Effects of forest thinning on community level properties have not been understood yet in Korea. We investigated regen-eration patterns and trajectories after a disturbance by applying a chronosequence approach. Light availability, litter and woody debris cover, and species composition were determined for twenty 50 m line-transect samples representing a dis-turbance duration gradient (within 11 years). Environmental factors such as light availability and coverage of woody de-bris and litter changed abruptly after thinning and then returned to the pre-disturbance state. Although species richness was gained at shrub and ground layer in a limited way in both forests, cover of various functional types revealed diversity in their responses. Notably, Alnus firma stands exhibited a larger increment of cover in woody plants. Ordination analysis revealed different regeneration trajectories between natural and planted stands. Based on ordination analysis, rehabili-tated stands showed movement to alternative states compared with natural ones, reflecting lower resilience to perturba-tion (i.e., lower stability). Our results suggest that community resilience to artificial thinning depends on properties of the dominant species. But to get more explanatory ecological information, longer-term static observations are required.
In Korea, forest tending works (thinning) for natural and planted forests have been conducted on a broad scale after International Monetary Fund (IMF) financial sup-port (Woo 2003). In this operation, canopy compositors (as the dominant species) are partially removed (by visu-ally selecting healthy or degrading individuals), and sub-canopy and shrub layer species are eliminated to improve growth of target species and environmental factors such as water retention capacity and soil physical properties. Canopy openness remained at 50-80% compared with the pre-thinned state (Korea Forest Service instruction No. 890). Thus, structural and environmental changes due to thinning are smaller than to clear cutting.
Management activities (clear cutting, thinning and so on) in forest ecosystems are broadly applied in most nations. Tremendous disturbances such as clear-cutting usually alter the structure and function of forest commu-nities. As a result, various successional stages commonly appear. Thinning also changes understory vegetation indirectly by modifying overstory openness and density and their effects on understory microclimate, light, and soil water (Dodson et al. 2008, Jung et al. 2008). In addi-tion, thinning may result in higher availability of water and mineral nutrients through the formation of root gaps (Parsons et al. 1994) and increased competitive perfor-mance of residual or invaded species. In managed forest ecosystems, residual species and rapidly establishing in-vaders are primary contributors to secondary succession after disturbances (logging) (Halpern 1988, 1989).
Forest operations result in various responses of plant communities, such as increasing (Metlen and Fiedler 2006, Dodson et al. 2008), decreasing (Metlen et al. 2004, Fule et al. 2005) species richness or neutral response (Nel-son et al. 2008). Responses of species cover also varied among functional types, suggesting differences in toler-ance or resilience to disturbances (Halpern 1988, Metlen et al. 2004, Metlen and Fiedler 2006, Moore et al. 2006). In understory vegetation, various consequences of second-ary succession after forest management are predictable, but the emerging patterns and pathways of disturbed for-ests are not yet clarified in natural and planted forests in Korea.
Communities may be considered stable if these path-ways, after management activities, result in a return to the initial composition within an appropriate time scale (Sutherland 1981). In this perspective, understory com-munities play a central role in the dynamics and func-tioning of forest ecosystems by influencing long-term patterns of vegetation development (D’Amato et al. 2009). Understory vegetation is also a critical component of for-est ecosystems, typically supporting the vast majority of components of total ecosystem floristic diversity (Halp-ern and Spies 1995, Thomas et al. 1999, D’Amato et al. 2009).
As was mentioned above, the effects of forest manage-ment practices in Korea such as clear cutting and thin-ning on species diversity and composition have not been understood sufficiently (but see Cho et al. 2011). This insufficiency and lack of basic information reflects the paucity of community-level studies in natural and man-aged stands. Combined with emerging principles in the landscape and in restoration ecology of natural or man-aged ecosystems, changes in the society’s conceptions of a broader range or monotonous forest management op-tions are causing a reassessment of forestry practices. For-est and natural resource managers are making an effort to balance economical and ecological values of the forest by applying alternative models of forest management. In this viewpoint, the introduction and founding of manage-ment and operation options and policies surely require a fundamental understanding of vegetation processes and conceptual models that can predict responses to pertur-bations in target systems.
The chronosequence approach to study vegetation dy-namics has been validated (Foster and Tilman 2000), and has provided significant insights into the patterns and mechanisms of plant succession (e.g., Inouye et al. 1987, Pickett 1989, Lee et al. 2002, 2008, Lee 2006a, 2006b, Hau-go and Halpern 2007, Cho et al. 2011). The present study aims to clarify how management activity influences com-munity resistance and resilience. In order to arrive at this goal, we analyzed successional patterns and trajectories (pathways) of understory vegetation and abiotic variables (light availability, woody debris and litter fall) by applying a chronosequence approach after forest thinning in natu-ral and planted broadleaved forests.
