Macroalgae are important primary producers in marine ecosystem. They don’t only play an important role as bio-indicators but also provide economic resources for humans. Seasonal and vertical variations in seaweed species com-position and biomass were examined to determine the ecological status of seaweed beds around Udo, near Jeju Island. We obtained samples at two sites in the high-intertidal to subtidal zones using the quadrat method between June 2010 and May 2011. A total of 262 species were collected, including 31 green, 61 brown, and 170 red algae. The composition of algal species revealed a decrease in species growing in cold water in comparison with the list 20 years ago. The macroal-gal mean biomass (g wet wt m-2) was 3,476 g and 2,393 g from the two sites, respectively. Ecklonia cava had the greatest biomass at both sites. The seasonal dominant species by biomass at site 1 from the low-intertidal to 1-3 m depth of the subtidal zone was mostly comprised of thick-leathery form, such as Sargassum hemiphyllum, S. coreanum, and Ecklonia cava, whereas site 2 was comprised of the turf form, such as Chondrophycus intermedius, Chondracanthus intermedius, Dictyopteris prolifera, and Gelidium elegans. The current ecological status of the seaweed community in Udo is stable based on diversity and dominance indices.
Macroalgae, key primary producers in marine ecosys-tem, play important ecological roles such as providing spawning grounds and habitat for fish and shellfish. They also provide medicinal and industrial materials, includ-ing feed, fertilizer, and bioenergy resources (Graham et al. 2009). Algae-based biofuels are receiving attention as an alternative to petroleum and grain-based fuels (Aresta et al. 2005). The reasons for this interest are that 1) marine algae have a very high productivity level by fixing CO2 and 2) they are not a staple food. Additionally, annual produc-tivity of macroalgal community on rocky shores exceeds that of tropical rain forests (Dawes 1998). Investigations into macroalgal biomass have been undertaken to deter-mine whether macroalgae have a role in CO2 reduction along Korean coasts (Choi et al. 2008, Kim et al. 2008, Ko et al. 2008).
Furthermore, the pharmaceutical industry is inves-tigating the use of seaweed polysaccharides to develop better and safer drugs with fewer side effects than syn-thetic drugs (Athukorala et al. 2007). Marine algae have been explored as a resource for treating various medical conditions due to their antihypertensive, antioxidant, antibiotic, and antiinflammatory properties (Aoun et al. 2010, Tierney et al. 2010). Despite the vast potential ben-efits of seaweeds, collecting target algal species is com-plicated. The process requires a detailed understanding of algal life patterns, including habitat, seasonal or ver-tical variations, and life history. The macroalgal vertical distribution is mainly controlled by exposure to moisture in intertidal zones and light intensity in subtidal zones. Moreover, temperature is one of the most important fac-tors controlling latitudinal distribution (Dawes 1998). Expansion and retreat of marine species along coastlines during temperature changes have been documented. For example, seaweed distribution changes along the British Coast were documented during the first half of the twen-tieth century (Luning 1990, Hiscock et al. 2004, Muller et al. 2009).
Kang (1966) divided the Korean coast into five sections based on water temperature and other hydrological con-ditions. He designated Jeju Island as an independent sec-tion. Jeju Island is comprised of 2% boreal, 74% temper-ate, 10% subtropical, and 15% cosmopolitan macroalgal species. After his study, more than 340 macroalgal species were added to the list of algae, and about 860 species are currently present along the Korean coastline (Lee and Kang 2001, Lee 2008). However, little published informa-tion is available on long-term macroalgal variations asso-ciated with changes in seawater temperature in specific regions of the Korean coastline.
Udo is one of the most popular tourist spots in Korea. Many important commercial seaweeds occur along this coast, such as
The macroalgal community serves as a valuable bio-indicator, because macroalgal species respond rapidly to environmental changes in the coastal ecosystem (Tri-bollet and Vroom 2007). This study of the Udo area pro-vides baseline data needed for long-term monitoring of the macroalgal community that serves as a bioindicator for the region. The specific objectives of this study were (1) to document the macroalgal species composition to elucidate the relationship between algal composition and seawater temperature due to global climate change, (2) to identify the seasonal variations in biomass to esti-
mate the annual productivity of the macroalgal commu-nity, and (3) to determine the ecological characteristics that are useful for evaluating the ecological health of the macroalgal community prior to any increase in sewage volume.
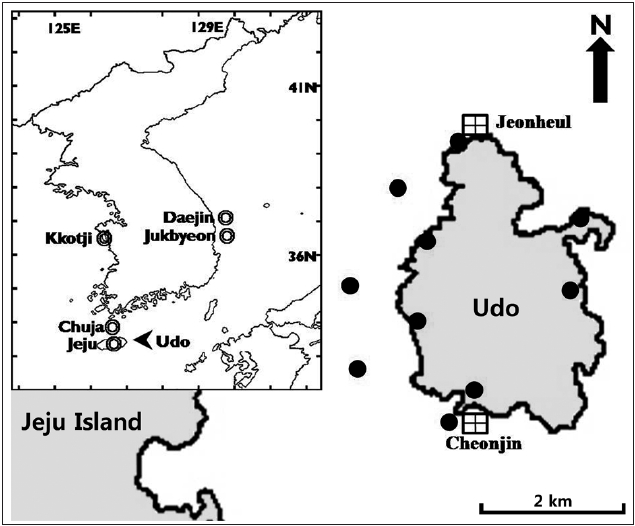
Udo is an elliptical islet extending north to south in the easternmost part of Jeju Island. We selected the southern (Cheonjin) and northern (Jeonheul) parts of Udo to in-vestigate macroalgal community structure. The Cheonjin site is situated on a concave coastline that is protected from strong winds and waves by both a seawall and Jeju Island itself. In contrast, the Jeonheul site protrudes into the sea and is exposed to strong wave action (Fig. 1). The distance between the two sites is about 3.7 km. At both sites, the substrate consists mostly of basaltic bedrock covered with sand in a depression of bedrock 7 m below the surface.
