Time series of ocean climate indices and catch records were used to identify the alternation patterns of pelagic fish popu-lations in relation to climate regime shifts. During 1910-2008, an orderly alternation of dominant pelagic fish groups was observed in the Tsushima Warm Current (TWC; Yellow Sea-East China Sea-East Sea/Japan Sea) and Kuroshio-Oyashio Current (KOC; Northwestern Pacific) regions. After the collapse of herring fishery in the late 1920s, the sardine (A group)dominated in the 1930s, 3 other species (C group; Pacific saury, jack mackerel, and anchovy) dominated in the 1950s-1960s, chub mackerel (B group) dominated in the 1970s, and then sardine (A group) dominated again during cool regime in the 1980s. As sardine biomass decreased in association with the climate regime shift that occurred in the late 1980s, catches of C group immediately increased after the regime shift and remained at high levels during warm regime in the 1990s. Alternations of dominant fish groups occurred 6 times between 1910 and 2008. The dominant period of the 7 spe-cies lasted for 10-20 years. The catch of Pacific sardine in the TWC and KOC regions showed a negative correlation with the catch of the other 5 species (Pacific herring, anchovy, jack mackerel, Pacific saury, and common squid), suggesting that the abundance of the 5 species is strongly affected by the abundance of Pacific sardine in relation to the climate regime shifts. The total catch level of the 7 species in the KOC region was generally higher than that in the TWC region before 1991 but was lower after 1992, suggesting that the fish populations in the Pacific side are shifted to the TWC region by zonal oscillation of the oceanic conditions in relation to the climate regime shift in the late 1980s.
Growing evidence suggests that abrupt environmen-tal regime changes can influence pelagic fish production(Lluch-Belda et al. 1989, Beamish 1995, Hare and Mantua 2000, Yatsu et al. 2005, Tian et al. 2008). Sardine landings show synchronous variations in the northwestern, north-eastern,and southeastern Pacific (Kawasaki 1983). The discovery of these biological regime shifts preceded the description of the underlying physical variability. Fluctua-tions in air temperature, ocean temperature, atmospheric circulation, and carbon dioxide were remarkably similar in phase and duration to the biological records (Chavez et al. 2003). As a result, it has been suggested that a regime or climatic shift may be best determined by monitoring marine organisms rather than climate (Hare and Mantua 2000). It is also suggested that the ocean conditions were warmer than normal at the upper layers in the Tsushima Warm Current (TWC) region in the 1990s (Gong et al.2008, 2010, Tian et al. 2008). Pacific sardine, file fish, and walleye pollock disappeared from the TWC region after the climate regime shift in the late 1980s.
The abrupt changes in the fish community in the East Sea (Japan Sea) that occurred in the mid-1970s and late 1980s seemed to closely correspond with the climate re-gime shifts in the North Pacific (Tian et al. 2006). A fishery regime is defined as a decadal-scale period of high or low abundance of fish populations. Both the marine ecosys-tem as the fish population habitat and the production system as the bottom level of the food web shifted in the mid-1970s and the late 1980s in the North Pacific and its adjacent regions.
Fluctuation of fish population abundance may result from environmental fluctuation, density-dependent re-production rate, or interspecific interactions. The alterna-tion of dominant pelagic fish species in the waters off of Japan are well documented (Nakahara and Ogawa 1979, Matsuda et al. 1991, 1992, Kawasaki 1992, Watabe 1992a, 1992b, Schwartzlose et al. 1999, Yatsu et al. 2005). It was noticed that the year-to-year catch of some pelagic fish populations (e.g., chub mackerel) in the Kuroshio-Oyas-hio Current (KOC) region showed a different pattern than the catch in the TWC region after the climate regime shift in the late 1980s (Gong et al. 2008, 2010). The replace-ment patterns in relation to climate change in the waters around the Korean peninsula (TWC region) have not been fully explained compared with the patterns in the KOC re-gion due to the limited specific regional catch data.
This study investigates the temporal correspondence between the main patterns of abundance fluctuation among Far East exploited pelagic fish populations and large scale climate and temperature indices.
