Here we report the first finding of Cryptonemia rotunda and Schizymenia apoda in Korea based on plastid rbcL se-quence analysis and morphological observations. C. rotunda occurred in the subtidal in Hongdo and Namyeodo on the south coast. Thalli have short stipe with foliose, membranous blades with dentate margin, a length of 2-8 cm. S. apoda occurred in many locations along the coast of Korea. Thalli are foliose, simple or broadly lanceolate blades with irregular lobes and proliferations from the margins, sometimes undulate, soft fleshy, and slippery, up to 32 cm tall, and have cys-tocarps with ostioles on the blade. Plastid rbcL sequences reveal that each of these species is clearly separated from other species of their respective genera. S. apoda was well resolved within the genus. However, C. rotunda was distantly related to other members of the genus, and this result needs an urgent revision of the genus.
During floristic study of Korean marine algae, we found two unrecorded species;
>
Taxon sampling and morphology
Field collections of
[Table 1.] Materials used in the present study
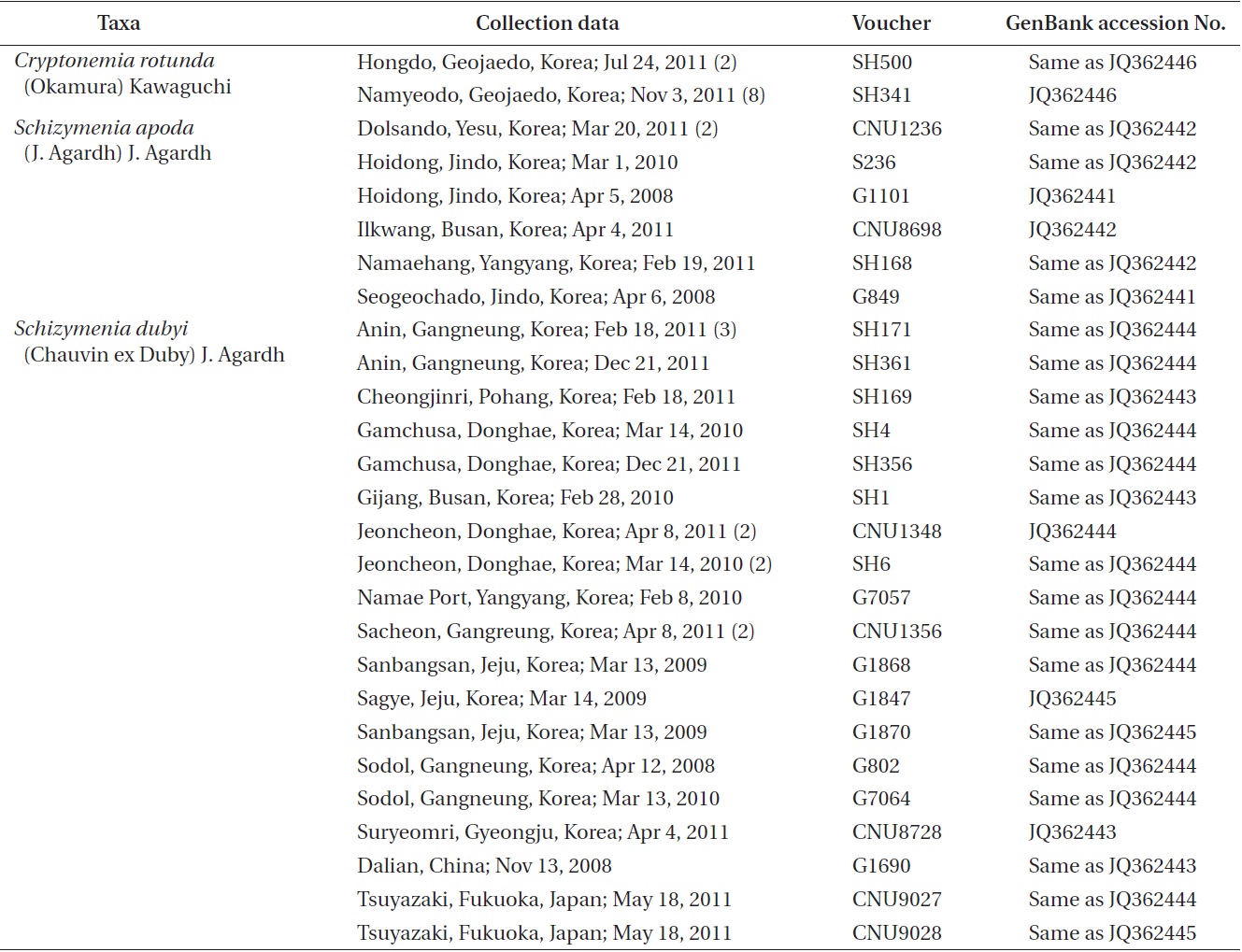
Materials used in the present study
using SCUBA diving. Tissues were sectioned with a freez-ing microtome (FX-80; Yamato Kohki Industrial Co. Ltd., Saitama, Japan). Photographs were taken with a DP-71 camera (Olympus, Tokyo, Japan) attached to a micro-scope (BX 51; Olympus). Voucher specimens are housed at the herbarium of Chungnam National University, Dae-jeon, Korea (CNUK).
