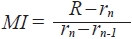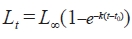Blackfin flounder Glyptocephalus stelleri is distributed in the East Sea and South Sea of Korea, Japan, Sakhalin, Tatar Strait, South Kurils Strait, and Bering Sea (Matarese et al., 1989; Choi et al., 2002). This benthic fish, mostly inhabits depths of more than 300 m moving to the coast water during its spawning season (Choi et al, 2002; Fedorov et al., 2003; National Fisheries Research and Development Institute, 2004).
G. stelleri are caught by gill nets, longlines, eastern sea Danish seines and bottom trawls in the East Sea, and the catch ratio of this fish reaches up to 54.9% of the total flounder catches in the East Sea (National Fisheries Research and Development Institute, 2010). Recent increases in the flounder catches are related to increases in the catch of G. stelleri, Recently G. stelleri was selected as one of the species for a stock rebuilding project in the East Sea, and research on its ecology has been promoted. There have been many studies on G. stelleri, including studies on its age and growth (Hashimoto, 1953; Ishida and Kitakata, 1953), larvae (Okiyama, 1963), spawning ecology (Ivankova, 1974), feeding (Hayase and Hamai, 1974; Pushchina, 2000), and geographic distribution (Shvydkii and Vdovin, 2001; Tokranov, 2008). Studies of the biology and ecology of G. stelleri in Korean waters have included research on growth (Lee, 2008) and reproduction (Cha et al., 2008), and the species has also been reported in wider studies of fish (Park et al., 2007; Yoon et al., 2008; Lee, 2011).
Research conducted in Korea on Pleuronectidae species includes studies of the ages and growths of Limanda herzensteini (Choi et al., 1986), Verasper variegatus (Jeon et al., 1996), Pleuronectes yokohamae (Park, 1997)and Kareius bicoloratus (Jun and Im, 2004).
The present study analyzed data on the age and growth of G. stelleri. These results can be used to provide a resource management plan for the continuous exploitation of G. stelleri resources.
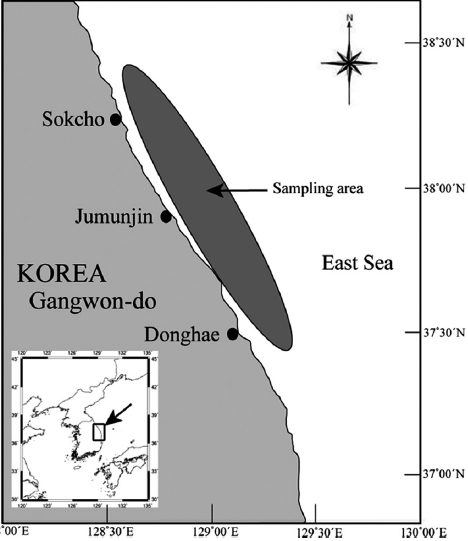
The G. stelleri samples were caught by the eastern sea Danish seine fishery in the sea adjacent to Gangwon-do, Korea, in 2007 (Fig. 1). Monthly samples were collected from landing ports at Mukho and Sokcho. The fish were sexed and total lengths (TL) and total weight (TW) were maeasured to the nearest 0.1 cm and 0.1 g, respectively.