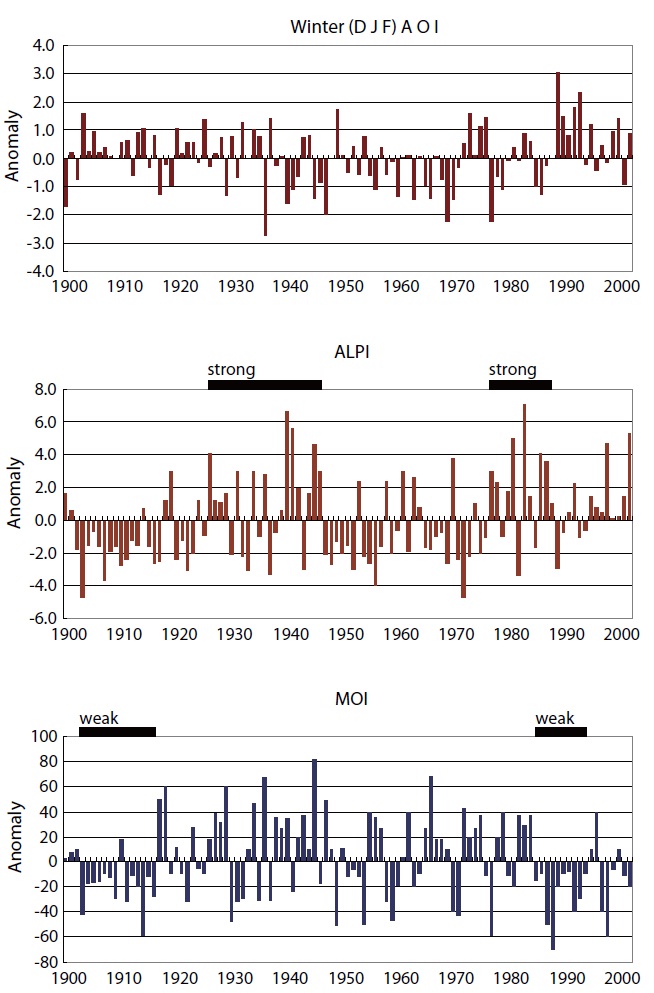
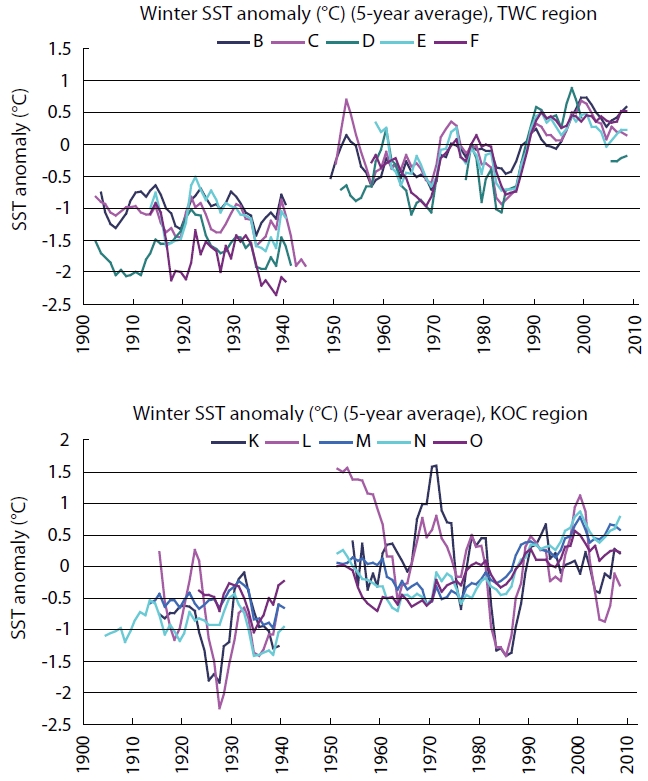
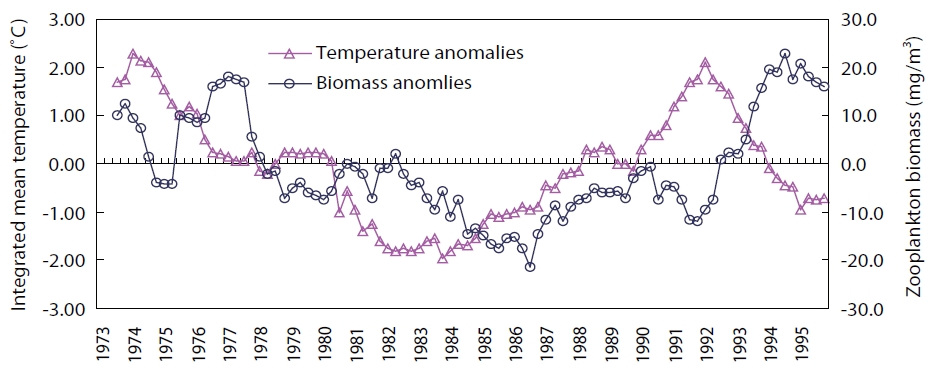
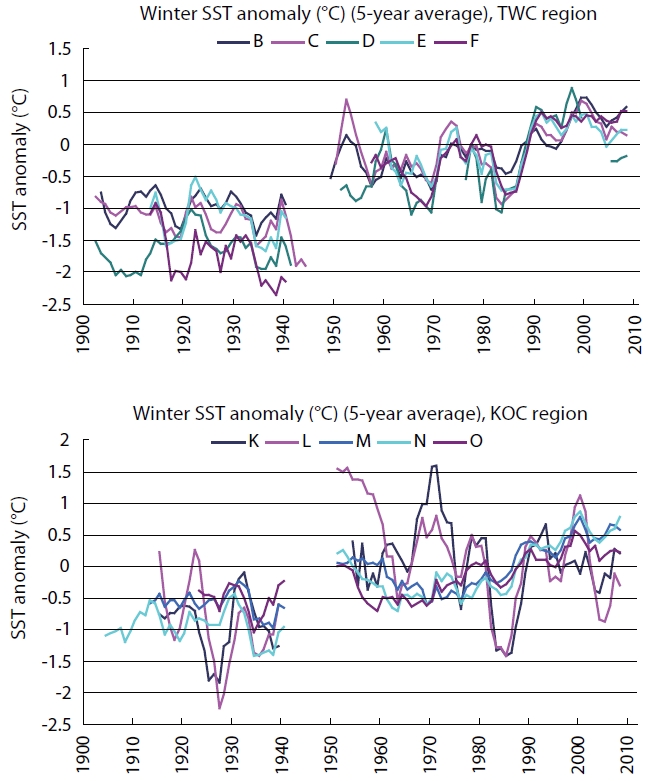
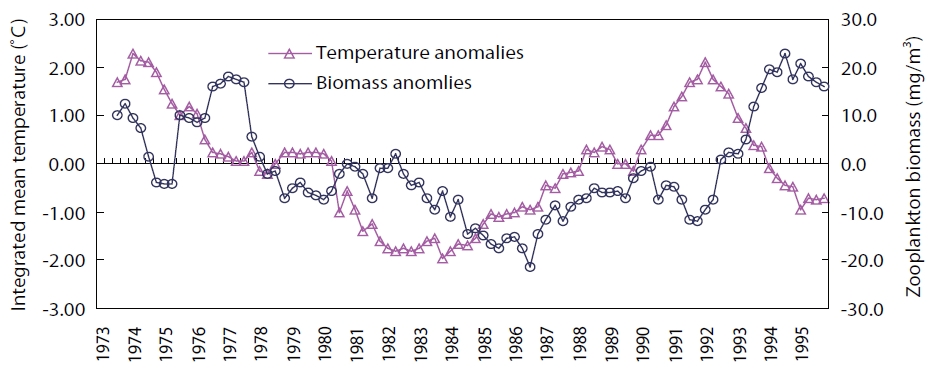
The Arctic Oscillation Index (AOI), Aleutian Low Pres-sure Index (ALPI), and Monsoon Index (MOI) were chosen as climatic indices for the Far East region (Fig.1 ). A time se-ries of winter (Jan, Feb, Mar; JFM) sea surface temperature (SST) anomalies in the TWC and KOC regions for the period of 1900-2010 (Fig.2 ) (Japan Meteorological Agency 2011) and anomalies of integrated mean temperature (0-150 m) and zooplankton biomass along the PM line (36˚ N, 136˚ E-44˚ N, 132˚ E) in the TWC region for the period 1973-1998 (Fig.3 ) (Minami et al. 1999) were used to examine the fluctuations in the oceanic conditions in relation to the cli-mate changes.
Year-to-year catch series of the 7 pelagic fishes (Pacific herring, Japanese sardine, anchovy, chub mackerel, jack mackerel, Pacific saury, and common squid) were con-structed for the TWC (East China Sea and Yellow Sea and
East Sea/Japan Sea) and KOC (Northwest Pacific includ-ing the Okhotsk Sea and Seto Inland) regions. Since the range of the pelagic fish populations is quite large, fishing records from all fishing countries in the Far East (FAO area 61; north of 20° N, west of 175° W) were included to exam-ine the long-term trend in abundance of the fish popula-tions across the entire distribution range.
The historical catch data for the period 1910-2008 are based on the Yearbook of Fishery Statistics of Korea and Japan, FAO Yearbook of Fisheries Statistics, and previous reports including fishery data (Chikuni 1985, Schwartzlose et al. 1999, Klyashtorin 2001, Gong et al. 2007).
Although catch alone is acknowledged to be a crude measure of abundance, direct surveys have only recently started in most systems, so there are no comparative indi-ces for periods of high and low abundance of the pelagic fish populations. Nevertheless, the changes in catches
have been so marked (Gong et al. 2007, 2008), there can be little doubt that they reflect real changes in population abundance.
A comparison was performed using time series of hydro-climatic (AOI, ALPI, MOI, and SST) anomalies and