Forty-one specimens were available for extraction of DNA: 10 for
Maximum likelihood (ML) phylogenetic analysis of
>
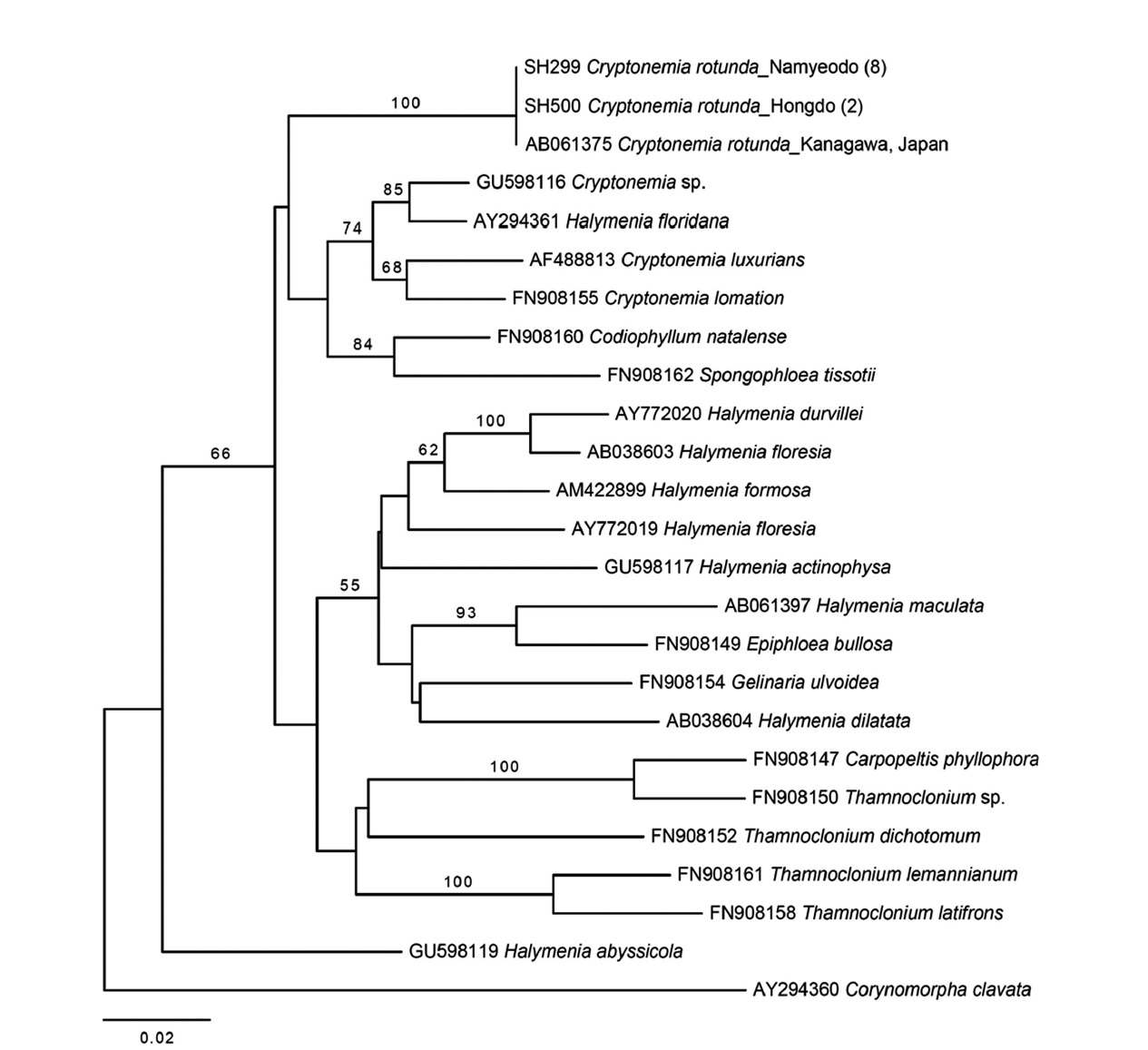
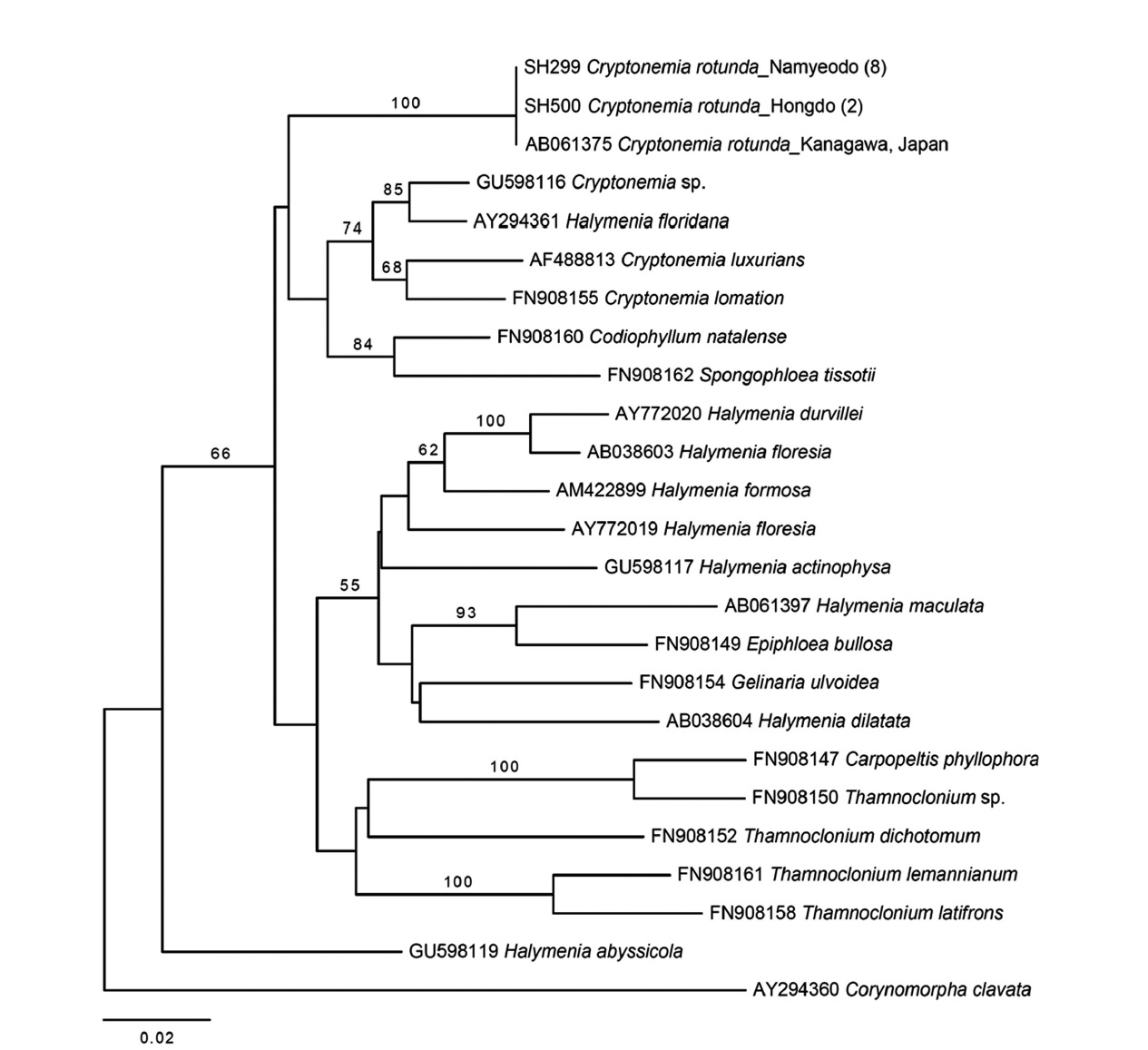
Cryptonemia rotunda (Okamura) Kawaguchi
Molecular analyses. Thirty-three
Plastid
