 40 g of feces than in the control plots (Figs. 3 and 6). In the case of TP in the soil, the final content was not different from the initial content in the plots containing 40 g of feces but was higher in the plots containing 80 g of feces (Fig. 3d). These results indicate that despite not having an instantaneous impact, nutrients from waterfowl’s feces flowing into rice paddy fields have the potential to influence rice growth over the long term. Therefore, if rice paddy fields can be increasingly used as a habitat for wintering waterfowl, this may have a nutritionally favorable impact on conditions for rice plant growth in these fields.
We found that waterfowl’s feces positively impact rice plant growth. Thus, rice plant growth could be promoted by the presence of more wintering waterfowl populations in paddy fields. Although Lee et al. (2006) argued that many waterfowl could cause damage to the crops, our results support the concept that the inhabitation of paddy fields by wintering waterfowl has a favorable impact on rice growth. Therefore, our discussion lends credence to the idea that conservation and creation of more paddy field habitats for wintering waterfowl may increase the agricultural and ecological value of these fields.
The present study investigated the impact of birds’
feces on nutrient availability under controlled conditions. With regard to nutrient cycling, the conditions in the greenhouse environment of this study were different from those found in the real environment of paddy fields. In paddy fields, the inflow and outflow of nutrients occur via complicated and highly variable routes (Reddy 1982, Cho et al. 2000). Wintering waterfowl can have additional influences on rice growth by the feeding behavior (Cargill and Jefferies 1984, van Groenigen et al. 2003). Therefore, although the present study showed the potential of wintering waterfowl’s feces to favorably impact rice growth, further experiments on paddy fields are needed. These will help deepen our understanding of the ecological impact of waterfowl flocks on the ecosystem of paddy fields.
To investigate the effects of wintering waterfowl’s feces on nutrient fluctuation, rice growth, and decomposition of rice straws, their feces were added to paddy soils for rice transplanting. In this study, effects were not immediately observed. However, the nutrient content in the surface water increased over the month following the addition of feces. During this period, the greater the amount of feces added, the greater the increase in available nutrients. In addition, the decomposition of rice straws was facilitated, supplying even more nutrients to the experimental environment. The nutrient level rapidly decreased during the tillering stage when the rice needs more nutrients. The treatments with more available nutrients exhibited a greater number of rice shoots, resulting in an overall increase in production. In the natural environment, the potential impacts of waterfowl’s feces are affected by rice decomposition and seasonality in ways that are not entirely predictable. In conclusion, the addition of waterfowl’s feces has a positive impact on rice growth. Our results support the concept that increased waterfowl inhabitation of paddy fields can positively influence rice plant growth.