biological parameters (zooplankton biomass and year-to-year catch as abundance indices). Correlation coef-ficients between catch time series for the 7 species were calculated to examine the patterns in fluctuations among fish popu-lations. Alternations of the dominant fish populations were examined for the TWC, KOC, and TWC + KOC regions.
>
Climate and ocean regime shifts
The AOI showed decadal-scale variation patterns: dis-tinct changes in 1971, 1977, and 1989. The wintertime ALPI showed a distinct change in 1977 from negative to posi-tive values, indicating strengthening of the Aleutian Low after 1977. The wintertime MOI showed an abrupt change around 1987 from positive to negative anomalies, indicating the weakening Asian monsoon in the 1990s, while only slight changes occurred in the mid-1970s. The abrupt changes that occurred in 1976-1977 are common for AOI and ALPI, while the changes that occurred in the late 1980s (1988-1989) are common for AOI and MOI. These 3 indices also point to pos-sible climate changes in the late 1990s (Fig.1 ).
The winter (JFM) SST anomalies (5-year running aver-age) in most areas of the TWC and KOC regions were far below the long-term average in the earlier period (1900s-1940s). The anomalies in the TWC region were below the average in the late 1960s and 1980s but far above the av-erage in the 1990s and 2000s. The winter SST anomalies in the northern KOC region (Area K, Sanriku; and Area L, Kanto-east) were above the average in the 1950s and late 1960s and below the average in the 1980s. The winter SST anomalies in the TWC and KOC regions were above the average after the climate regime shift (1988-1989) and re-mained at high levels in the 1990s and 2000s (Fig.2 ).
A positive MOI (+MOI) is associated with anomalously high pressure over the Asian Continent and intensified northwesterly winds which co-varies with the deep ALPI (+ALPI) and negative AOI (-AOI). Consequently, the in-tensified northwesterly winds transport cool and dry air masses over the Far East, which results in a severe winter and cool regime in the TWC and KOC regions as seen in the 1980s. The opposite situation is marked by a nega-tive MO index (-MOI), indicating reduced northwesterly air flow. Simultaneously, the inflow of cold and dry air masses from the Asian Continent is reduced, resulting in mild winters and warm SST over the Far East as seen in the 1990s (Figs. 1 and 2).