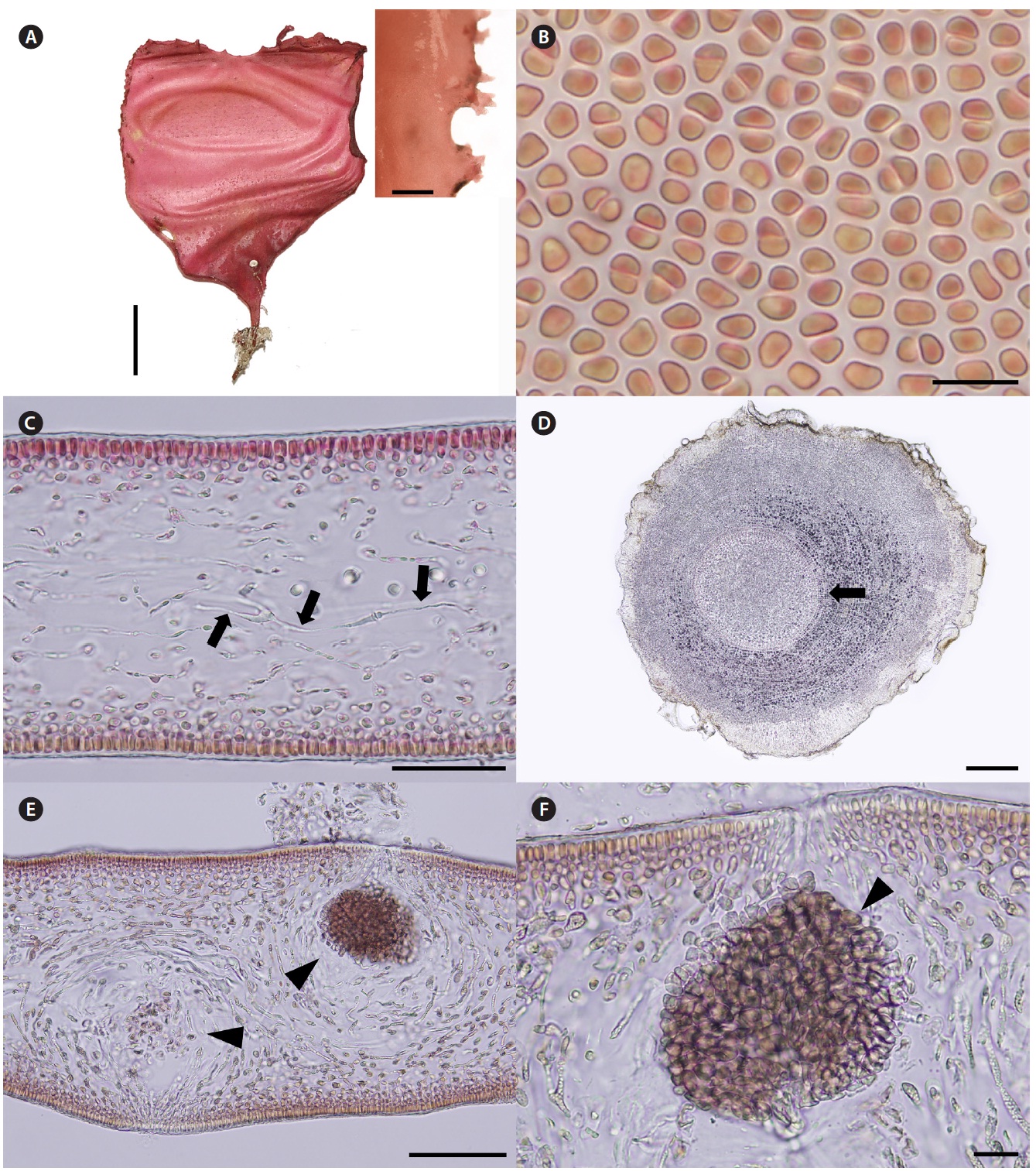
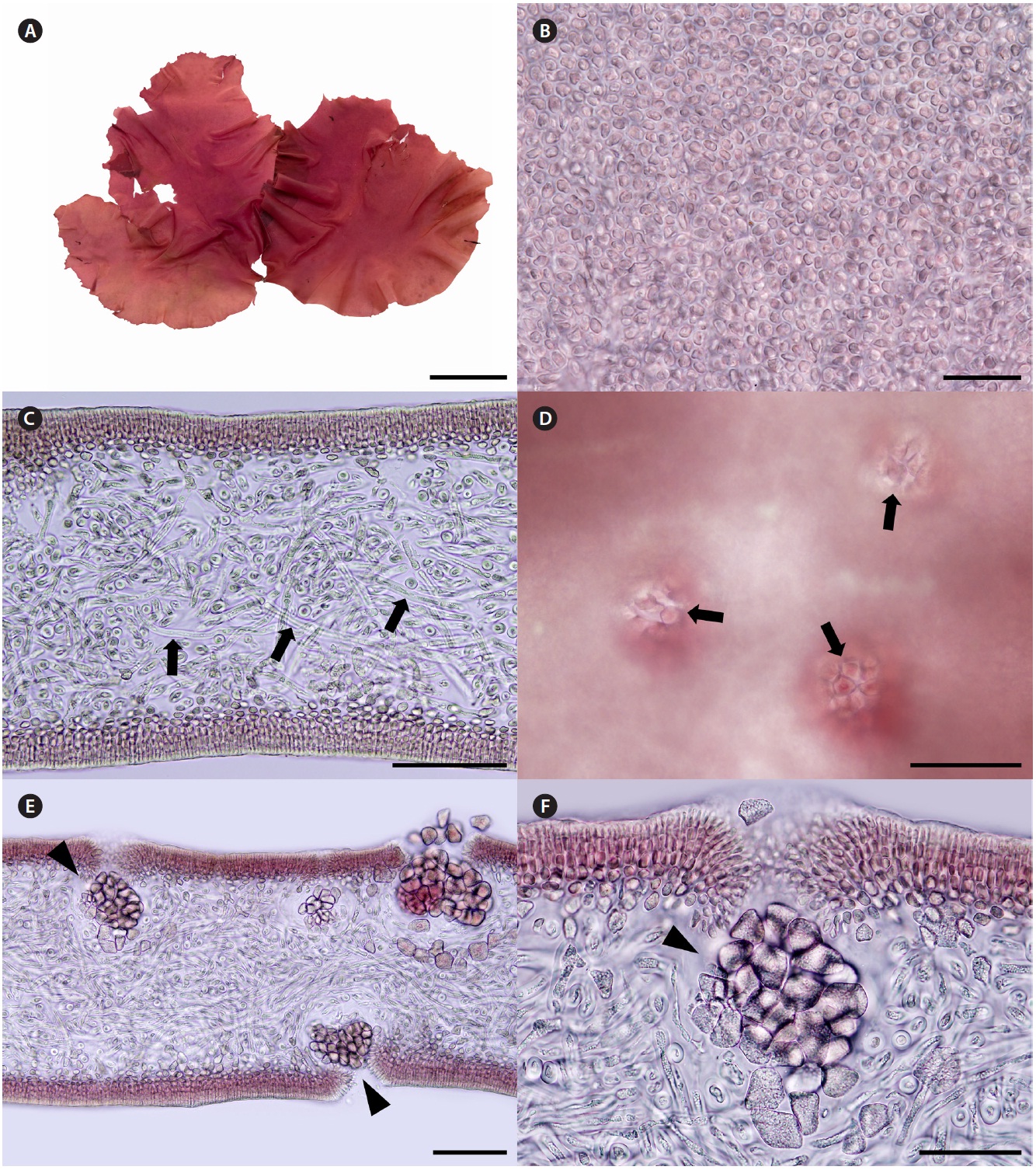
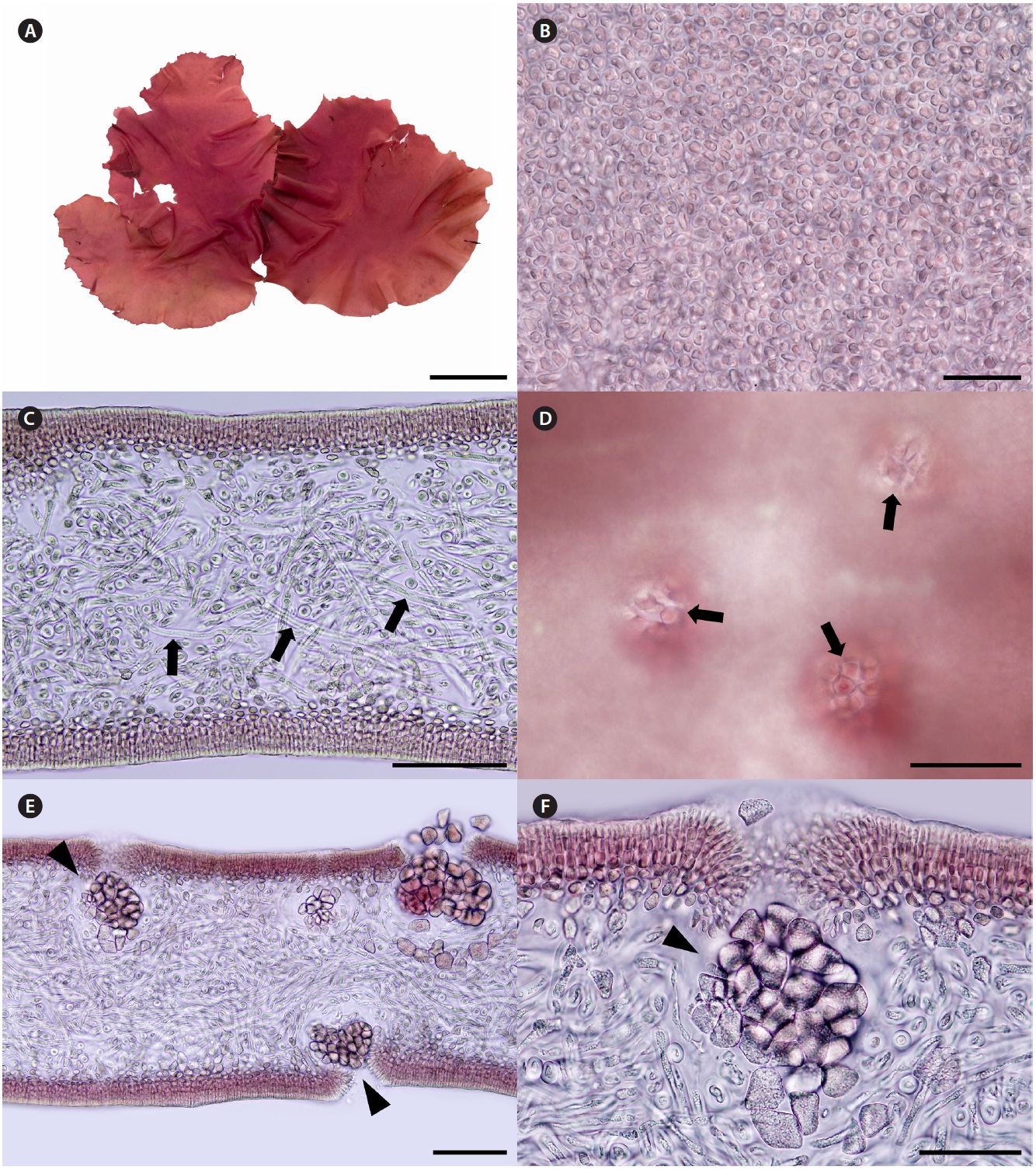
Korean name. 둥근바다선인장 (신칭) (Fig. 2A-F)
Description. Thalli are medium to dark red, membra-nous, circular or ellipsoid, rarely irregularly shaped, up to 8 cm in length and 4 cm in width, expanding from a stipe (Fig. 2A). Blades are thickened along the margin and beset with numerous, small protuberances, looking toothed (Fig. 2A), up to 200 μm thick (margins 300-370 μm thick). Cells at the surface of blade are angular (Fig. 2B). Tissue consists of cortex and medulla (Fig. 2C). Corti-ces are compact with 2-5 layers of cortical cells and 20-30 μm thick. Medulla are 120-180 μm thick, of entwined re-fractive filaments 1-4 μm in diameter (Fig. 2C). Holdfasts are discoid, 1-3 mm across, bearing stiff stipe which have several growth rings in cross section (Fig. 2D).