Macroalgal flora collections were conducted 17 times from June 2009 to May 2010 along the Udo coastline, in-cluding five subtidal sites and six intertidal sites (Fig. 1). For the analysis of species composition, 1-5 individuals of each species were collected and fixed in 5% formalin-seawater. The samples were identified using a micro-scope based on the descriptions of Lee (2008), and Yo-shida (1998). All identified species were divided into four regions (W, warm; T, temperate; C, cold; B, broad) and
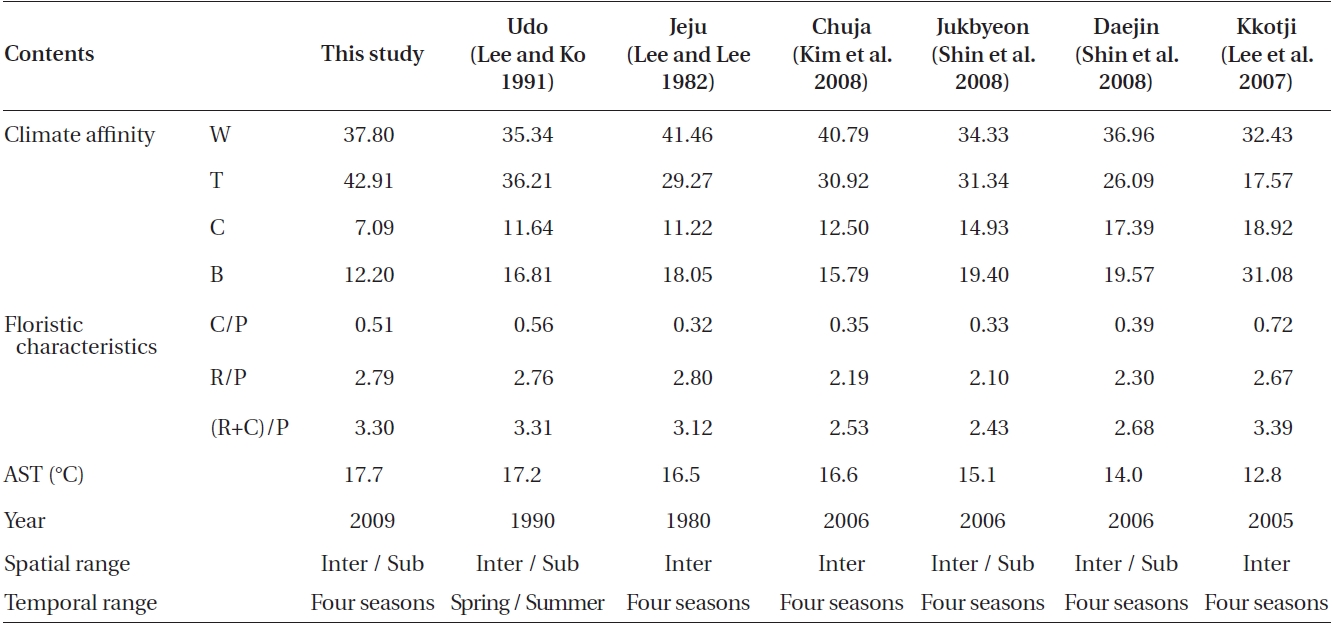
then the species composition was compared with previ-ously published studies of other coastal regions in Korea (Lee and Lee 1982, Lee and Ko 1991, Lee et al. 2007, Kim et al. 2008, Shin et al. 2008). Four regions were sectioned based on the geographical distribution of the macroal-gal species (http://www.algaebase.org, Guiry and Guiry 2011), and the seven groups of biogeographical regions in the world seas as defined by Luning (1990). We used the species composition data and annual mean sea sur-face temperature from the Korea Hydrographic and Oceanographic Administration for the correlation analy-ses (http://www.khoa.go.kr/). The C/P, R/P, and (R+C)/P ratios were also compared (Kim et al. 2008).