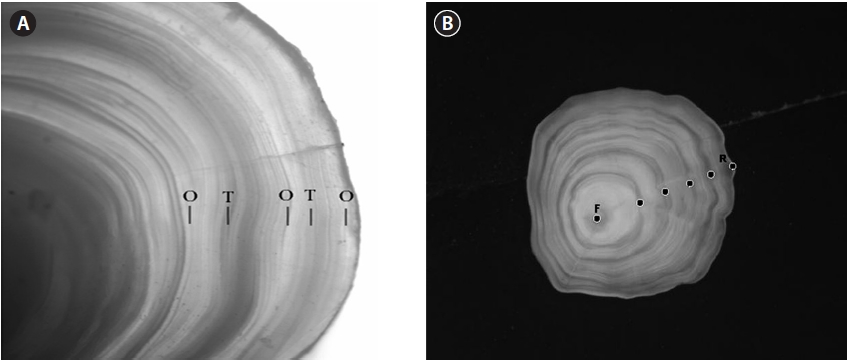
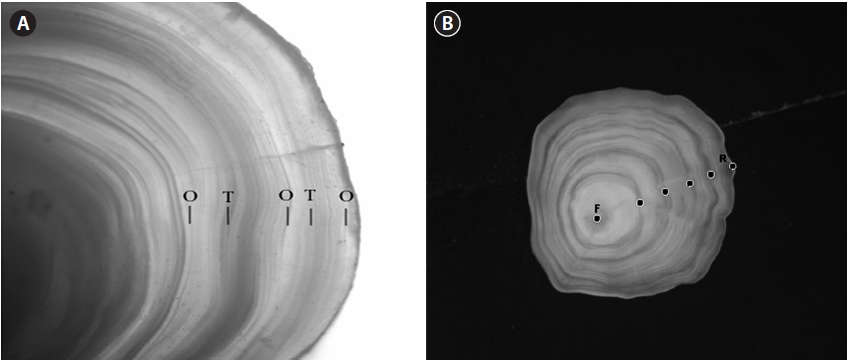
A pair of left/right otoliths was extracted and, after the removal of organic matters on the surface, they were dried and treated for easy observation by grinding close to the nucleus with sandpaper. Translucent and opaque zones appeared alternately in the otolith (Fig. 2A). Annuli were found at the boundary between opaque and translucent zones, and these were analyzed on a PC monitor (with the image analyzer package iSolution Lite, IMT i-Solutin Inc., Daejeon, Korea) connected to a dissecting microscope.
The otolith radius (R) was measured along the longest axis from the otolith center to the posterior margin, and the ring radius (rn) was measured in μm from the otolith center to each annulus (Fig. 2B).
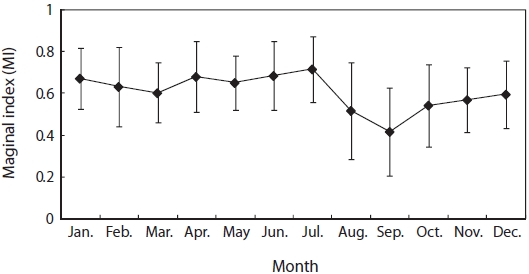
To determine the period of annulus formation, the monthly variation in the marginal index (MI) was calculated by the following equation:
R is otolith radius and rn is the ring radius to the n-th annulus.
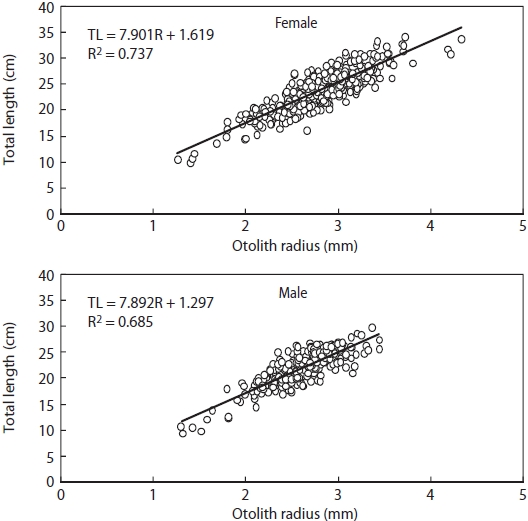
Relationship between TW and TL was estimated by TW=aTLb, where a and b are parameters. Relationship between TL and otolith radius (R) was estimated by linear regression analysis.
Growth in length of G. stelleri was obtained from the growth function of Von Bertalanffy (1938). The parameters obtained by the Walford method (Walford, 1946) were used as the initial values of the growth formula, which was presumed by nonlinear regression (Microsoft Excel solver routine):
Lt is the TL at age t, L∞ is the theoretical maximum length, K is the growth coefficient, and t0 is the theoretical age at length zero.
An analysis of covariance (ANCOVA) was used to compare weight-length relationships between sexes, and growth difference between males and females for each age was examined by t-tests in the SPSS version 12.0 (SPSS INC., Chicago, IL, USA).