Female reproductive structures are scattered around upper side of the blade. Cystocarps (Fig. 2E & F) are sub-globose, 80-100 μm across, and swollen on the surface of blades with small ostiole. Both male plants and tetraspo-rangial plants were not found.
Thalli grow on rocks at a depth of 14-18 m in the sub-tidal zones. Thalli are larger in July than in November.
>
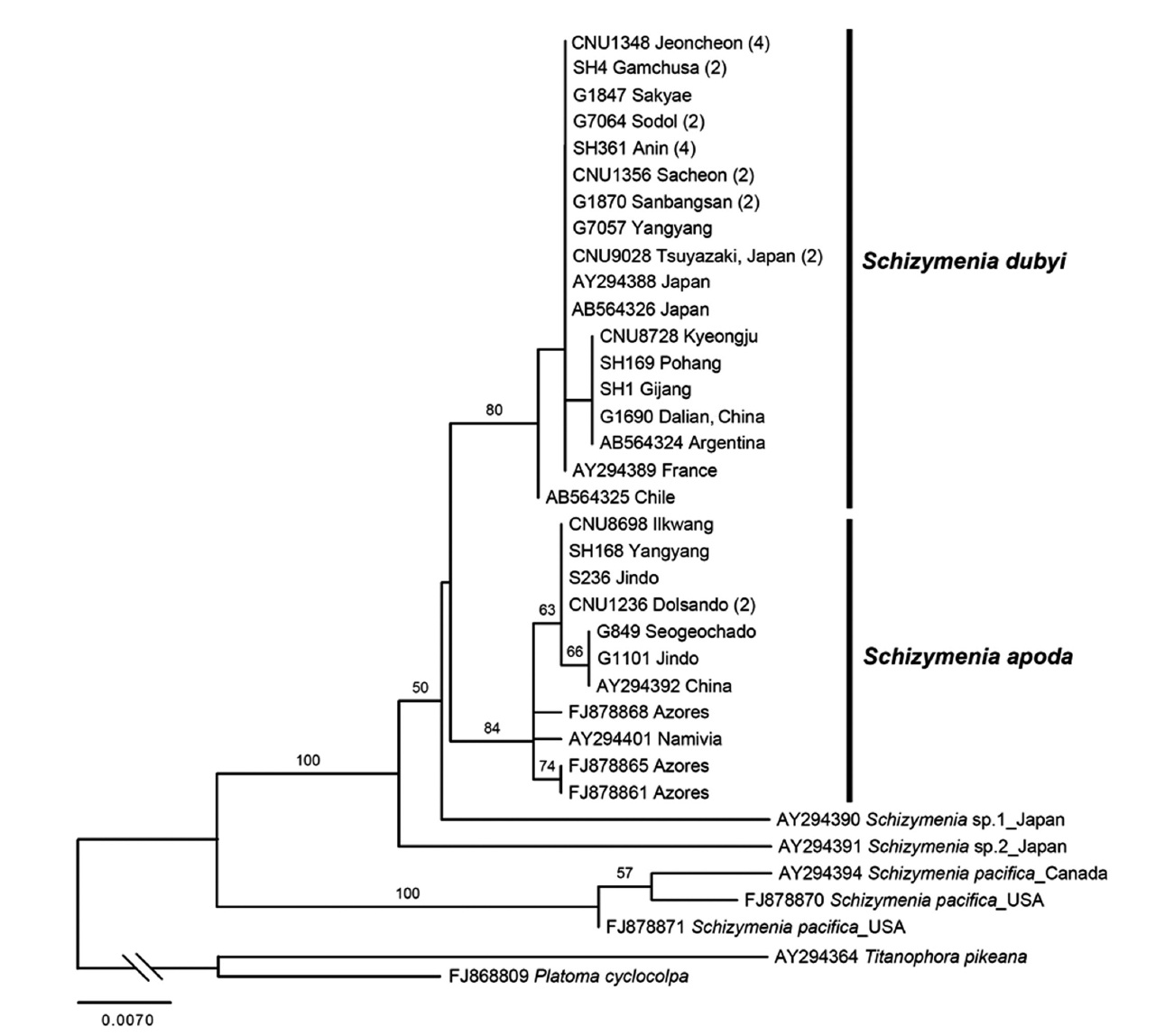
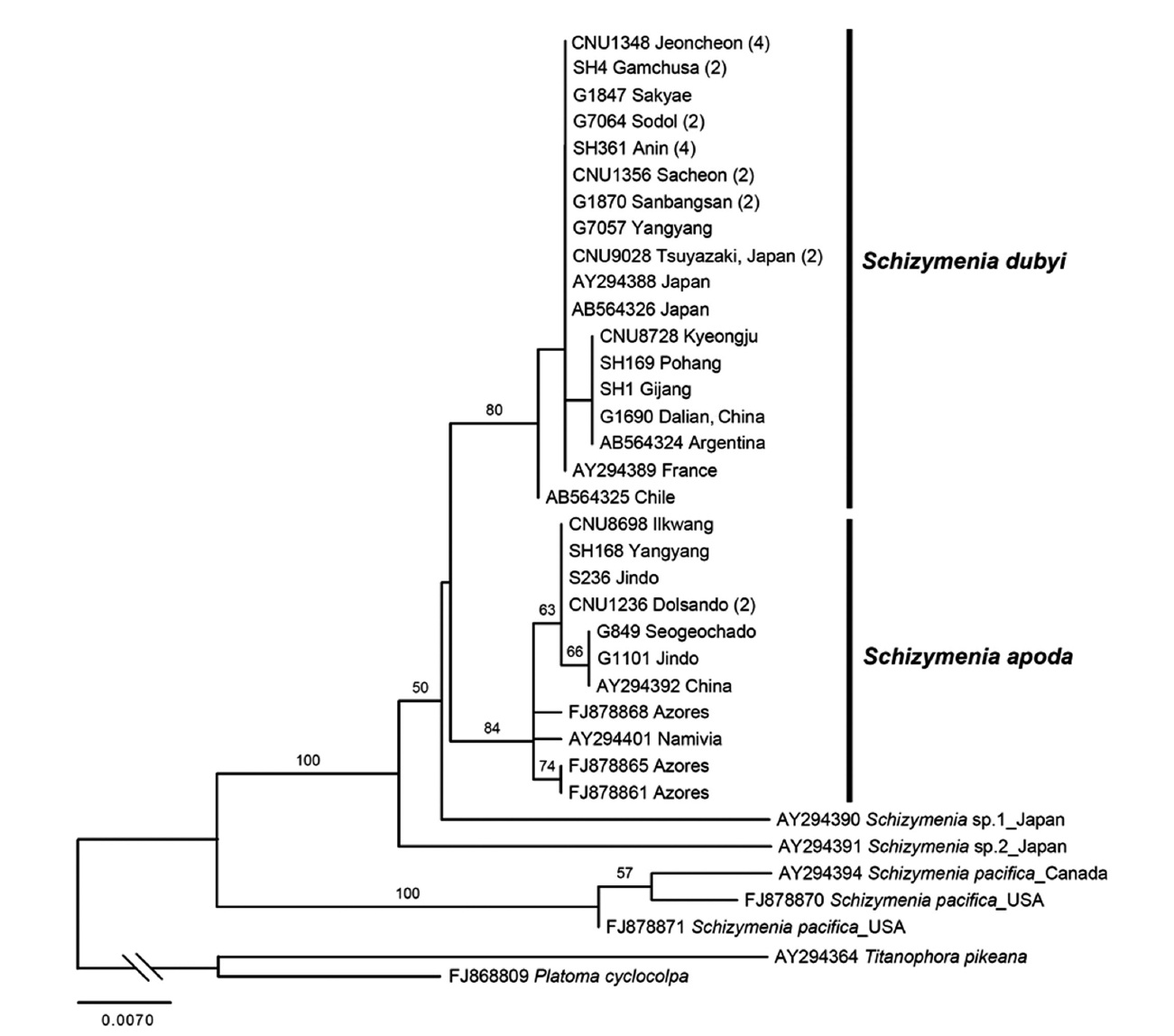
Schizymenia apoda (J. Agardh) J. Agardh
Molecular analyses. A total of 48
France and Chile.