The macroalgal community structures were investi-gated seasonally for 1 year. Three 0.25 m2 quadrats were randomly placed on the high-, mid-, and low-intertidal zones at each site. In the subtidal zone, nine 0.25 m2 quad-rats were placed at interval depths of 1 m at each site. All marine algal species except melobesioidinean algae, were collected in the quadrats, and the biomass of each species was recorded (g wet wt m-2) in the laboratory. To exam-ine the variations in the macroalgal community structure along the shore gradients, we divided the shore into three intertidal (high, mid, low) and three subtidal (1-3 m, 4-6 m, and 7-9 m depths from the mean sea level) zones and placed three quadrats at each vertical shore zone. We per-formed cluster analysis using Pearson’s method (SPSS version 17.0; SPSS, Inc., Chicago, IL, USA) based on the macroalgal species lists. The diversity (
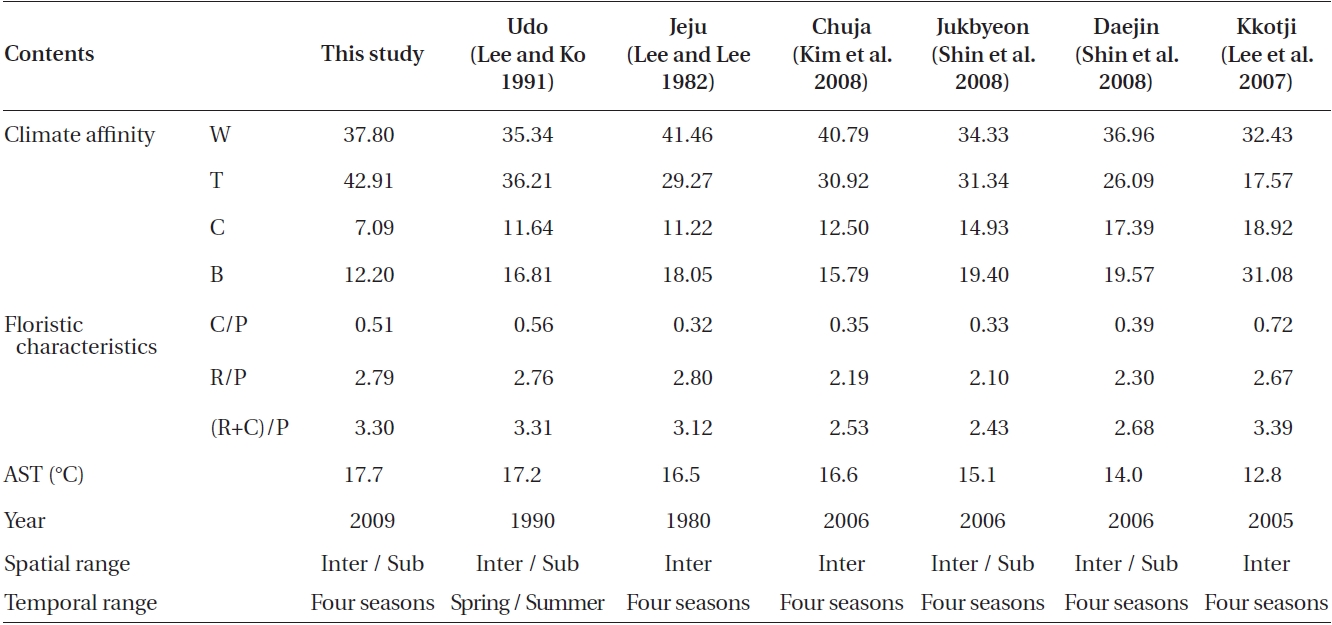
A total of 262 macroalgal species, including 31 Chlo-rophyta, 61 Phaeophyta, and 170 Rhodophyta were col-lected from Udo from June 2009 to May 2010. The C/P, R/P, and (R+C)/P ratios were 0.51, 2.79, and 3.30, respec-tively. The compositions of macroalgal species classified within the four climate affinities were 18 cold (7.09%), 119 temperate (42.91%), 96 warm (37.80%), and 31 broad (12.20%) species (Table 1).
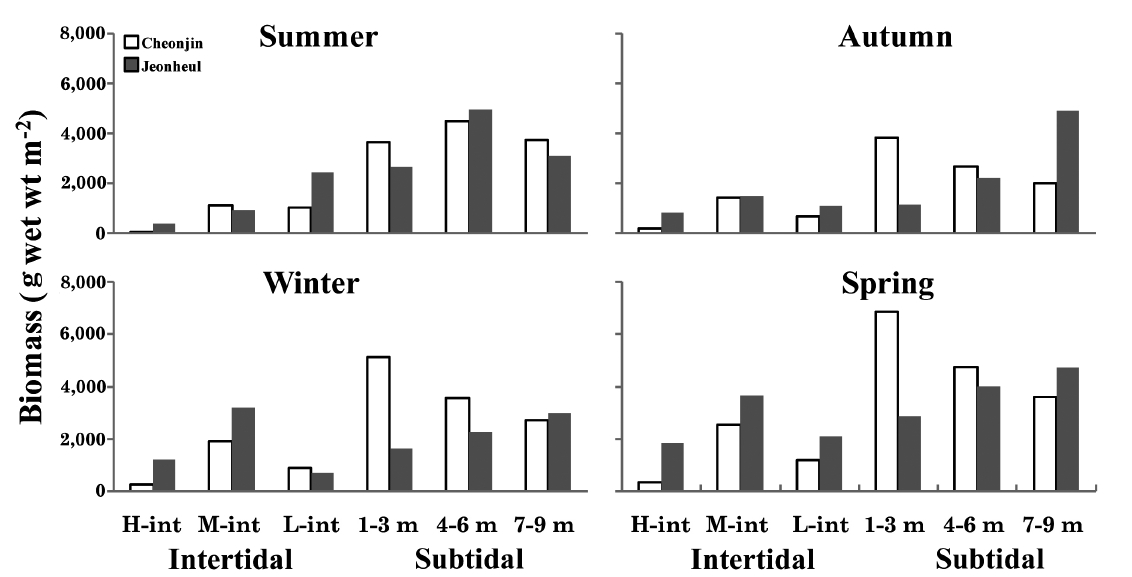
The annual mean biomasses (g wet wt m-2) of seaweeds were 3,476.4 from Cheonjin and 2,392.8 from Jeonheul.