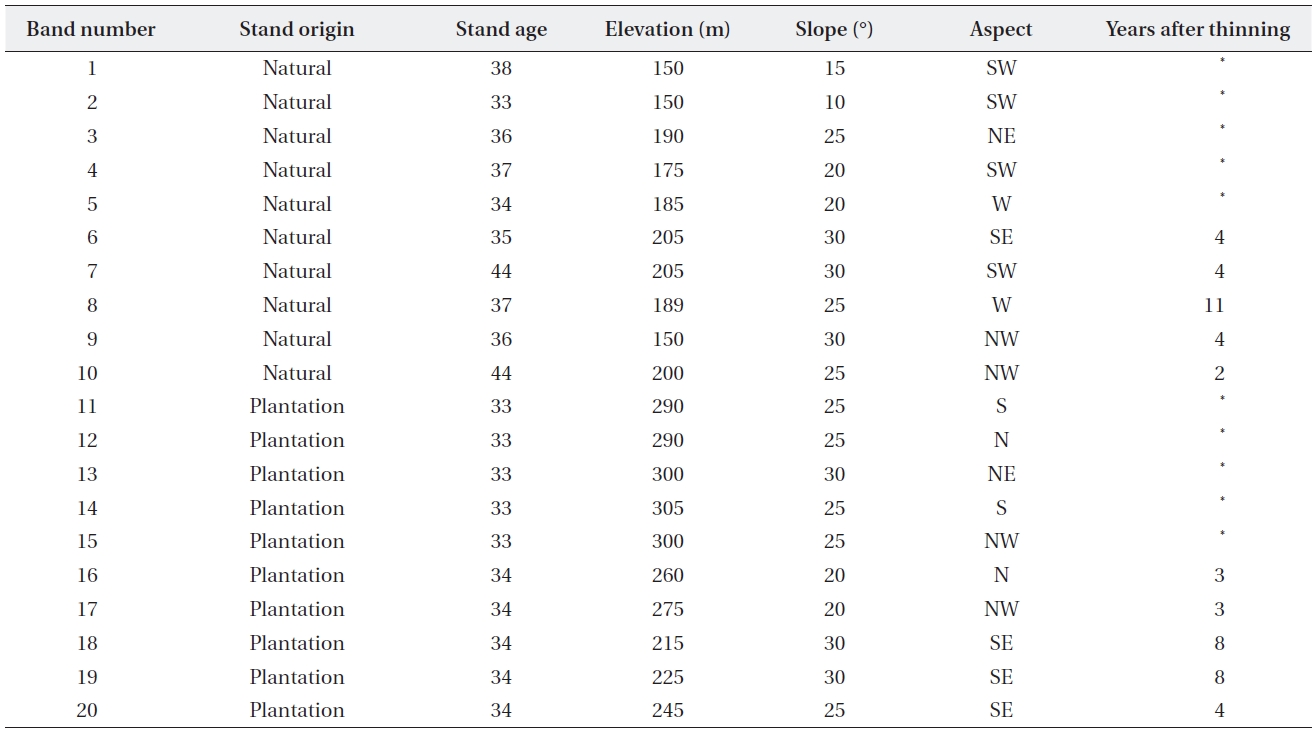
The study sites were located in rural areas in Pohang-si, Gyeongsangbuk-do, southeastern Korea. The altitudinal distributions of sites were between 150-305 m above sea level (Table 1). The study area is classified as a warm-tem-perate deciduous broad-leaved forest (such as
Soils were shallow, sandy loams and loams derived from mudstone and shale on the tertiary deposit in the Cenozoic Era based on digital detailed soil maps pub-lished by the Rural Development Administration. Annual mean temperature and precipitation (1971-2007) ob-tained from the Pohang-si weather station, the closest to this study area, were 14.0°C and 1,166.8 mm, respectively.