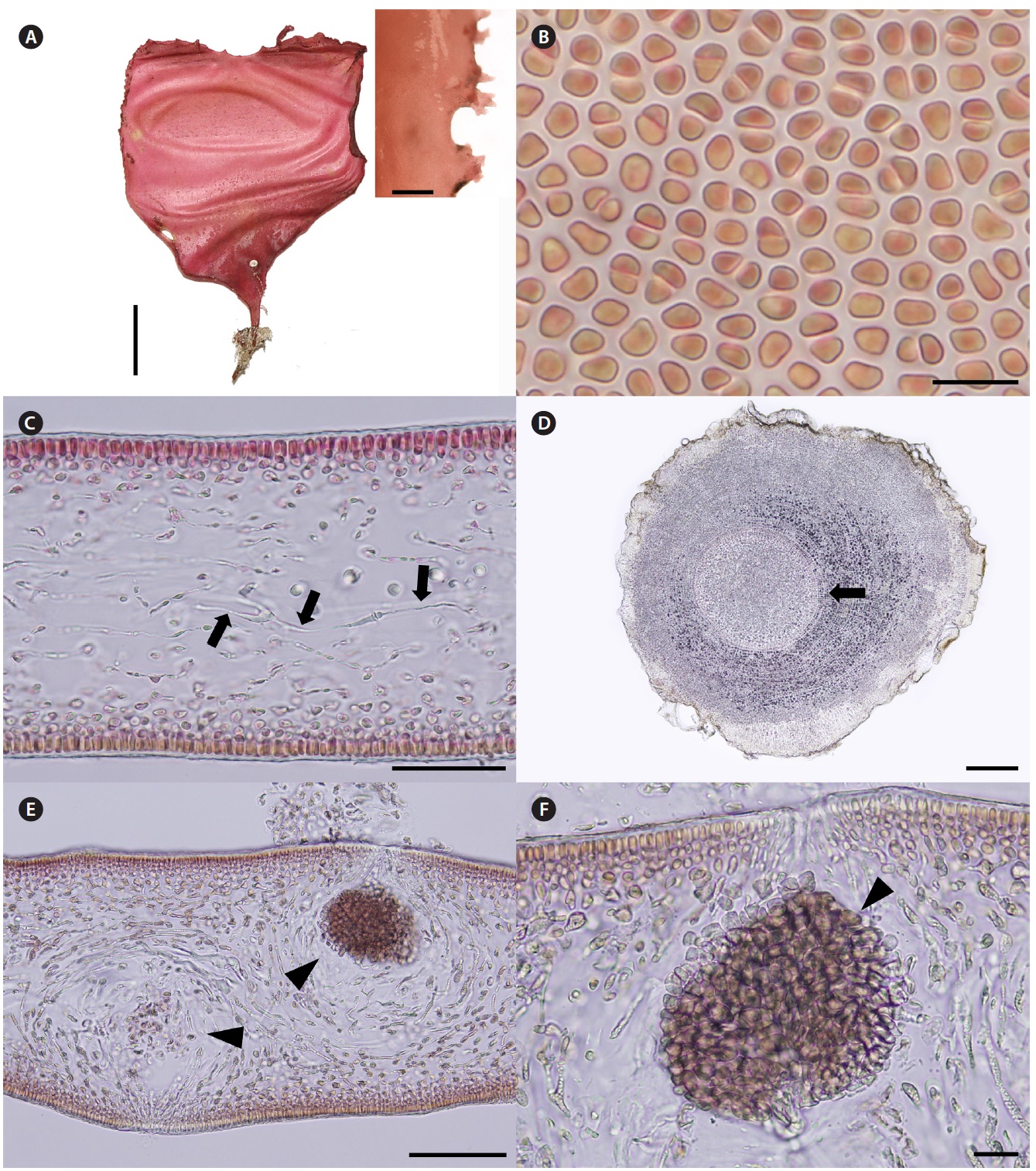
Korean name. 자루갈래잎(신칭) (Fig. 4A-F)
Description. Thalli are bright to dark red, soft fleshy, circular or ellipsoid with undulated margins, up to 32 cm in length and 15 cm in width (Fig. 4A). Cells at the sur-face of blade are irregularly elongate (Fig. 4B). Tissues are compactly composed, and 300-330 μm thick (Fig. 4C). Cortices are 5-8 layers of cortical cells and 40-60 μm thick. Medulla are composed of thin filaments that are densely interwoven with one another, and 200-240 μm thick.
Female reproductive structures are scattered over en-tire blade, and easily distinguished by visible ostioles (Fig. 4D). Cystocarps (Fig. 4E & F) are subglobose, 40-80 μm across. Spermatangia and tetrasporangia are not found.
Thalli grow on the rock with coralline algae at intertidal zones. They were collected in February, March, and April from 2008 to 2011.
Plastid
The second species added to the algal inventory of Ko-rea is
All specimens of Korean
Together with the previous study by Gabriel et al. (2011), our result demonstrates that both