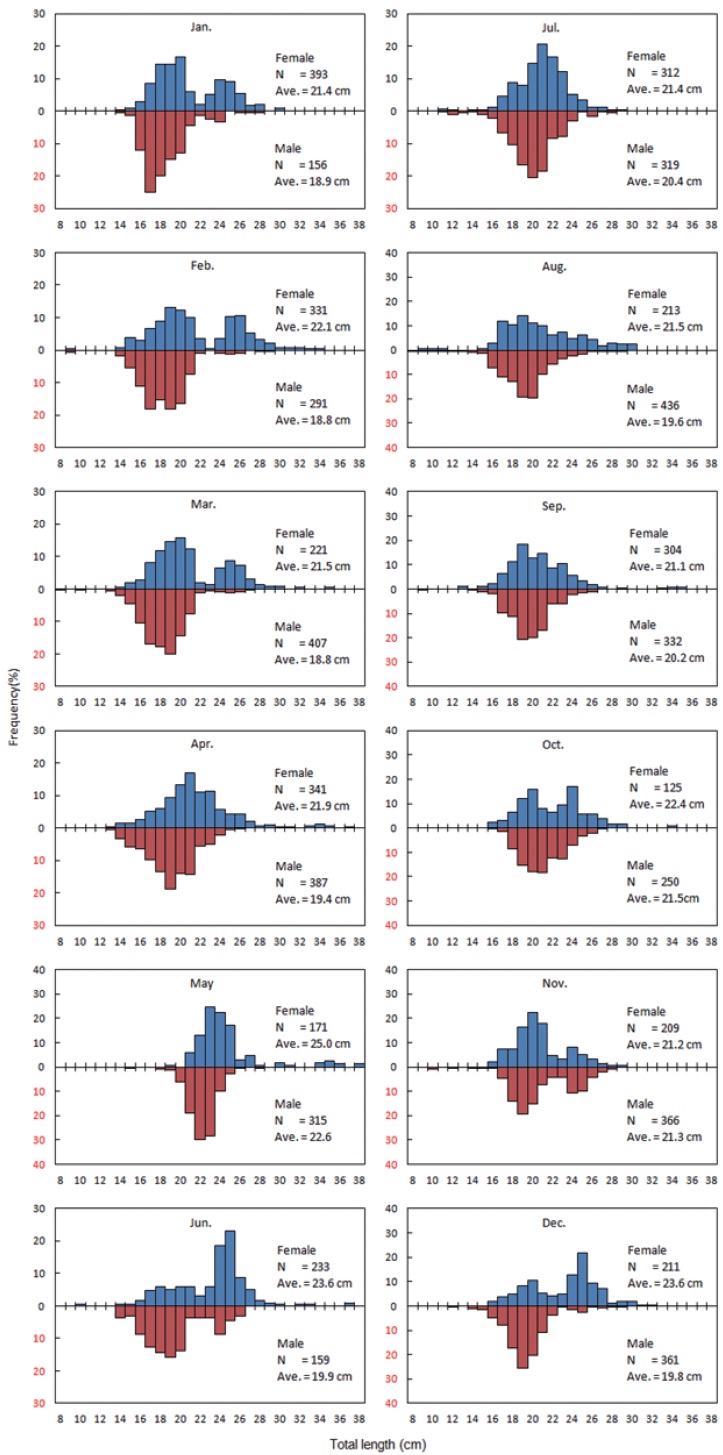
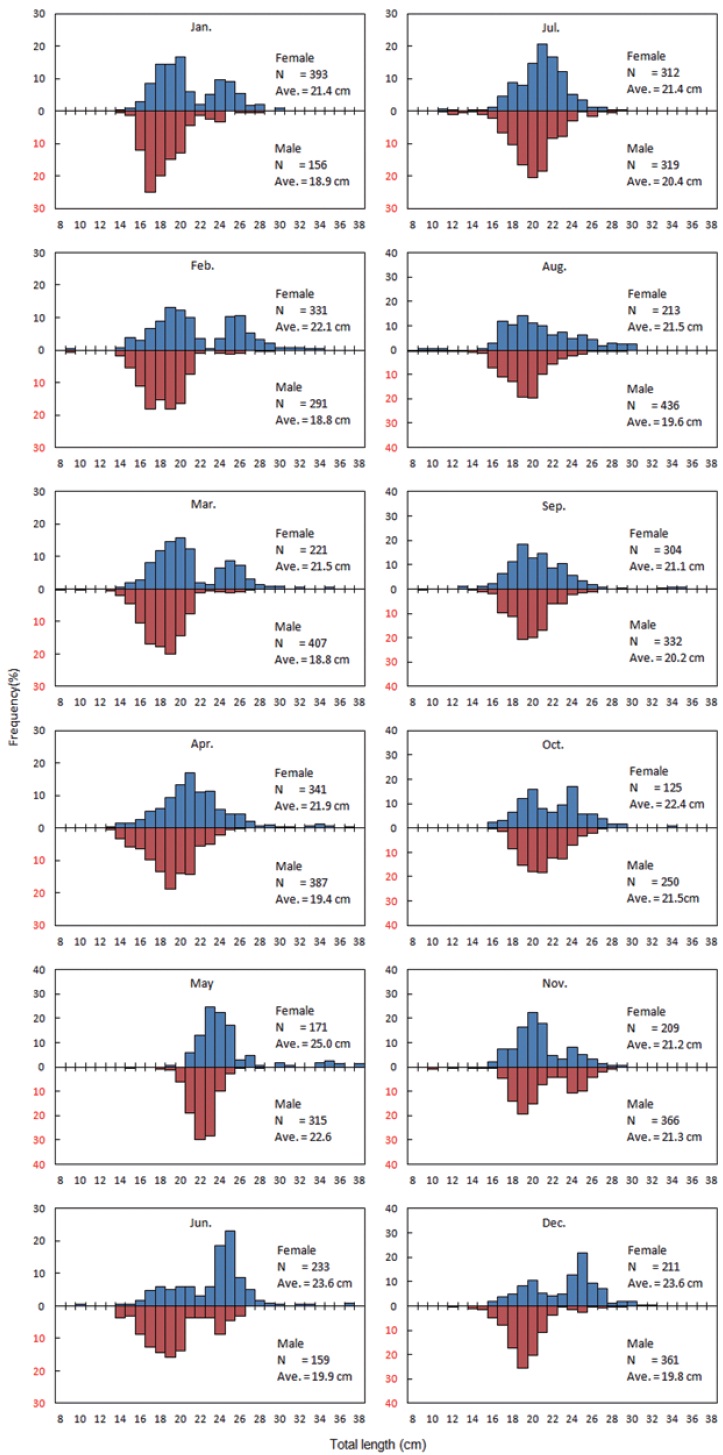
The total number of G. stelleri collected in the survey was 6,843, of which 3,064 were females and 3,779 males. TL was 9.5 - 38.1 cm for females and 8.6 - 29.5 cm for males, showing
that females are longer than males (Fig. 3).
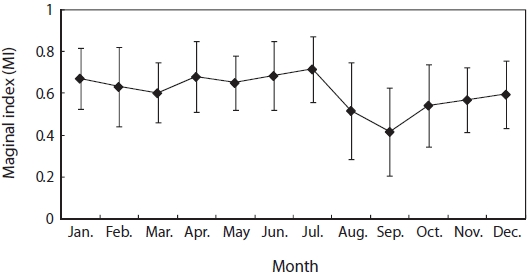