We explored the study area in a vegetation survey and selected 10 stands in the natural broadleaved forest (dom-inated by
For rehabilitated stands, a total of 10 stands were se-lected in
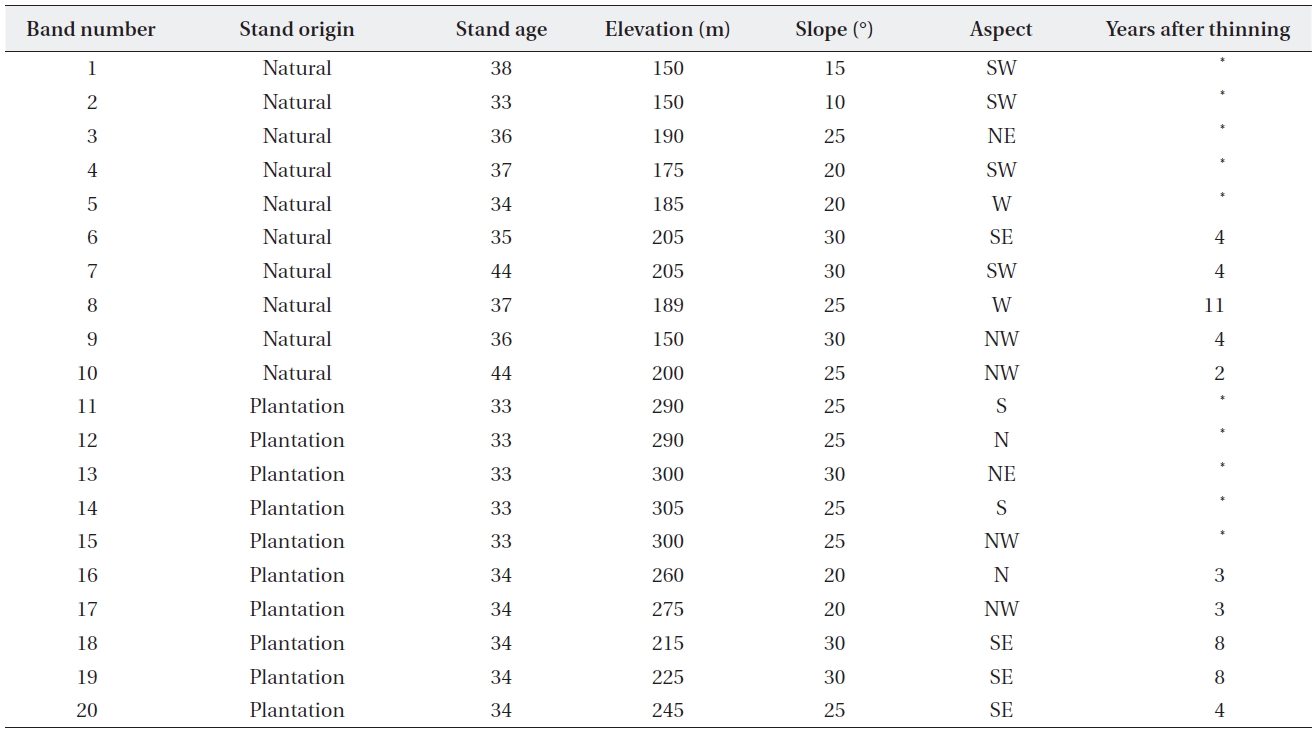
[Table 1.] General environmental features of 20 vegetation survey sites
General environmental features of 20 vegetation survey sites
in the herbaceous layer.
>
Vegetation and abiotic variables sampling
Vegetation sampling was conducted from August to September 2008. In selected stands, a 50 m line transect was installed parallel with the contour and was > 30 m from all stand edges. The transect was consisted of five plots (5 m × 5 m) spaced at 5 m intervals and 10 subplots (1 m × 1 m). Within each plot (1-2 m in height) and sub-plot (< 1 m in height), coverage (%) of all vascular species was recorded. Coverage of litter and woody debris was also determined in subplots. The age of each stand was estimated by counting tree rings from annual ring cores for the biggest tree species in the plot. Mean ages of
In order to estimate light availability and canopy open-ness (%), we took digital hemispherical photographs at each subplot from a height of 1.8 m using a Nikon D80 digital camera (Sendai, Japan) with Sigma 4.5mm F2.8 EX DC Circular Fisheye HSM lens (Fukushima, Japan). Pictures were taken on uniformly overcast days to avoid direct sunlight. Total transmitted light or photosynthetic photon flux density (mol m-2 day-1) averaged over a year was calculated with Gap Light Analyzer 2.0 (Mansfield, CT, USA) employing the standard overcast sky model (Frazer et al. 1999, Haugo and Halpern 2007).