Comparisons of species composition (%) based on the habitat climate zone and flora characteristics at several localities from the coasts of Korea
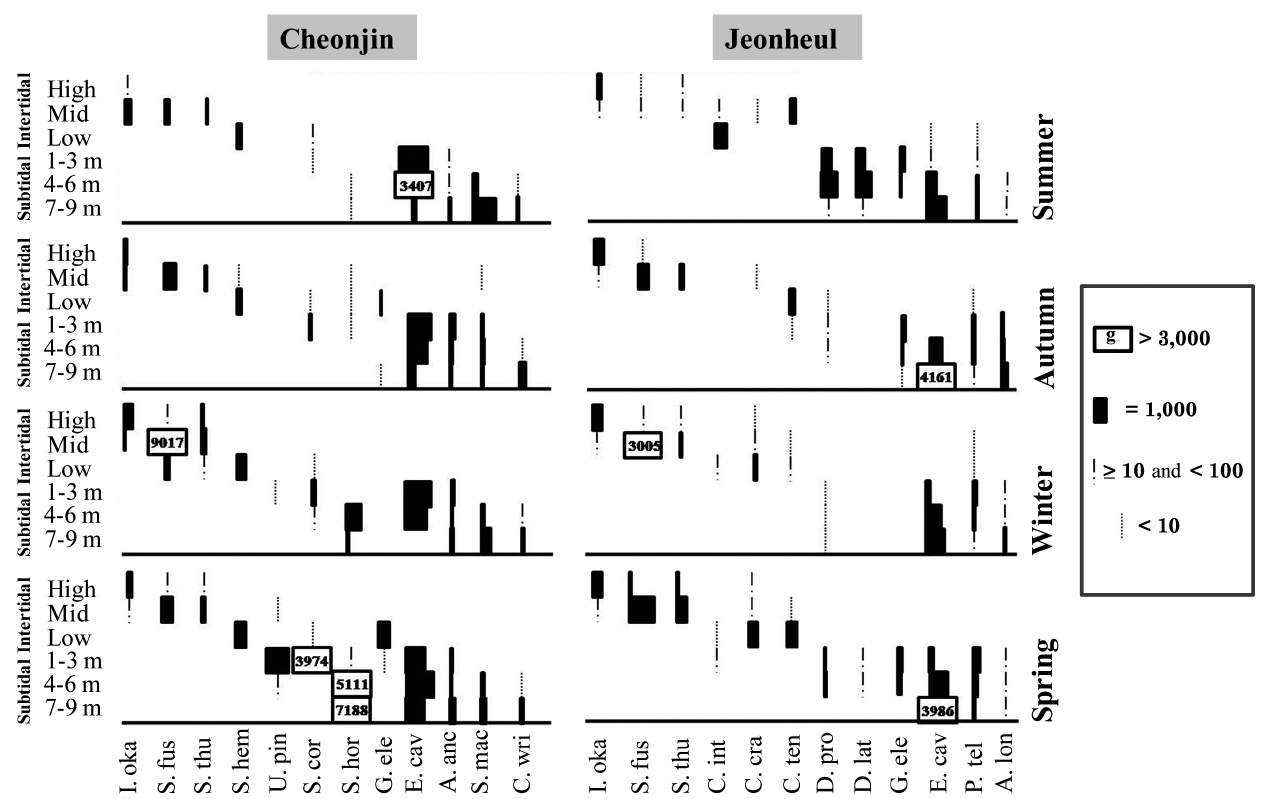
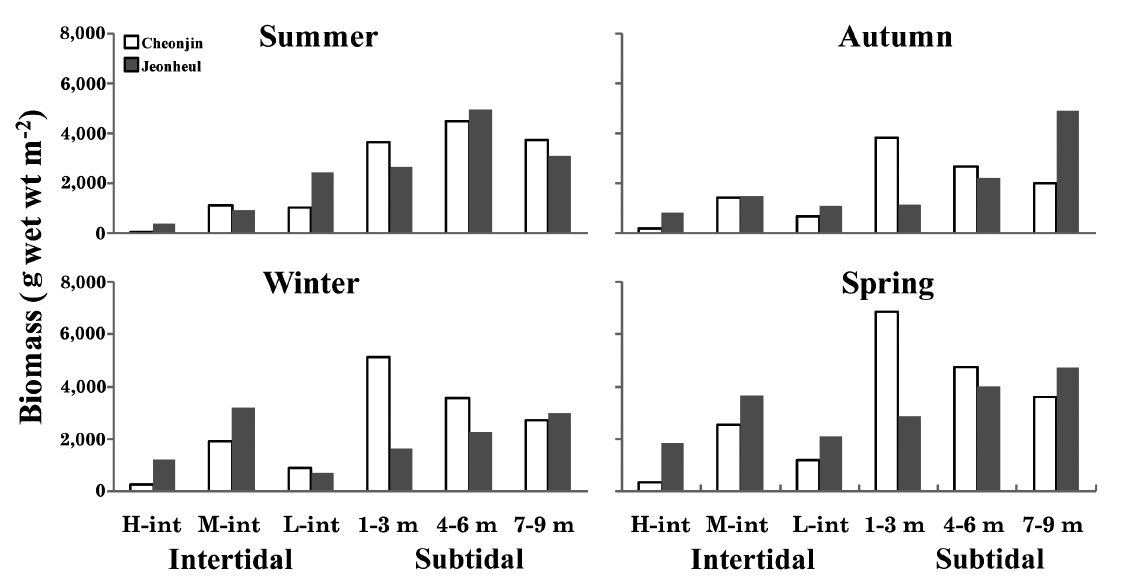
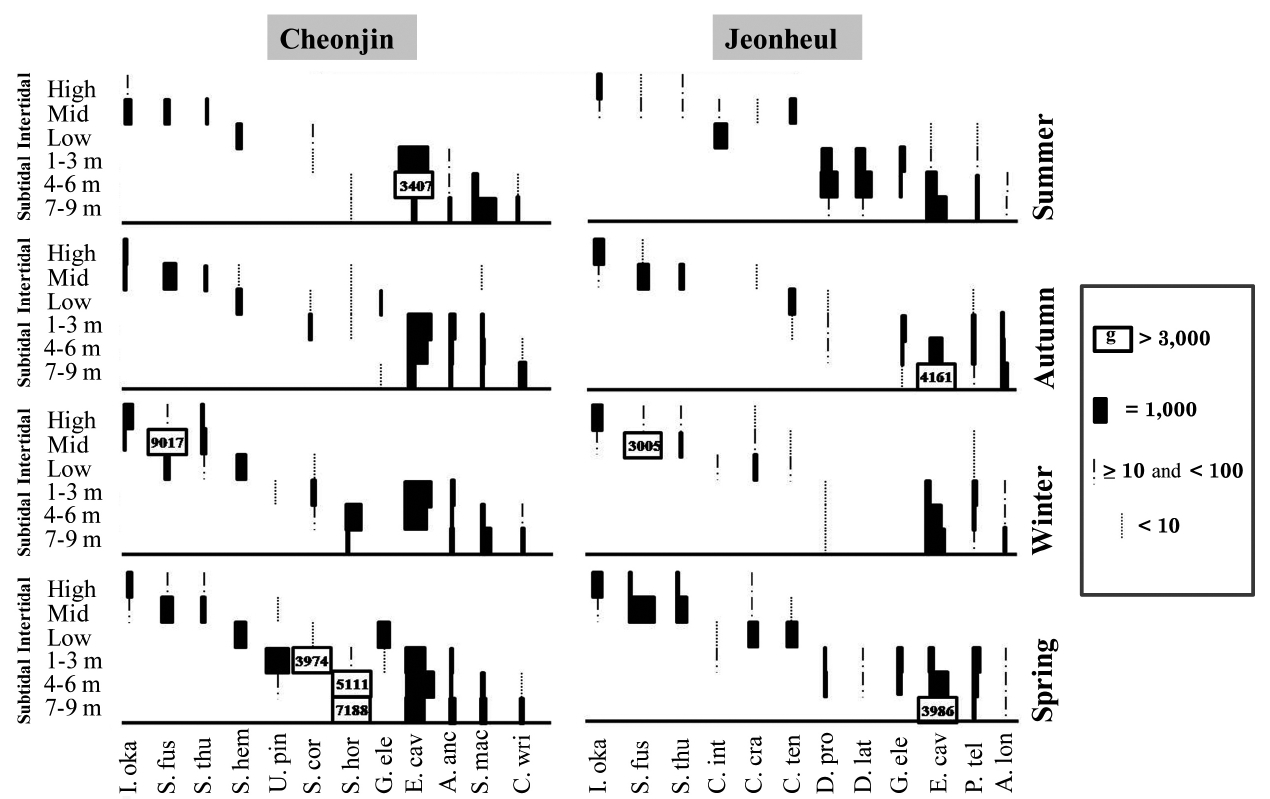
The seasonal and vertical variations of the dominant seaweed biomasses are shown in Fig. 3. The annual domi-nant species of each six vertical levels at Cheonjin were
Fig. 4 shows a cluster analysis dendrogram based on macroalgal species compositions. The dendrogram is di-vided into two groups corresponding to the intertidal (I) and subtidal (II) regions. Group I was divided into three subgroups and Group II into two subgroups (Fig. 4).
The five subgroups are characterized as follows: sub-group a) represents the high- and mid-intertidal zones at both sites, b) contains most of the low-intertidal zone at Jeounheul, c) includes all of the low-intertidal zone at Cheonjin, d) includes generally the 1-6 m depth level of the subtidal zone at Jeonheul, and e) contains most of the subtidal zone at Cheonjin and the 7-9 m depth level of the subtidal zone at Jeonheul.
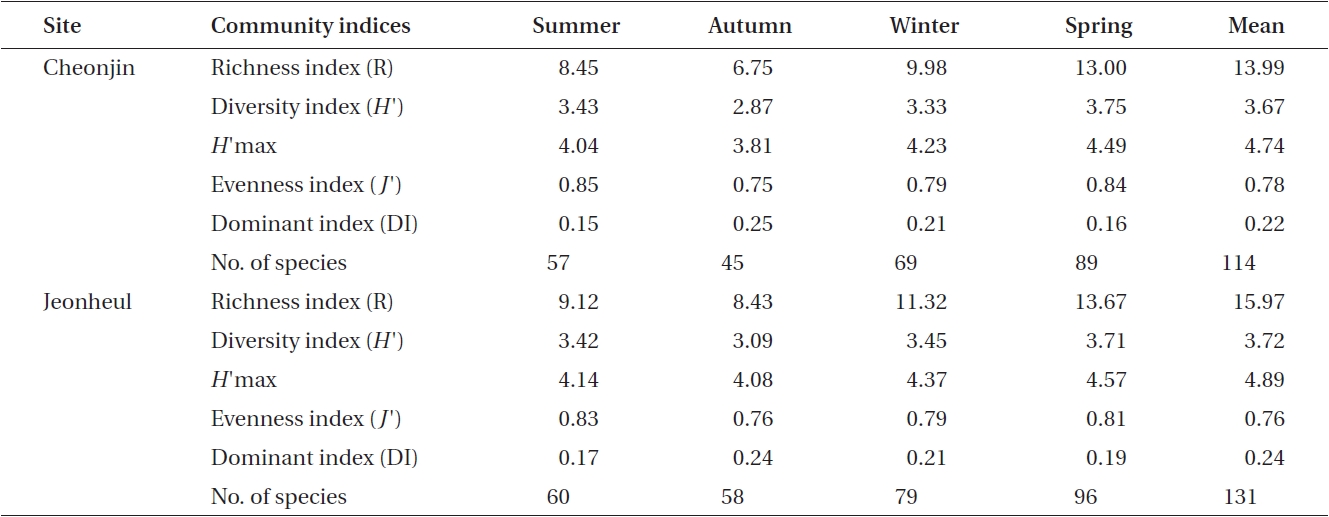