Winter temperatures in the 50-m layer in the East Sea (Ja-pan Sea) showed decadal variation patterns together with large inter-annual variations (Katoh et al. 2006). The cool regime in the 1980s and warm regime in the 1990s are clearly shown in the year-to-year strength anomalies of the TWC defined by the area >10°C in the 100-m layer in the East Sea (Japan Sea) during the period of 1961-2007 (Japan Meteorological Agency 2009).
The MOI is significantly correlated with winter SST in the TWC and KOC regions, suggesting that ocean variability is linked with the winter Asian monsoon. The oceanic shifts from the warm to cold regime in the mid-1970s and from the cold to warm regime in the late 1980s are associated with the climate regime shifts.
The inter-decadal variability of integrated (0-150-m layer) mean temperature and zooplankton biomass along the PM line in the TWC region was positively correlated with the 3-year time lag. Both variables were low during the 1980s after the climate regime shift (1976-1977) but increased after the regime shift (1988-1989) and remained higher than average during the warm regime in the 1990s (Fig.3 ).
The abrupt climate changes that occurred in 1976-1977 are common for AOI and ALPI, while the changes occurred in the late 1980s (1988-1989) are common for AOI and MOI (Tian et al. 2004, King 2005). Year-to-year SST anom-alies in the southern East Sea (Japan Sea; 34.5˚ N-38˚ N) denoted the cool regime (1920s-1930s and 1980s), warm regime (1950s and 1990s-2000s), and the long-term in-creasing trends in the last century (Fig.2 ). There was a cold period from the mid-1970s to the late 1980s, and then temperatures were high from the late 1980s to the early 2000s in the TWC region in the East China Sea and southern East Sea (Japan Sea) (Naganuma 1992, Katoh et al. 2006).
Decadal averaged winter SST anomalies in the East Sea (Japan Sea) sawed a see-saw variation pattern spatially. Cold anomalies were limited to the TWC region in the south in the 1970s but then extended to the whole East Sea (Japan Sea) during the 1980s. The whole region be-came warm in the 1990s, and then the warm areas were reduced to the south during the 2000s. The time series of diatom abundance in the East Sea (Japan Sea) showed decadal variability and large inter-annual variations. Step change occurred in the spring diatom in 1979-1980 and 1991-1992 and in the autumn diatom in 1995-1996 (Tian et al. 2008).
Six climate regime shifts were detected in the northern hemisphere SST fields during the period from the 1910s to the 1990s: 1925-1926, 1945-1946, 1957-1958, 1970-1971, 1976-1977, and 1988-1989 (Yasunaka and Hanawa 2002). The 1976-1977 and 1988-1989 regime shifts have been de-scribed as major shifts having Pacific-wide impacts, while the 1957-1958 and 1970-1971 regime shifts have been de-scribed as minor shifts with limited ecological impacts (Yatsu et al. 2008). The time series of winter SST and the surface current velocity in the Kuroshio axis in the KOC region denoted the oceanic regime shifts in the mid-1970 and late 1980s (Tian et al. 2004). These results suggest that a distinct regime shift in the oceanographic conditions in the TWC region occurred in the 1976-1977 and 1988-1989 regime shifts.
The effects of the 1976-1977 and 1988-1989 climate re-gime shifts on regional ecosystems of the Northwestern Pacific (KOC region) from winter to spring can be sum-marized as follows:
After the climate regime shift in the mid-1970s (1976-1977), strong winter wind stress was associated with the deep Aleutian Low enhanced vertical mixing and cool-ing in the Oyashio region. Limited light intensity in the winter-spring seasons resulted in decreased biological production and delayed springtime blooming. The deep Aleutian Low enhanced vertical mixing and nutrient sup-ply in the Kuroshio/subtropical region. Biological produc-tion deceased in the winter but increased in the spring, delaying the springtime bloom (Yatsu et al. 2008).
After the climate regime shift in the late 1980s (1988-1989), weak wind stress under the weakened Aleutian Low limited vertical mixing in the Oyashio region. Biological production increased in the winter-spring seasons under the strong light intensity and the springtime bloom oc-curred earlier than usual. As a result of the limited vertical mixing and nutrient supply in the Kuroshio/subtropical region, biological production increased in the winter but decreased in the spring. However, the springtime bloom-ing occurred earlier than usual (Yatsu et al. 2008).
>
Distribution of pelagic fish populations and fish-ing nature
The 7 pelagic fish populations (Pacific herring, Pacific sardine, Japanese anchovy, jack mackerel, chub mackerel, Pacific saury, and common squid) inhabit the waters from the subtropic to the subarctic zone in the Northwest Pa-cific and its adjacent regions. They spawn in the southern part of their distribution range in the autumn, winter, and spring and then migrate to the north in the summer to feed in the northern part of their population area.