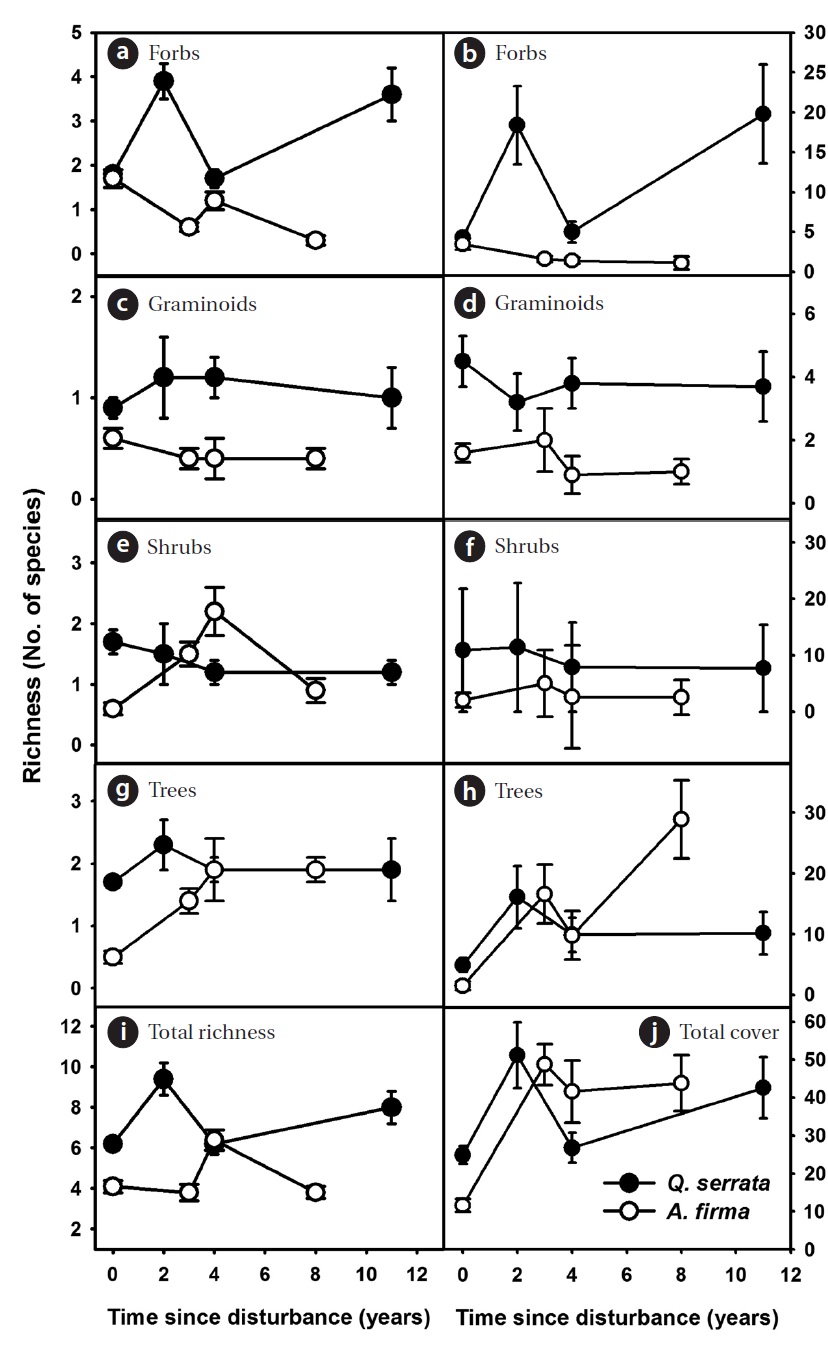
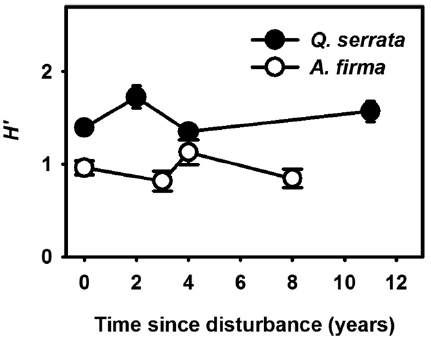
Functional types such as growth form and life trait (or habitat affinity) are often used to generalize species’ per-formance (Smith et al. 1987). To compare regeneration patterns after thinning, each species recorded was classi-fied into five growth forms, i.e., tree, shrub, forb (includ-ing ferns), graminoid and vine, and mean numbers and coverage of each form to samples of focal successional stage were calculated. Average coverage of litter and wood debris (WD), and light availability were also computed for each successional stage. In addition, in order to assess species diversity, a Shannon index (
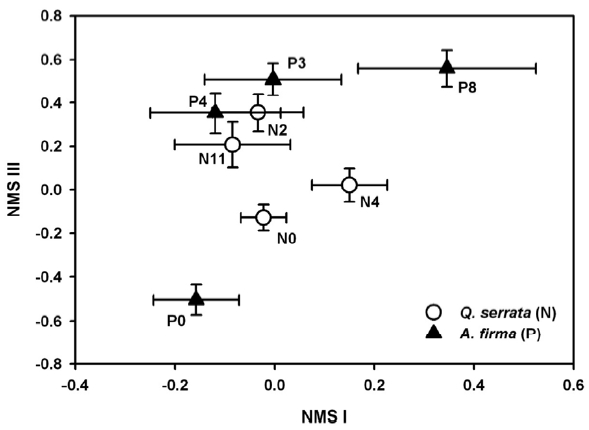
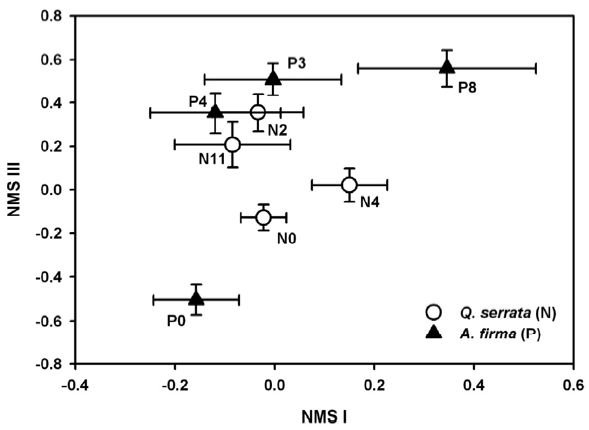
To analyze successional pathways of stands, the impor-tance value of each life form in each subplot was calculat-ed. The values were determined by multiplying 100 by the fractional value of each life form to the summed values of all functional types in each subplot. Then, a matrix of importance values for life forms was constructed and that data were fed to ordination analysis. We used Nonmetric Multidimensional Scaling (NMS) (Kruskal 1964) ordina-tion to