Ecological community indices from the two sites are shown in Table 2. The DI ranged from 0.15 to 0.25 at Cheonjin, and from 0.17 to 0.24 at Jeonheul. The highest DI was measured at both sites in autumn when the bio-mass of the two major species (
Seawater temperature is thought to be one of the im-portant physical factors determining seaweed distribu-tion (Druehl 1981). Macroalgal floras have been classified by the ratio of Chlorophyta and Rhodophyta species rela-tive to Phaeophyta species (C/P, R/P, [C+R]/P), and the ra-tios increase as the tropical zone is approached (Cheney 1977). The macroalgal species composition at Jeju Island compared to the east coast of Korea is characterized by a higher proportion of Rhodophyta. This observation is believed to be a consequence of the Tsushima Current throughout the winter and spring, and the Yellow Sea Warm Water Current during summer and autumn (Kang 1966, Lee and Kim 1999). Korean seaweed flora belong to the warm temperate (Van den Hoek 1984, Luning 1990)
[Table 2.] Seasonal variations in ecological macroalgal community indices from two sites at Udo
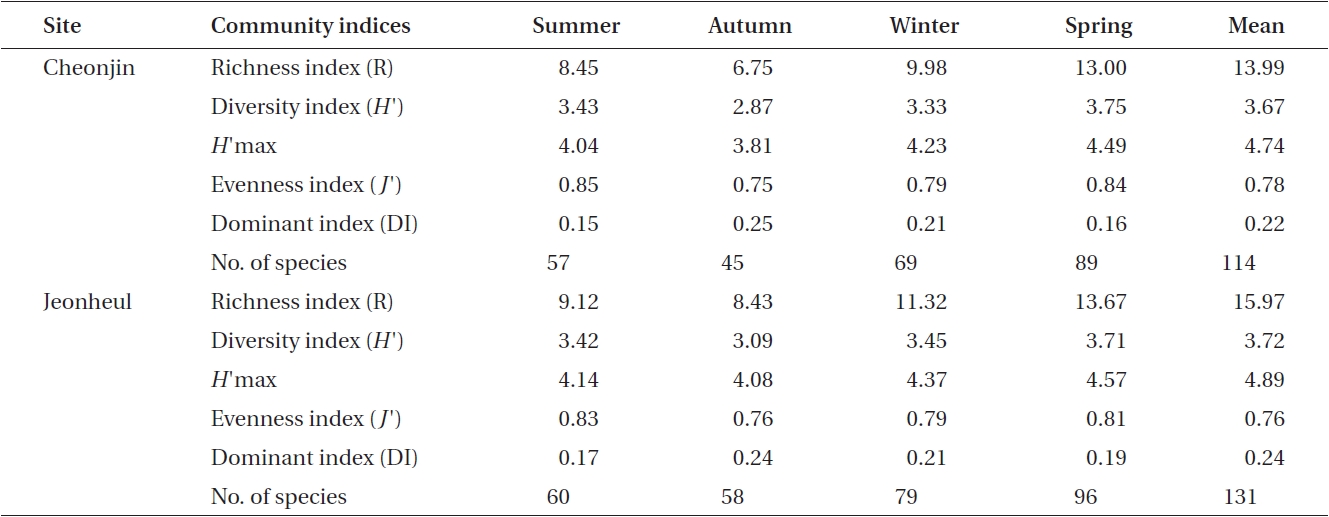
Seasonal variations in ecological macroalgal community indices from two sites at Udo
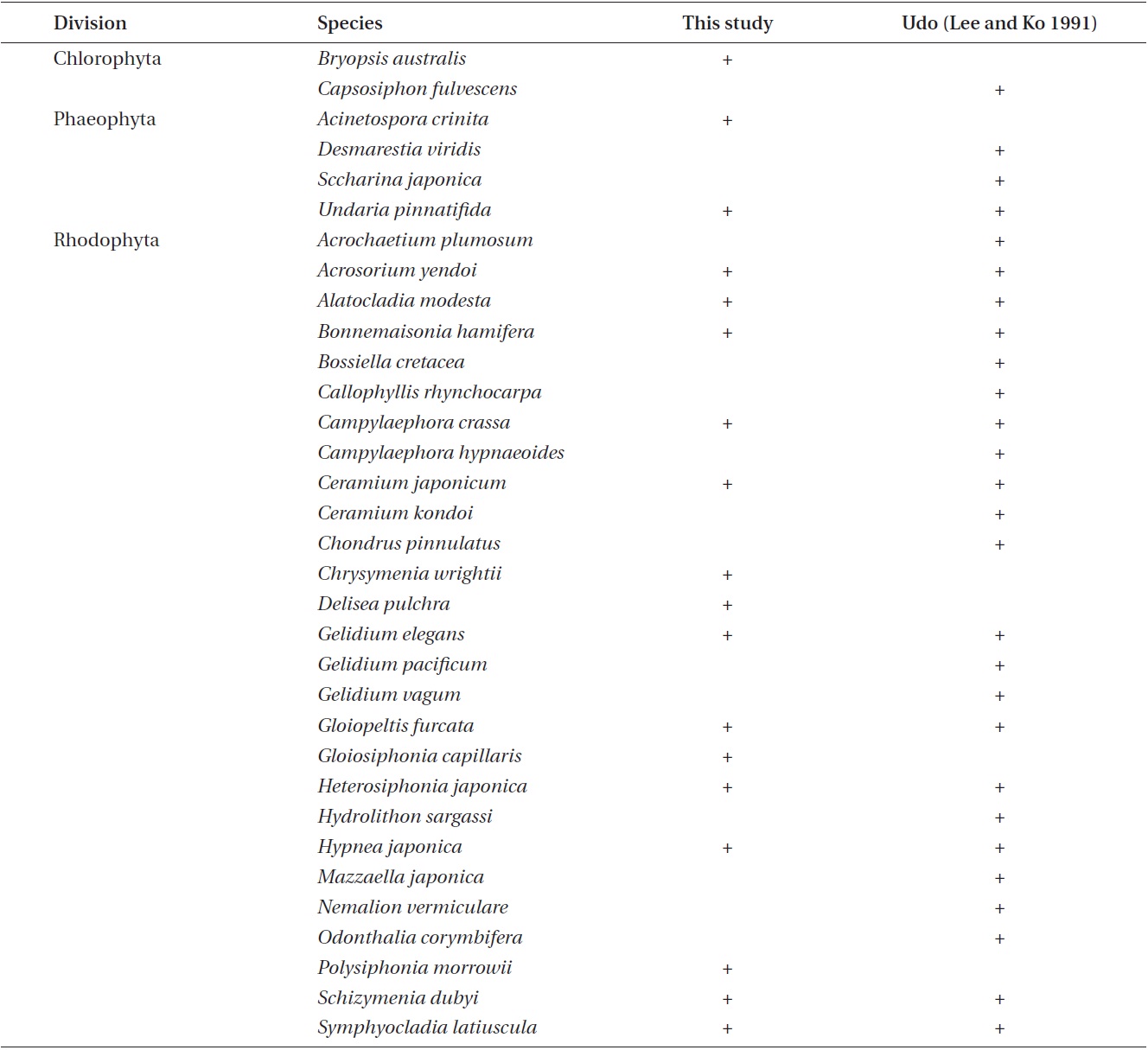