Pacific herring (
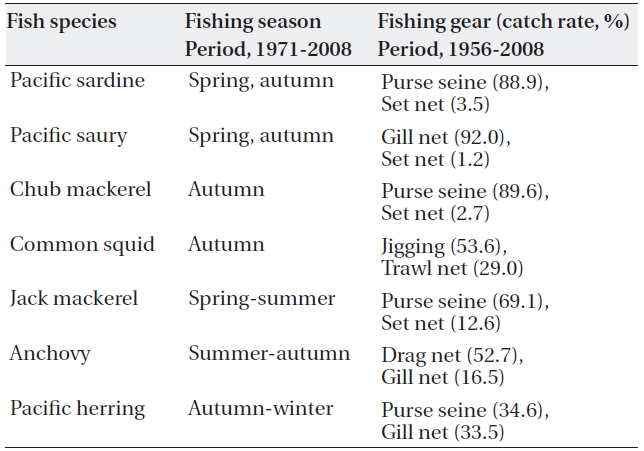
Most of the 7 species are fished at the midpoint of their northward or southward migration. Fishing seasons of the 7 pelagic fishes are fairly different in the northwestern TWC region (off Korea). Chub mackerel (
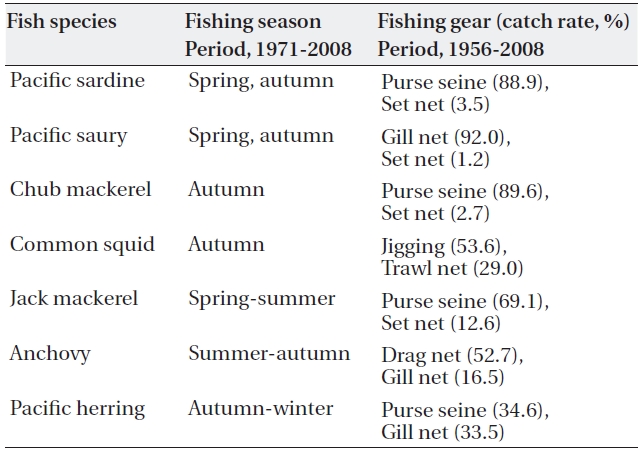
Main fishing seasons and catch rates of the 7 pelagic fishes taken by main fishing gears in the western TWC region (off Korean peninsula)
some (about 30%) by trawl in recent years (National Fish-eries Research and Development Institute 2010).
Except in the main fishing seasons, most of the 7 pe-lagic fish species emigrate from their fishing grounds. Table 1 shows main fishing seasons and catch rates of the 7 pelagic fishes taken by main fishing gears in the western TWC region (off Korean peninsula ).
>
Regional catch of the pelagic fish populations
Records of the fisheries targeting the 7 pelagic fish pop-ulations in the FAO Statistical Area 61 (north of 20° N, west of 175° W) date back to the early 1900s. Since the fishing operations by a certain country covers only small part of a population’s large range, the catch rates of pelagic fish species by the fishing countries have remarkable monthly variations. Therefore, we collected all of the catches for the 7 species from all fishing countries in the Far East.
Year-to-year catches of the 7 pelagic fish populations from the FAO Area 61 are divided into 2 regions, the TWC (Yellow Sea, East China Sea and East Sea/Japan Sea) and the KOC (Northwestern Pacific and Okhotsk Sea), for the period of 1910-2008. The catches by Korea were from the TWC region and catches by Japan were from the KOC, and the TWC regions before 1951 and the catches after 1952 are from the TWC and KOC regions by all of the fishing countries (China/Taiwan, Korea, Japan, and Russia).
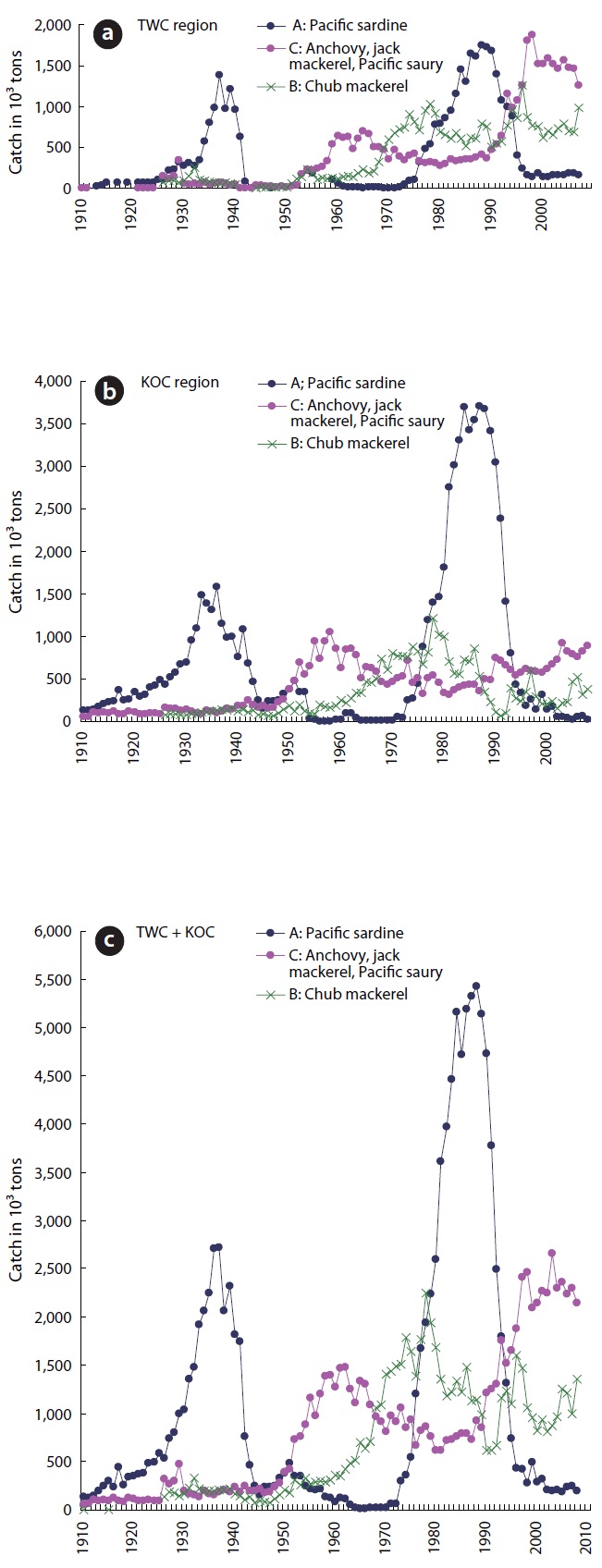
Year-to-year total catches of the 7 pelagic fishes in the TWC region steadily increased from 1,000,000 tons in the early 1960s to 3,000,000 tons in the late 1980s. The catch level was high (average 3,000,000 t) with large annual fluctuations for 11 years (1988-1998) and then slightly low (2,755.000 t) with small annual fluctuations for the next 10 years (1999-2008) (Fig.4 ).