examine differences in successional pathways due to management activities in the understory of natural and planted stands. NMS ordination was performed us-ing Søresen’s distance measure and we selected the “slow and thorough” autopilot of PC-ORD ver. 4.0 (MjM Soft-ware, Oregon, USA) conducting 40 runs that yield one to six dimensional solutions (McCune and Mefford 1999). A Monte Carlo test was performed to compare the stress of randomized data (McCune and Grace 2002). In the re-sult of NMS ordination, the minimum stress configura-tion included three axes, but the two axes accounting for the most variability in the original data set are presented (NMS I as 22.2% and NMS III as 34.7%). SPSS ver. 15.0 (SPSS Inc., Chicago, IL, USA) and PC-ORD ver. 4.2 (Mc-Cune and Mefford 1999) were applied to univariate and multivariate analyses, respectively.
>
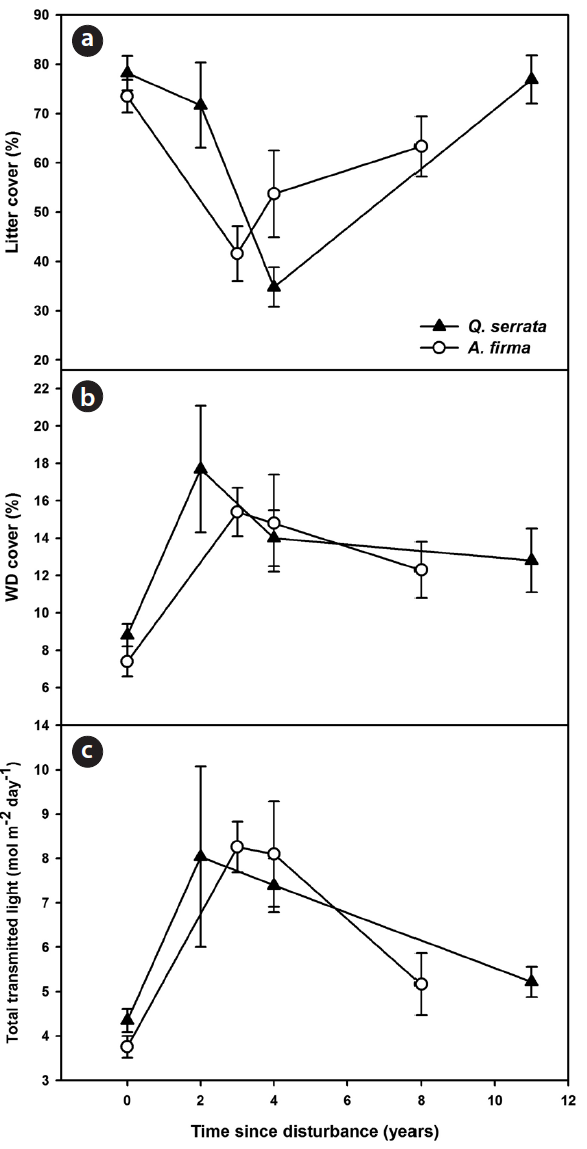
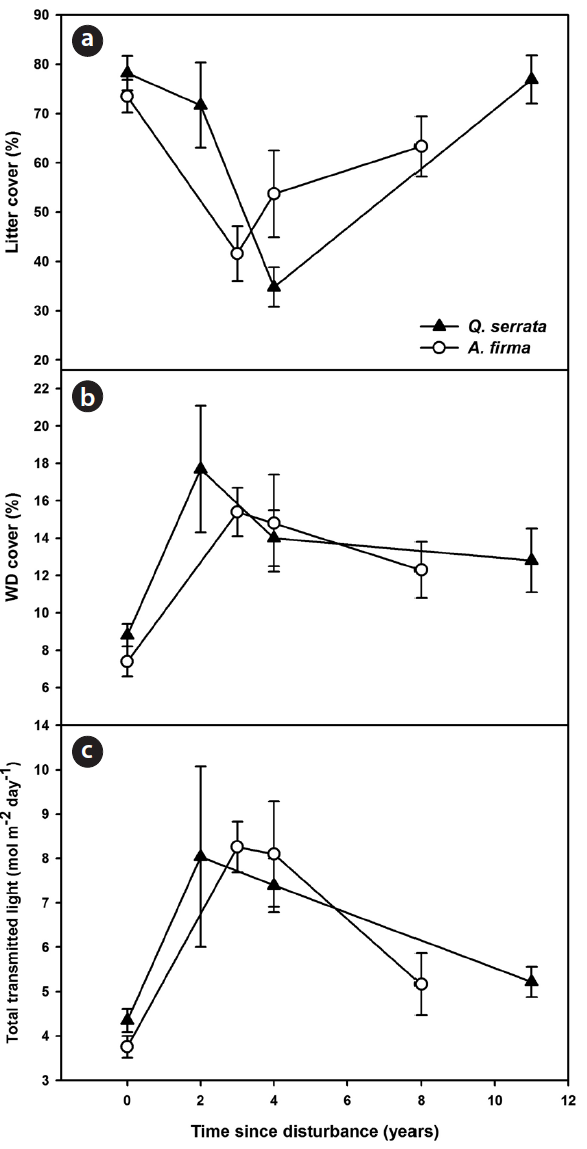
Changes in abiotic environments
Canopy conditions (estimated light availability) and ground conditions (coverage of litter and WD cover) changed dramatically in the earlier stages after thinning, and as years passed, means of environmental variables gradually approached the control state (Fig. 1). Coverage of litter decreased immediately after thinning and then approached a reference state with the passage of time in both