or cold temperate (Michanek 1979) region. It is clear from Table 1 that our results contrast with these earlier reports. The C/P ratio of our study was lower than that reported 19 years ago from Udo at Jeju (Lee and Ko 1991) and more recently on the western coast of Korea by Kkotji (Lee et al. 2007). The C/P ratio is somewhat higher than that documented 28 years ago at Jeju (Lee and Lee 1982), Chuja on the southern coast of Korea (Kim et al. 2008), and Jukbyeon and Daejin on the eastern coast of Korea (Shin et al. 2008). The R/P and (R+C)/P ratios are similar to the pattern of the C/P ratio except for the R/P ratios of Jeju (Lee and Lee 1982) and Kkotji (Lee et al. 2007). In our study, Udo had a greater proportion of Chlorophyta and Rhodophyta species than those on other Korean coasts. The C/P, R/P, and (R+C)/P values were directly propor-tional with latitude, although exceptions were observed, such as the C/P and (R+C)/P ratios for Kkotji (Lee et al. 2007). These exceptions could be related to local envi-ronmental factors or some unexplained spatio-temporal investigation variance such as the intertidal / subtidal, depth, period, or periodicity. Notably, the C/P, R/P, and (R+C)/P values obtained at Udo in this study were very similar to those reported about 20 years ago (Lee and Ko 1991).