The total catch in the KOC region increased sharply from about 2,000,000 t in the mid-1970s to about 5,000,000 t in the mid-1980s. However, the catch decreased sharply in the early 1990s and then remained at the level of ap-proximately 1,500,000 t after the mid-1990s. The catch level in the KOC region was generally higher than that in the TWC region before 1991 but was lower than that in the TWC region after 1992.
Year-to-year total catches of the 7 pelagic fish popu-lations in the entire TWC + KOC (FAO, Area 61) region showed a peak amounting to approximately 3,300,000 t in the 1930s and then decreased to approximately 900,000 t in the mid-1940s. Thereafter, the total catch steadily in-creased to about 8,000,000 t in the mid-1980s and then decreased to 4,500,000 t in the 1990s and 2000s (Fig.4 ). The 2 peaks in the year-to-year total catch in the whole area (TWC + KOC) of the 7 pelagic fish populations dur-ing the cool regimes in the mid-1930s and mid-1980s are attributed to the high abundance of Pacific sardine in the entire population range.
In the TWC and KOC regions, Pacific sardine catch in-creased after Pacific herring catch decreased in the late 1920s and late 1970s, and the anchovy catch increased soon after the sardine catch decreased in the late 1980s. These patterns of fluctuation in the Pacific herring popu-lation are completely out of phase with those of the Pa-cific sardine (Nakahara and Ogawa 1979, Kawasaki 1991). We can see a series of concurrences of a sardine peak (trough) and a herring trough (peak) as well as a sardine peak (trough) and an anchovy trough (peak), meaning that the 3 species are in opposite positions with regard to the climatic change.
In the mid-1970s, the northeastern Pacific changed from a cool anchovy regime to a warm sardine regime. A shift back to an anchovy regime occurred in the mid- to late 1990s (Chavez et al. 2003). In the mid-1970s, the northwestern Pacific changed from a warm anchovy re-gime to a cool sardine regime. A shift back to an anchovy regime occurred in the late 1980s (Lluch-Belda et al. 1989, Gong et al. 2008).
The chub mackerel catch increased soon after the jack mackerel catch decreased in the mid-1960s (Nakahara and Ogawa 1979, Takeshita and Hayashi 1991). Catches of Pacific saury and common squid were high during the warm regime in the 1950s-1960s and 1990s-2000s but were low during the cool regime in the mid-1970s and the 1980s. This switching of catches suggests alternations in abundance of the species related to the climate ocean change.
A large amount (half or one third) of the squid taken in the TWC region (mainly East Sea/Japan Sea) landed at the fishing ports along the Pacific coast (KOC region) in the late 1960s and early 1970s and were recorded in the Yearbook of Fishery Statistics of Japan. The catches of squid taken in the TWC region that landed in the fishing ports along the Pacific coast were added to the TWC re-gion counts. Therefore, actual catches of common squid in the KOC region were very low in the late 1960s and early 1970s but increased after the climate regime shift in the late 1980s (Gong et al. 2006, Gong and Choi 2008).
>
Correlation among catches of the 7 fish popula-tions
Correlation coefficients among catches of the 7 pelagic fish populations are presented for the TWC, KOC, and TWC + KOC regions (Table 2).
In the TWC region, the catch of Pacific sardine shows a negative correlation with the catches of other 5 species (Pacific herring, anchovy, jack mackerel, Pacific saury, and common squid). In particular, the correlation between sardine and saury shows a significant negative value (
In the KOC region, the catch of Pacific sardine shows a negative correlation with the catches of the other 5 spe-cies (Pacific herring, anchovy, jack mackerel, Pacific saury, and common squid) as in the TWC region. The catch of anchovy shows a positive correlation with the catches of the other 3 species (jack mackerel, Pacific saury, and common squid) but a negative correlation with the chub mackerel.