Ordination results explained 56.9 % of the variation in the raw data (final stress = 17.3, final instability = 0.0297). NMS ordination of subplots was expressed in mean (± standard error) of successional stages (Fig. 2). Variation in vegetation among successional stages was explained by NMS I (22.2%), which represented a gradient from sites with higher coverage of vine species in the negative pro-portion of the NMS I to those with higher scores of tree species in the positive portion. The arrangement of sam-ples along NMS III, which explained 34.7 % of the varia-tion, ranged from sites with higher coverage of graminoids to sites with higher cover of tree species. NMS ordination revealed different successional trajectories depending on forest type. Understory composition of natural stands (
>
Changes in species diversity and abundance
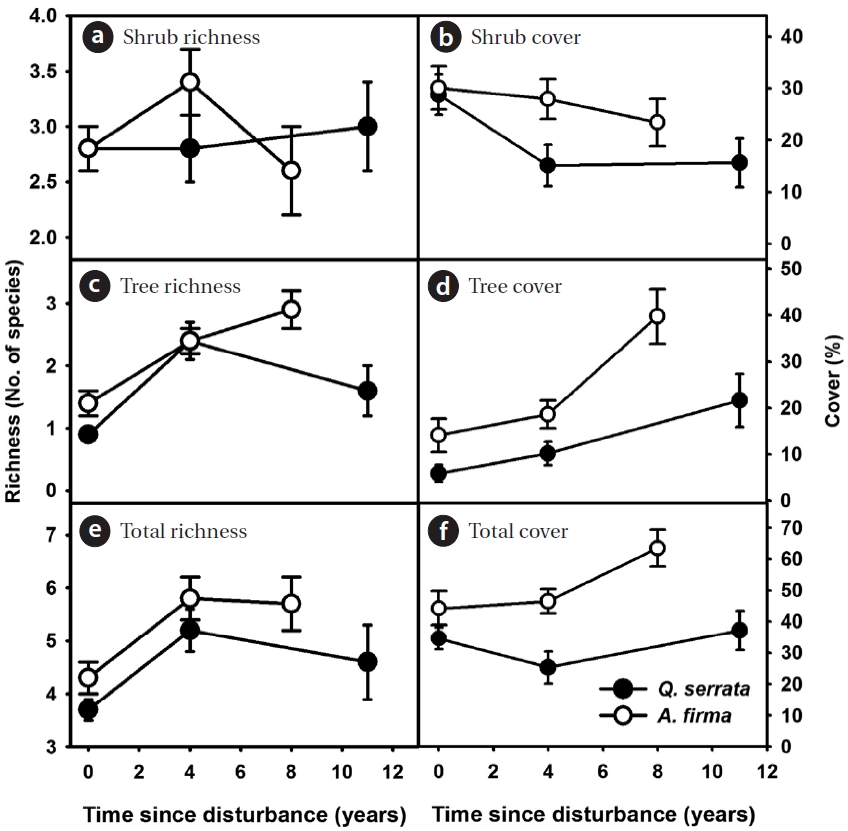
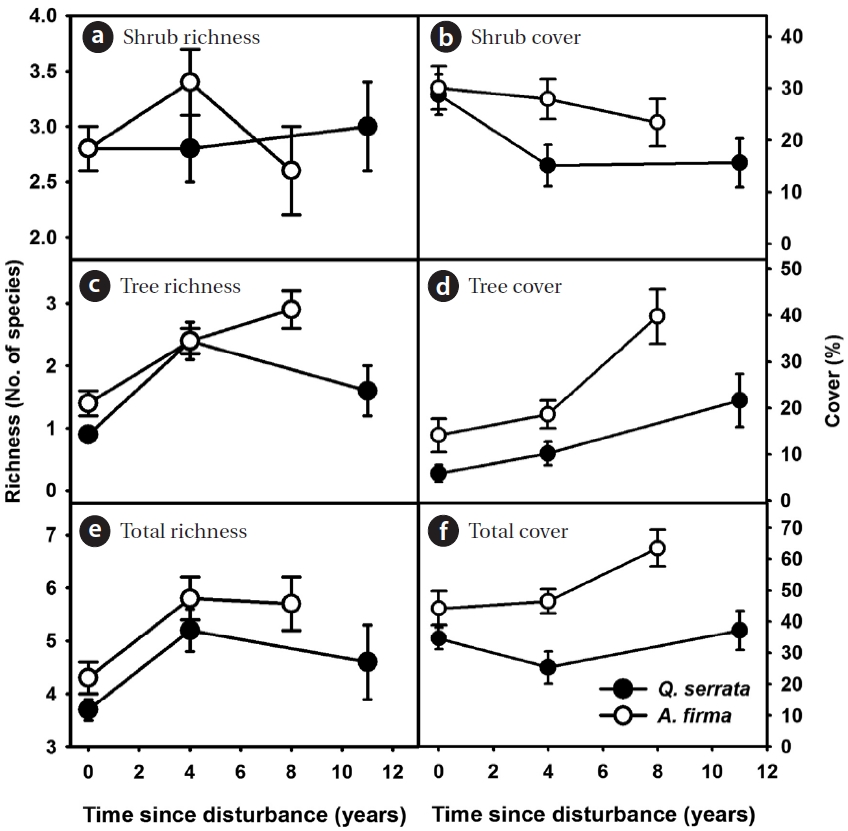
In the shrub layer, the increase in overall richness was limited after thinning (see scale of y axes, Figs. 3a, 3c, and 3e). The change in total coverage was affected largely by