In this study, the macroalgal species compositions from Udo were classified into four climate affinities: 7.09% cold, 42.91% temperate, 37.80% warm, and 12.20% broadwater species (Table 1).The cold and broadwater species comprised the lowest percentages in comparison with the ratio of other studies from Korea (Lee and Lee 1982, Lee and Ko 1991, Lee et al. 2007, Kim et al. 2008, Shin et al. 2008). The temperate-water species ratio trended in a manner different than those of broad and coldwater species. We could not discern any pattern in the warm-water species. The ratio of coldwater species showed a tendency to increase in relation to the annual sea surface temperature and latitude. In particular, the coldwater species in this study were considerably less abundant than those in previous studies of 19 years ago at Udo (Lee and Ko 1991) and 28 years ago at Jeju (Lee and Lee 1982). Lee and Ko (1991) reported some coldwater species from Udo, such as
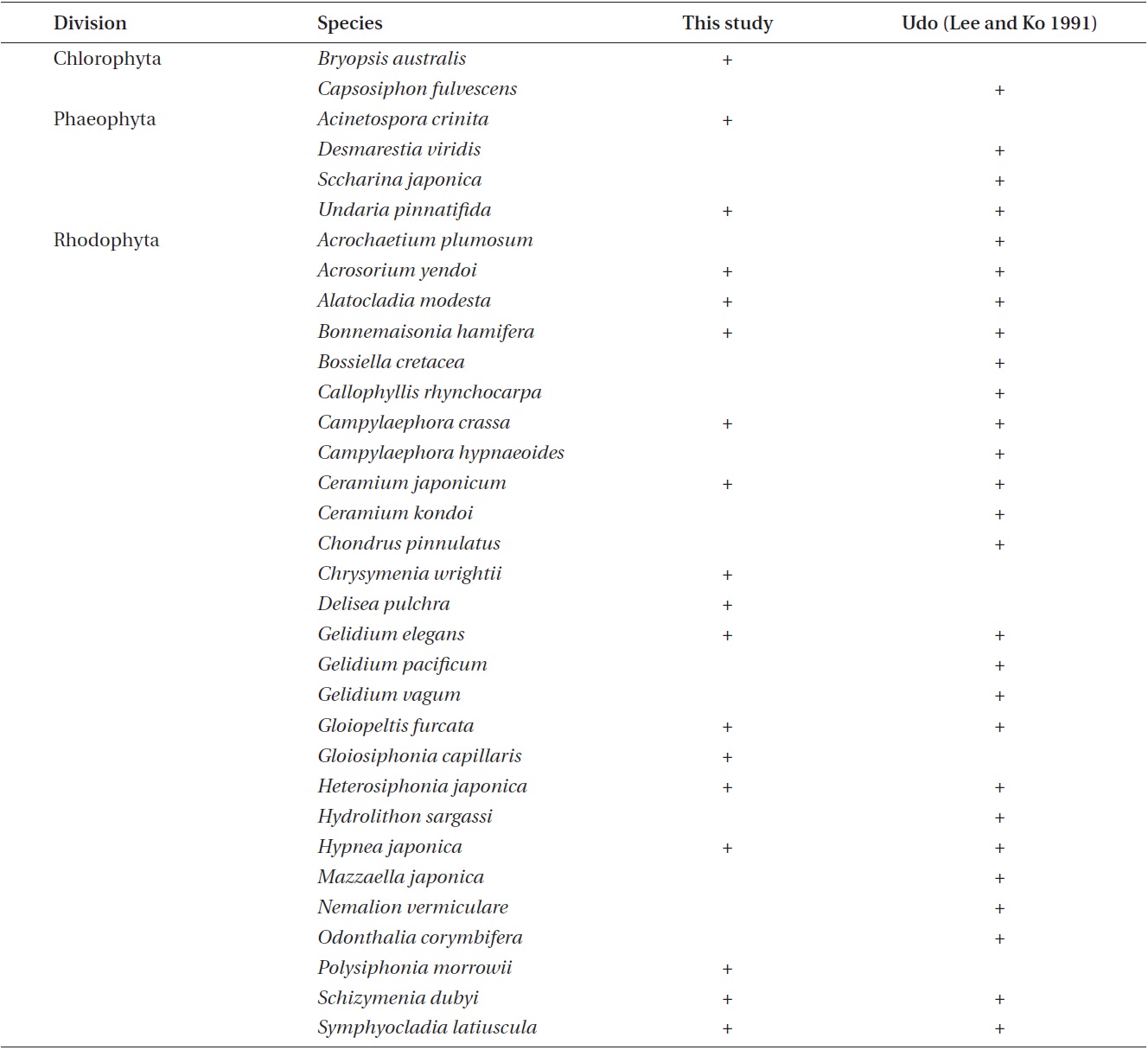
Comparisons of the appearance of coldwater species between this study and previous studies at Udo
paratively large in size and have obvious characteristics useful for identification, we did not find or collect any of these species in this study (Table 3). Instead, we were able to find and collect
Macroalgal and periphyton ecologists are interested in defining the factors that influence species occurrence and distribution patterns (Graham et al. 2009). Coastal marine communities have proven valuable as ecological models for determining the relative importance of physi-cal factors, algal morphological and physiological adap-tations, and biotic factors that structure macroalgal com-munities (Taylor et al. 1990, Orfanidis et al. 2001, Graham et al. 2009). Tide and light attenuation are the most im-portant determinants of intertidal and subtidal zonation patterns of macroalgal assemblages. As the intertidal re-gion is submersed at high tide and exposed at low tide, the upper intertidal region is exposed significantly for longer periods than that of lower areas (Doty 1946, Gra-ham et al. 2009). Clear vertical zonation patterns were ob-served in the macroalgal communities at both Udo sites (Fig. 3 & 4). The macroalgal communities at the high and mid levels of the intertidal zone were composed of
Water movement is one of the most important vari-ables influencing marine macroalgae, because it regu-lates nutrient availability, turbidity, and light penetra-tion (Nishihara and Terada 2010). In many studies of the wave-exposure effect on macroalgal communities, an intermediate level of water motion improves the diversity and abundance of seaweeds by accelerating nutrient and gas exchange, whereas a high level of water motion either decreases diversity or promotes the abundance of cer-tain species such as short clump turfs and kelps (Prathep 2005, Thongroy et al. 2007, Nishihara and Terada 2010). In the present study, the macroalgal community at the shel-