In particular, the correlation coefficients between the catches of Pacific herring and sardine (
In the entire population area (TWC + KOC), the catch of Pacific sardine shows a negative correlation with the catches of the other 5 species (Pacific herring, anchovy, jack mackerel, Pacific saury, and common squid), while orthe catch of anchovy shows a positive correlation with the catches of the other 4 species (jack mackerel, chub mack-erel, Pacific saury, and common squid). Correlation coef-ficients between any pair of the common squid, anchovy, and chub mackerel are high in the TWC and TWC + KOC regions, while the chub mackerel and common squid are negatively correlated in the KOC region. The correlation coefficients between the catch of jack mackerel and Pa-cific saury (
The abundant period (1960s and 1990s) of anchovy, jack mackerel, and common squid are fairly simultaneous in the TWC + KOC region as evidenced by the catch trend
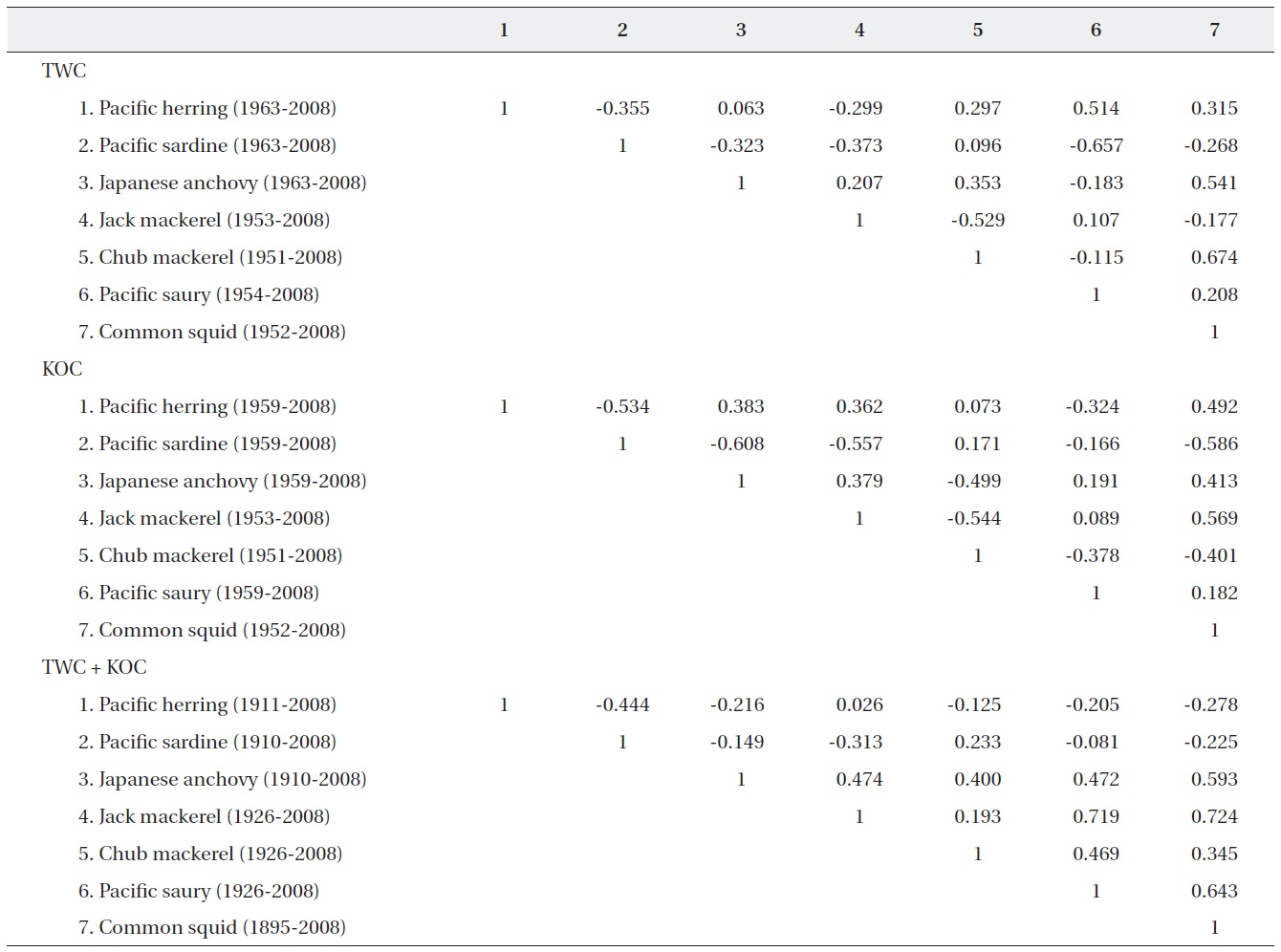
Correlation matrix of the annual catch fluctuation of the 7 pelagic fish populations in the Tsushima Warm Current (TWC) Kuroshio-Oyashio Cur-rent (KOC) and TWC + KOC regions
and fairly high correlation coefficients. The catch of Pacif-ic sardine shows a negative correlation with the catches of the other 5 species (Pacific herring, anchovy, jack mack-erel, Pacific saury, and common squid) in the TWC, KOC as well as in the TWC + KOC regions, suggesting that the 5 species are strongly affected by the Pacific sardine in the entire population area.
>
Alternation of the dominant fish species
Since the catches of anchovy, jack mackerel, and Pacific saury are negatively correlated with that of Pacific sardine (Table 2) and the catches of all 3 species are comparative-ly small, a combined catch of the 3 species (C group) and the catch of chub mackerel (B group) were compared with that of the Pacific sardine (A group) (Fig.5 ).