Monthly variation in MI was highest in July, low in August and September, showing an increasing trend from October (Fig. 4). The annuli of G. stelleri were formed from around September at the boundary between opaque and translucent zones in the otolith.
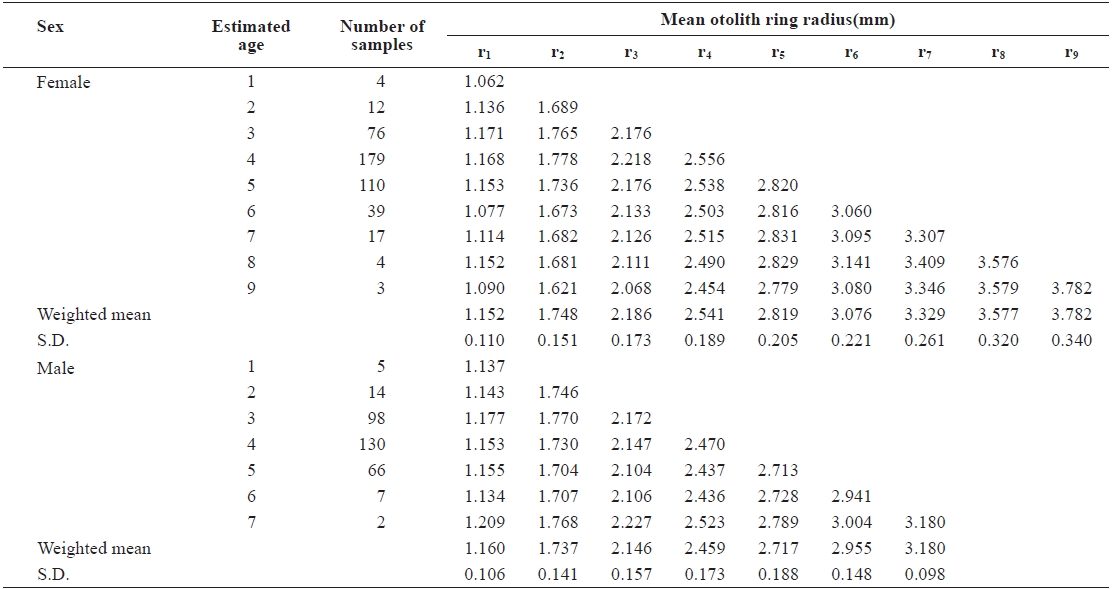
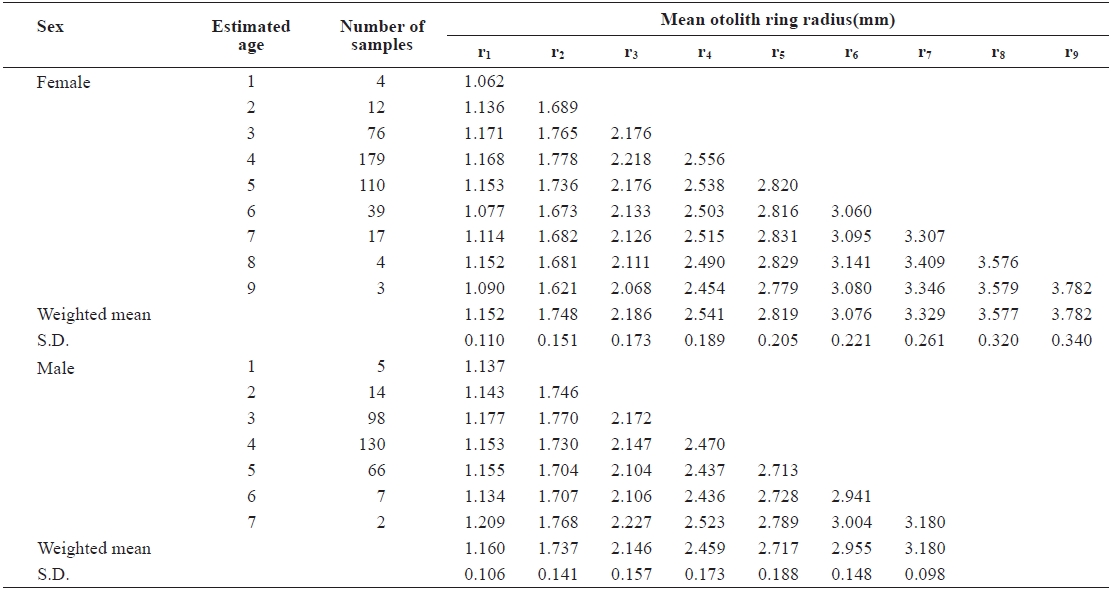