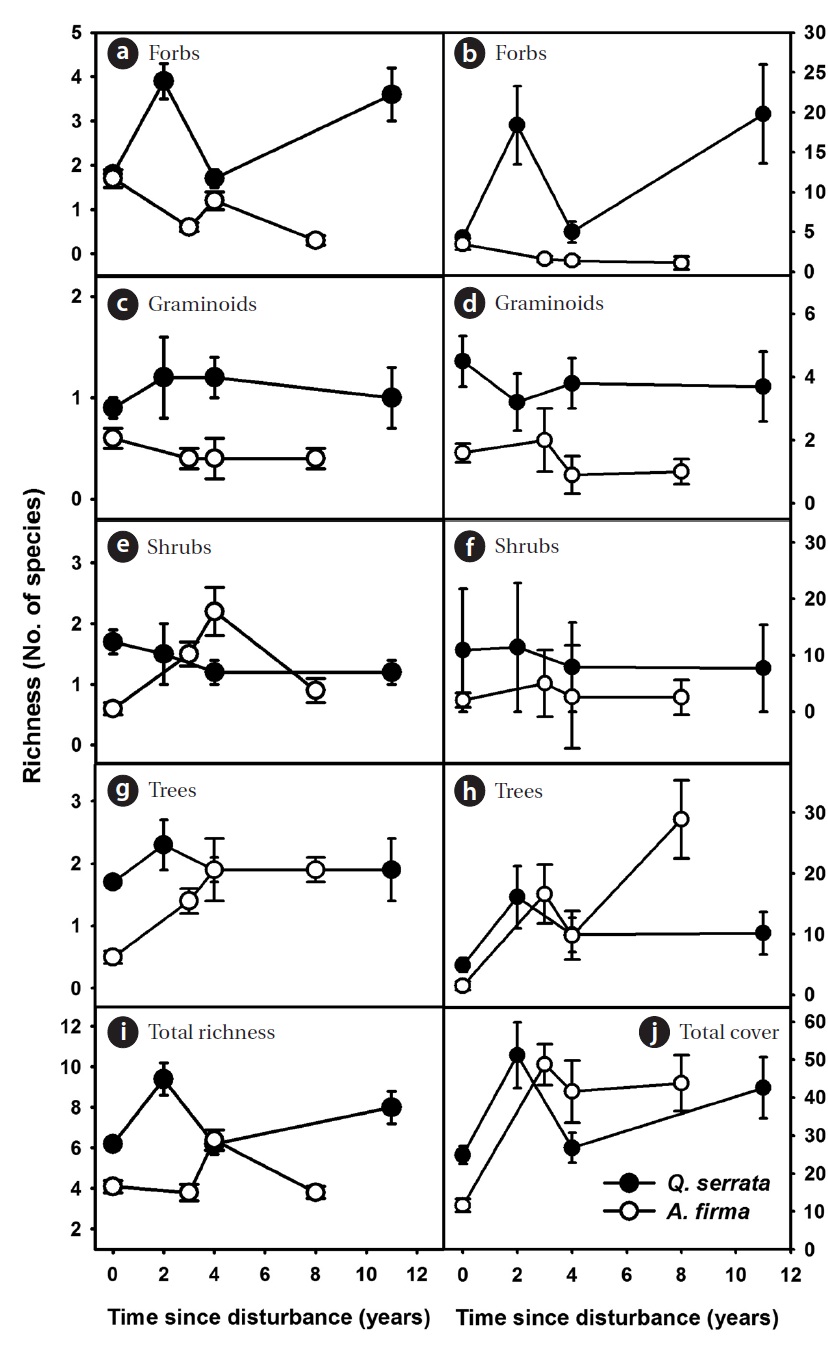
the gradual increase of tree species and the decrease of shrubs (Figs. 3b, 3d, and 3f). A limited gain in species rich-ness after thinning was observed in ground vegetation (Figs. 4a, 4c, 4e, 4g, and 4i). Vine species were excluded from analysis because of too little richness and coverage. In
Overall, total coverage of ground vegetation increased after thinning (Figs. 4b, 4d, 4f, 4h, and 4j). Peak coverage of ground communities appeared during earlier stages in both
and gained again. In
We applied a chronosequence approach to illustrate community resilience and resistance to a thinning distur-bance by showing the patterns (changes in species rich-ness and abundance) and pathways of
Tending works brought various changes in forest struc-ture and ground environments. Release of tree density resulted in alteration of light and nutrient products such as decrease of litter fall (Jung et al. 2008). Coverage of ground litter decreased in earlier stages and more than 10 years were required to restore the normal state in both forests (Fig. 1). Increases in ground woody debris in earlier stages are likely due to tree felling operations. Decreases in woody debris in later stages were due to decomposition and reduction of debris production. In addition, loss of rainfall interception because of thinning (Jung et al. 2008) may increase runoff and consequently, by wash away, can affect the amount of litter and woody debris in forests.
After thinning, species richness and diversity increased in earlier stages (within 4 years) but the magnitude was relatively limited in both
In herbaceous vegetation, forb species, which contrib-uted largely to increases in richness and total cover in natural stands, are likely able to take advantage of distur-bances in earlier stages. Different patterns of richness and coverage between natural and restored stands are largely explained by domino effects of thinning on
Community stability (or persistence) may simply re-flect the ability of resident species to resist change, or if altered by disturbances, their ability to readjust or re-cover (Halpern 1988). Different regeneration trajectories of functional type composition between stands reflect the variety of species group responses to disturbances. In addition, resistance and resilience varied between plant communities, and were likely better for natural stands and lower for plantation. The recovery pattern of the un-derstory of