During the long period of 1910-2008 (99 years), an or-
derly alternation of the dominant pelagic fishes groups was observed in the TWC, KOC, and TWC + KOC regions. After the collapse of herring fishery in the late 1920s, the sardine (A group) dominated in the 1930s, and 3 other species (Pacific saury, jack mackerel, and Japanese an-chovy) (C group) in the 1950s-1960s, chub mackerel (B group) in the 1970s, and then sardine (A group) again in the 1980s. As sardine (A group) biomass decreased in as-sociation with the climate shifts in the late 1980s, catches of the 3 other species (C group) increased immediately in the warm regime from the late 1980s to the 1990s (Fig.5 ). The dominant period of the 7 species lasted for 10-20 years. However, a comparatively low catch of chub mack-erel (C group) in the KOC region in the 1990s was noticed, suggesting that the fish population in the Pacific side de-clined or shifted to the TWC region (Gong et al. 2008).
Small pelagic schooling fish can respond dramatically and quickly to climatic variations (Hunter and Alheit 1995). Most such species are highly mobile, rely on short plankton-based food chains, are highly fecund, and have plasticity in growth, survival, and other life history traits. These biological characteristics make them sensitive to environmental forcing and are highly variable in abun-dance (Kawai 1987, Alheit and Hagen 1997). It has been suggested that most of the species exhibiting alternation of dominance are planktivore pelagic fishes that migrate in the large range. Pacific sardines feed on phytoplankton and zooplankton, while most of the small pelagic fishes feed on zooplankton and/or juveniles of fish.
An orderly succession of dominant (i.e., exhibiting greatest abundance) pelagic species has been observed in the KOC and California current systems (MacCall 1996), in the Humboldt system, and off the coast of South Africa. It is suggested that small pelagic fishes exhibit large expan-sion and contractions of range concomitant with changes in abundance (Nakahara and Ogawa 1979, Lluch-Belda et al. 1989, Kuroda 1991, Watabe 1992a, 1992b).
Chub mackerel and sardine extend their habitats from the north to the south; on the other hand, jack mackerel and anchovy extend from the south to the north when their population strengths increase in the waters off of Ja-pan, suggesting that the former group developed in the northern waters while the latter group developed in the southern waters (Nakahara and Ogawa 1979). The ocean-ic shifts from warm to cold regimes in the 1970s and from cold to warm regimes in the late 1980s are associated with the climate regime shifts. During the cool regime between the regime shifts in 1977-1978 and 1988-1989, low tem-perature from the surface to the subsurface water in as-sociation with the deep Aleutian Low Pressure enhanced the phytoplankton bloom in the spring; hence, the Pacific sardine population increased. During the warm regime (after the regime shift of 1988-1989), the warm tempera-ture from the surface to the subsurface in association with the weakened Asian monsoon and positive AOI enhanced the production of zooplankton biomass and, therefore, the biomass of warm water pelagic fishes (e.g., Pacific saury, anchovy jack mackerel, common squid) increased. Quasi-decadal alternations in dominant species known as species replacement have been evident in association with the climate regime shifts over the last century.
Climate variation governed the alternating periods of the A group (Pacific sardine), B group (chub mackerel), and C group (Pacific saury, anchovy, jack mackerel). The alternation sequence was as follows: the Pacific herring collapse (1920s), followed by the A group (cool regime in the 1930s), the C group (in the 1950s-early 1960s), the B group (late 1960s to early 1970s), and then A group again (cool regime in the late 1970s to the early 1980s). As Pa-cific sardine (A group) biomass decreased in association with the climate regime shift (1988-1989), the abundance of the C group increased during the warm regime in the 1990s-2000s.
The periods appear to correspond to different periods of winter wind related to the Monsoon modes as out-lined in the following 2 scenarios. In the first scenario, the group A (Pacific sardine) period, cold temperatures with negative SST anomalies in the Far East prevailed over to the +MOI with increased northwesterly winds, which are mainly responsible for the depth of the wind mixed layer and enhanced phytoplankton production in the spring (e.g., 1980s). This finding suggests that the collapse of the sardine population was caused not by over-fishing but by drastic recruitment failure in the 4 years after 1988, which in turn were attributed to environmental regime shifts (Watanabe et al. 1995, Ebisawa and Kinoshita 1998, Wada and Jacobson 1998, Noto and Yasuda 1999). During the deep winter mixed-layer years (e.g., 1980s), a phyto-plankton bloom occurs in spring; in contrast, during the shallow winter mixed-layer years (e.g., 1990s), the bloom occurs in winter in the Kuroshio Extension region (Ni-shikawa and Yasuda 2008). It was suggested that the de-cline in the sardine population during the late 1980s to early 1990s was due to an insufficient spring food supply in the KE region in which sardine larvae and juveniles are transported.
In the second scenario, the group C (anchovy, jack mackerel, and Pacific saury) period, warm temperatures with positive SST anomalies prevailed over the -MOI with reduced northwesterly winds that are responsible for the increased zooplankton biomass (e.g., 1990s). The group B (chub mackerel) period preceded the group A period. Therefore, the chub mackerel population (group B) is be-lieved to be a successor of the C group (anchovy, Pacific saury, jack mackerel, and common squid) (Fig.5 ) (Mat-suda et al. 1992, Kuznetsov 1995, Yatsu et al. 2003).