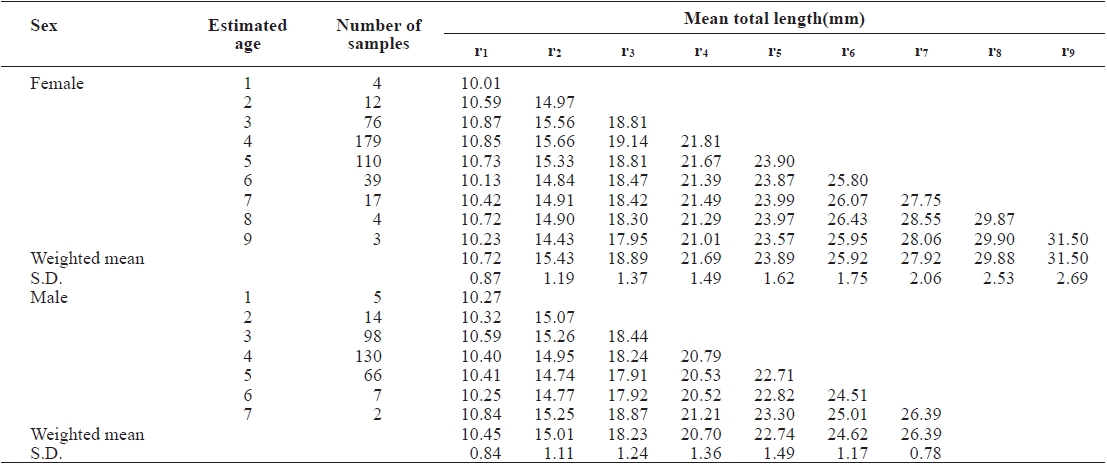
The average ring radius from the otolith center to each annulus is shown in Table 1. It ranged from r1 = 1.152 ± 0.110 mm to r9 = 3.782 ± 0.340 mm in females and from r1 = 1.160 ± 0.106 mm to r7 = 3.180 ± 0.098 mm in males. The average ring radius female was apparently longer than for males, for each age group.