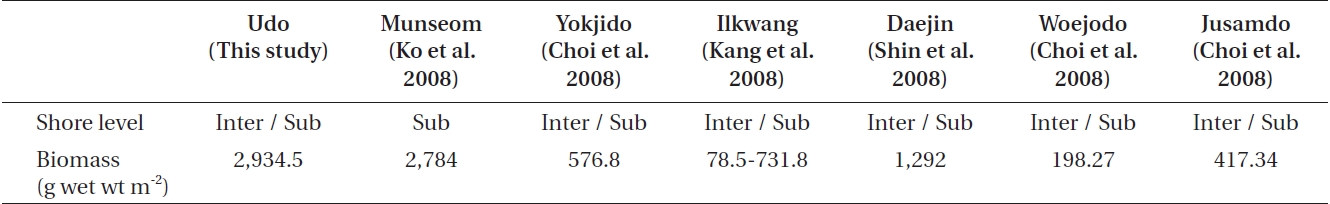
Comparisons of mean biomass among this study and previous studies from other Korean Peninsula coasts
tered Cheonjin site showed high biomass and more stable characteristics, whereas that in the exposed Jeonheul site contained various dominant species influenced by sea-son (Fig. 3). Kelps (
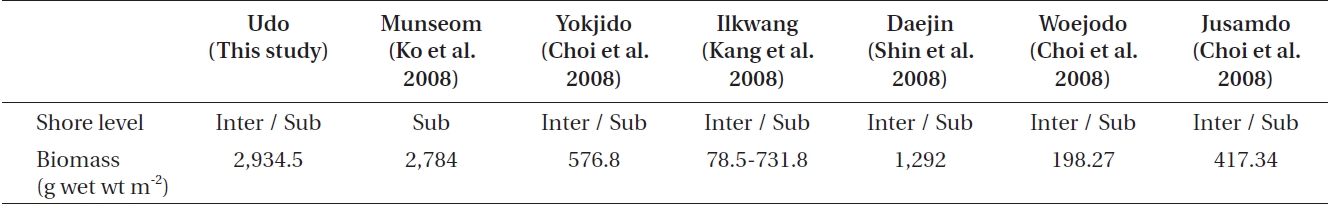
A comparison of the mean biomass between this study and recently published studies (Table 4) indicates that the mean biomass at Udo was similar with that of the subtidal zone of Munseom, Jeju Island (Ko et al. 2008). However, it was higher than that of other coastal regions of Korea, such as Yokjido and Ilkwang Bay, Gyeongnam (Choi et al. 2008, Kang et al. 2008), Daejin, Gangwon (Shin et al. 2008), and Woejodo and Jusamdo, Jeonbuk (Choi et al. 2008). The ecological community indices indicate that Udo has stable macroalgal communities (Lee et al. 2007).
In conclusion, Udo on Jeju Island maintains a higher macroalgal diversity level and biomass than other coastal regions of Korea. However, due to the construction of new water supply facilities from Jeju Island to Udo in 2010, it is possible that the overflow of fresh water or other pollu-tion from these facilities could damage the uncontami-nated sea areas of Udo. In contrast to a previous report (Lee and Ko 1991) on macroalgal species composition, we found that the number of species growing in cold water at Udo had decreased, whereas the number of high-temper-ature resistance species had increased. We speculate that this result is related to the increase in annual sea surface temperatures caused by the greenhouse effect. Addition-al studies involving continuous long-term monitoring of water quality, sea temperature, and species composition of seaweeds are needed. The differences in the compo-sitions of the macroalgal communities at the two sites suggest that the huge artificial construction along the shoreline could affect seaweed species composition due to changes in physical factors. The results of the present study indicate that the ecological status of the seaweed community at Udo is stable for now, and provide a base-line database to begin long-term monitoring studies to protect the abundant macroalgal beds of Udo.

![Map showing the collecting sites (qualitative analysis [dark circles] quantitative analysis [squares]) of macroalgal samples at Udo and the comparison sites (double circles) from references.](http://oak.go.kr/repository/journal/11258/JORHBK_2011_v26n4_333_f001.jpg)