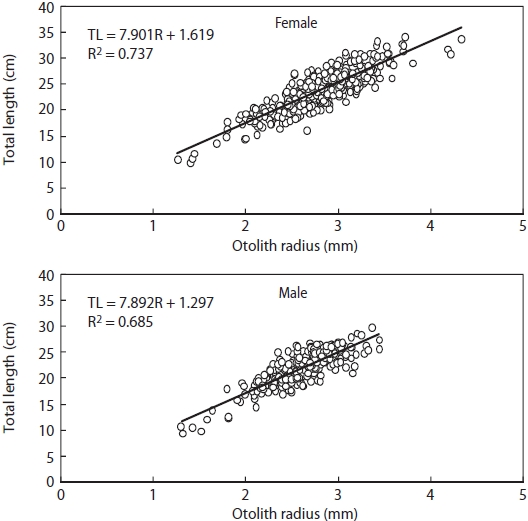
Relationship between R and TL is shown in Fig. 5. There were significant differences in this relationship between males and females (ANCOVA, F = 5.670, P < 0.05), with TL = 7.901R + 1.619 (r2 = 0.737) for females and TL = 7.892R + 1.297 (r2 = 0.685) for males.
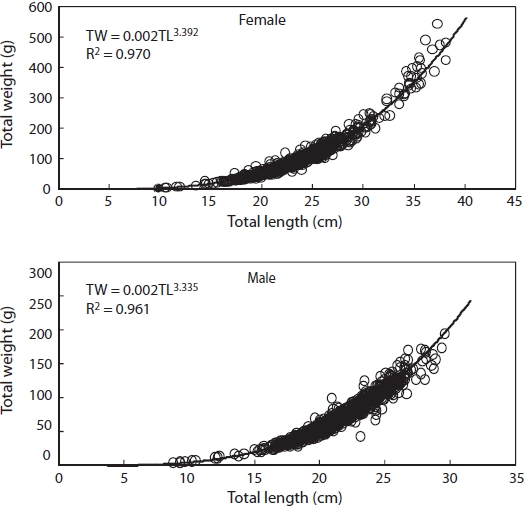
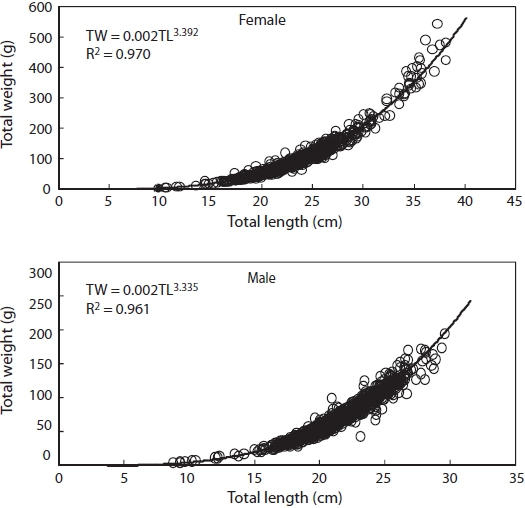
Relationship between TW and TL is shown in Fig. 6. It was TW = 0.002TL3.392 (r2 = 0.970) for females and TW = 0.002TL3.335 (r2 = 0.961) for males.
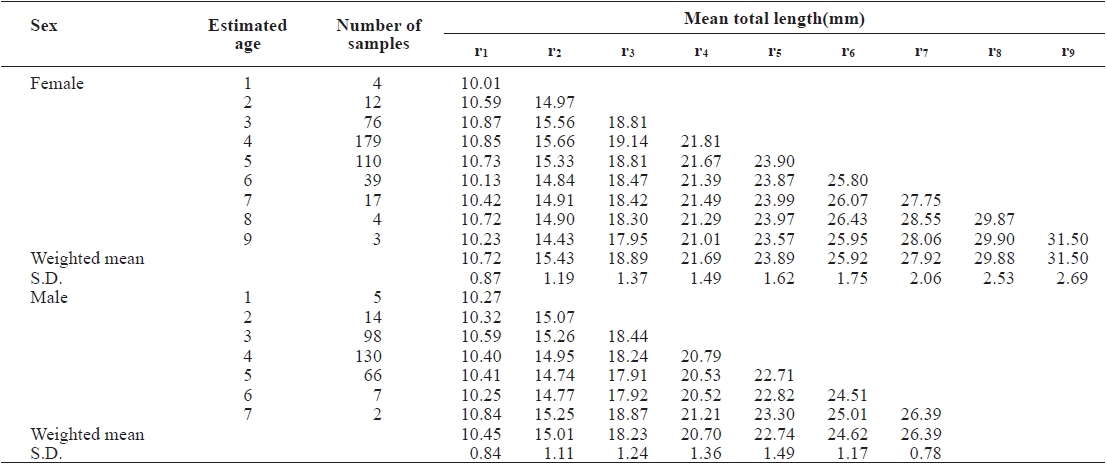
The average TLs for each annulus were back-calculated from the for going relationship between R and TL based on the average ring radii to the annuli (Table 2). The estimated TLs of
females were longer than those of males for all ages. The result of t-test used examine the growth difference for each age between males and females showed no significant difference from age 1 to 2 (P>0.05), but a significant difference from age 3 (P<0.05). It is assumed that the growth differential between male and female G. stelleri appears from the age of 3.
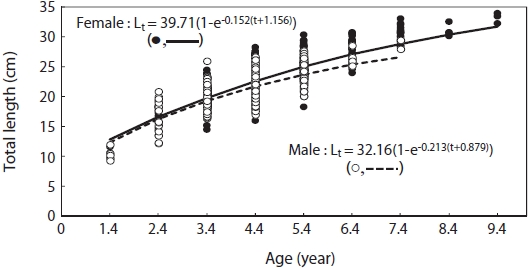
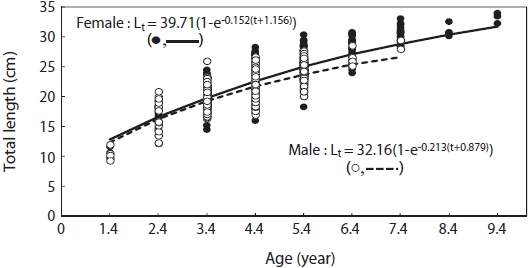
The Von Bertalanffy growth functions of male and female G. stelleri derived from nonlinear regression analysis , analysis are:
Lt = 39.71(1 - exp(-0.152(t+1.156))) for females; Lt = 32.16(1 - exp(-0.213(t+0.879))) for males (Fig. 7).
The age of G. stelleri can be determined from the otoliths by observation of the sagittal plane, where opaque and translucent zones clearly alternate. The ages of flounders have been determined through observation of the sagittal sections of otoliths in various species such as Pleuronectes herzensteini (Choi et al., 1986), Verasper variegatus (Jeon et al., 1996), P. yokohamae (Park, 1997), C. pinetorum herzensteini (Choe et al., 1999), Cynoglossus joyneri (Baeck and Huh, 2004a), and Cynoglossus abbreviatus (Baeck and Huh, 2004b).
The growth structures of the left/right otoliths and the annulus mark formation position are different in some flounders such as V. variegatus (Jeon et al., 1996) and P. herzensteini (Choi et al., 1986), but the annuli in the otoliths of flounders are formed uniformly in the eye-side otoliths. Therefore, the eye-side otoliths were used in this study.
Our results indicate that annulus of G. stelleri formed once a year in September, delineated by the boundary between opaque and translucent zones. Lee (2008) reported otoliths formed opaque zones from April. Hashimoto (1953) reported that vertebral annuli form translucent zones (annual mark)
during winter to spring. Ishida and Kitakata (1953) reported that otolith annuli form translucent zones from June to September, and that opaque zones are formed from March to May. Thus, the results of this study show differences in the formation of the annual mark.
The spawning period of G. stelleri has been reported to be April - June in the East Sea area of Korea (Cha et al, 2008), March - May in the Tottori, Japan (Tottori Prefecture Web Site, 2011), June - July in the Hokkaido, Japan (Hashimoto, 1953), April - July in Aomori, Japan (Hashimoto, 1953), May - September in Peter the Great Bay in Russia (Fadeev, 2005). Therefore, the spawning periods and formation of annual marks in G. stelleri show differences between sea areas.
We found that annulus formation occurred around September, with the first annulus formation at age 1.4 which is about 4 months after spawning period. Similar results have been reported for other flounders such as Eopsetta grigorjewi (Hwang et al., 1979), K. bicoloratus on the west sea coast (Jun and Im, 2004), and S. schlegeli inhabiting the Tongyeong marine ranch on the south coast of Korea (Park and Kang, 2007). Also, the annulus formation period of flounders in the East Sea was January - March for P. herzensteini (Choi et al., 1986), May for C. pinetorum herzensteini (Choe et al., 1999), and April - June for L. yokohamae (Kim et al., 1991), showing that the periods are different for each species, even when the environmental characteristics of their habitat are the same.
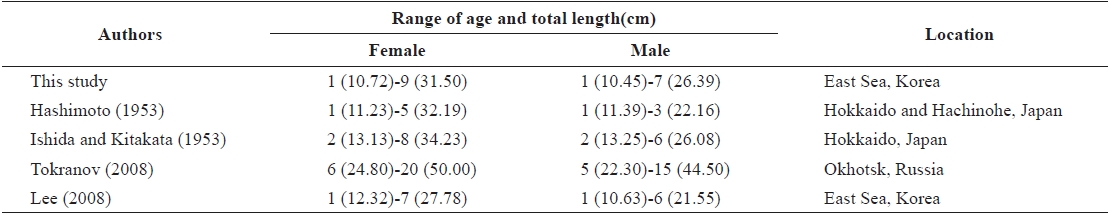
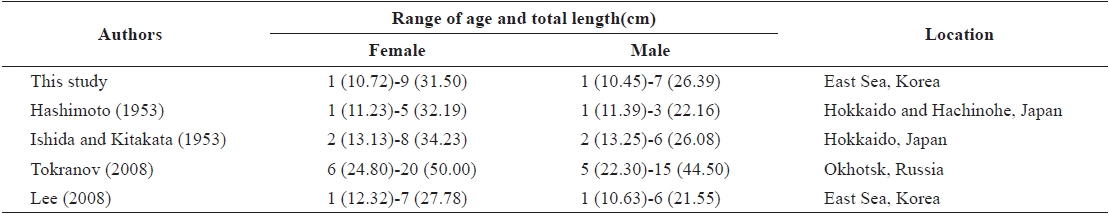
The growth of G. stelleri in Japanese and Russian sea is faster than that in the East Sea of Korea (Table 3). The maximum age of G. stelleri in Japanese sea is lower than that determined from the results of the present study, while the maximum age in Russian seas appears to be higher than that determined in the present study. Also, reviewing the actual size of samples, the maximum TL collected in this study was 38.1 cm, but the TL of G. stelleri in Russian seas was recently reported to be 62 cm by Orlov and Tokranov (2007), which is markedly longer than the fish examined in this study.
There was no significant difference between males and females in the annual growth rate of G. stelleri up to age 2 (P>0.05), but the growth rate of females was faster than that of males after age 3 (P<0.05). Hashimoto (1953), Ishida and Kitakata (1953), and Tokranov (2008) reported similar results, with the growth rates of females being faster than those of males at an older age. The oldest specimens in this study were a 9-years-old female and a 7-years-old male. The oldest individuals in other studies have been 8 and 6 in the G. stelleri samples from Hokkaido Ken, Japan (Ishida and Kitakata, 1953), 5 and 3 in samples from Hokkaido Sea and Aomori Prefecture, Japan (Hashimoto, 1953), and 20 and 15 in samples from Kamchatka in the Sea of Okhotsk (Tokranov, 2008) for females and males, respectively.
Geographic differences in the growth rates and longevities of G. stelleri are considered to be related not only to environmental factors, but also to fishing intensity. Catchs of G. stelleri in the East Sea have an increasing trend amounting to 54.9% of the total catches of the flounders (National Fisheries Research and Development Institute, 2010). High fishing intensity leads to the mean age of caught fish being reduced (Zhang, 1991). The low mean age of G. stelleri caught in the East Sea could indicate high fishing pressure. Intensive stock assessments and research on management plans are needed to rebuild the population of Korean flounders in the East Sea.