Biomass expansion factors, which convert timber volume (or dry weight) to biomass, are used for estimating the forest biomass and accounting for the carbon budget at a regional or national scale. We estimated the biomass conversion and expansion factors (BCEF), biomass expansion factors (BEF), root to shoot ratio (R), and ecosystem biomass expansion factor (EBEF) for Quercus mongolica Fisch. and Quercus variabilis Bl. forests based on publications in Korea. The mean BCEF, BEF, and R for Q. mongolica was 1.0383 Mg/m3 (N = 27; standard deviation [SD], 0.5515), 1.3572 (N = 27; SD, 0.1355), and 0.2017 (N = 32; SD, 0.0447), respectively. The mean BCEF, BEF, and R for Q. variabilis was 0.7164 Mg/m3 (N = 17; SD, 0.3232), 1.2464 (N = 17; SD, 0.0823), and 0.1660 (N = 8; SD, 0.0632), respectively. The mean EBEF, as a simple method for estimating the ground vegetation biomass, was 1.0216 (N = 7; SD, 0.0232) for Q. mongolica forest ecosystems, and 1.0496 (N =; SD, 0.0725) for Q. variabilis forest ecosystems. The biomass expansion factor values in this study may be better estimates of forest biomass in Q. mongolica or Q. variabilis forests of Korea compared with the default values given by the Intergovernmental Panel on Climate Change (IPCC).
The magnitude of forest biomass plays a key role influencing the global carbon cycle and assisting in meeting greenhouse gas emission targets (Ciais et al. 2008, Li et al. 2010b, Nabuurs et al. 2010). With a large number of statistically valid plots, forest inventories have been recognized as appropriate data for identifying the size and spatial patterns of forest biomass at a regional or national scale (Schroeder et al. 1997, Choi et al. 2002, Son et al. 2007b, Guo et al. 2010, Li et al. 2010b). Most forest inventories only record the merchantable timber volume and exclude non-commercial components, such as branches, foliage, and twigs (Fang and Wang 2001). Biomass expansion factors (BEF) that convert, expand, or reduce the volume (or dry weight) of the biomass estimate are normally used for predicting tree biomass as a function of stand structural variables to estimate forest biomass and account for the non-commercial components (Fang et al. 2001, Lehtonen et al. 2004, Somogyi et al. 2007, Son et al. 2007b).
BEF have been introduced in recent decades to estimate forest biomass on a landscape or regional level (Fang et al. 1998, Van Camp et al. 2004, Somogyi et al. 2007). For example, BEF were obtained by compiling data from the literature to estimate country-level biomass (Fang et al. 1998, Van Camp et al. 2004). Reliable estimation and representativeness of BEF are crucial for accurately estimating changes in forest biomass and carbon stock. BEF have also been strongly recommended by the Intergovernmental Panel on Climate Change (IPCC) guidelines (IPCC 2003, 2006). The IPCC (2006) revised the volume-based BEF and weight-based BEF into the biomass conversion and expansion factors (BCEF) and BEF, respectively.
This study estimated the biomass expansion factors (BCEF, BEF, R, and ecosystem biomass expansion factor [EBEF]) for
Studies on biomass were reviewed in the literature by tree species in Korea, and biomass data sets reported for
>
Biomass conversion and expansion
The general characteristics of
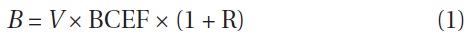
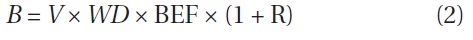
where
In addition, EBEF, the ratio of forest ecosystem biomass (total tree biomass plus ground vegetation biomass) to total tree biomass, is used to estimate ground vegetation biomass of
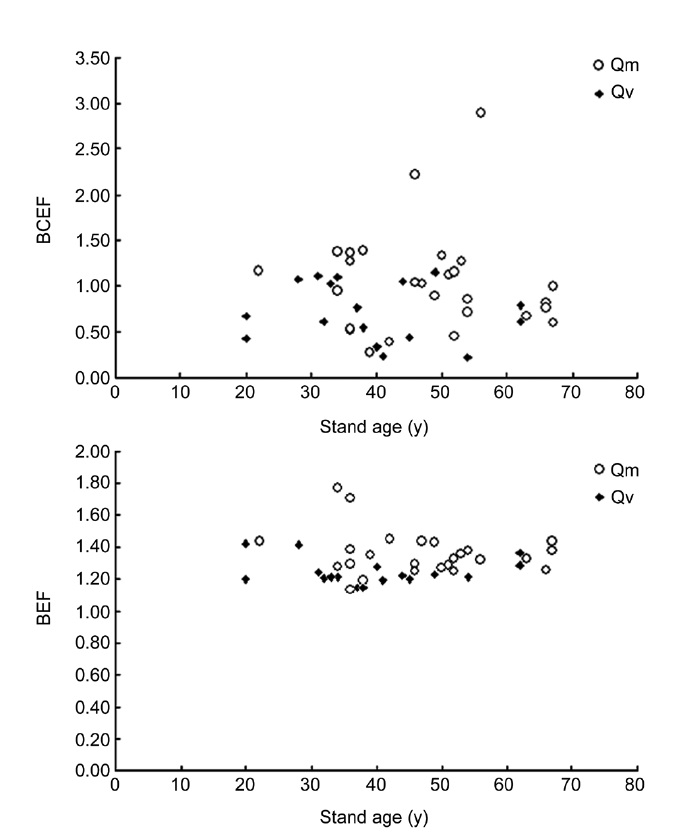
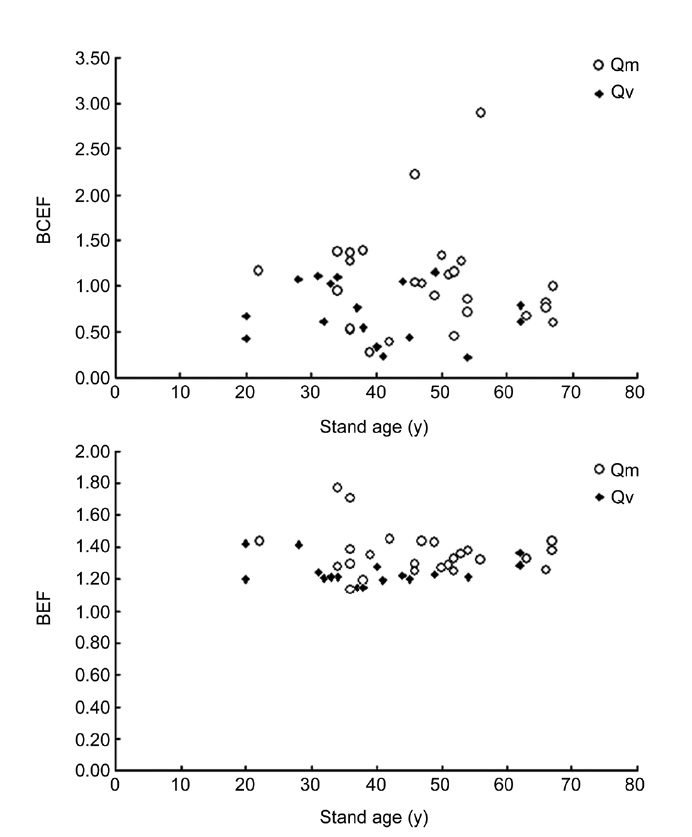
The mean BCEF, BEF, R, and EBEF for
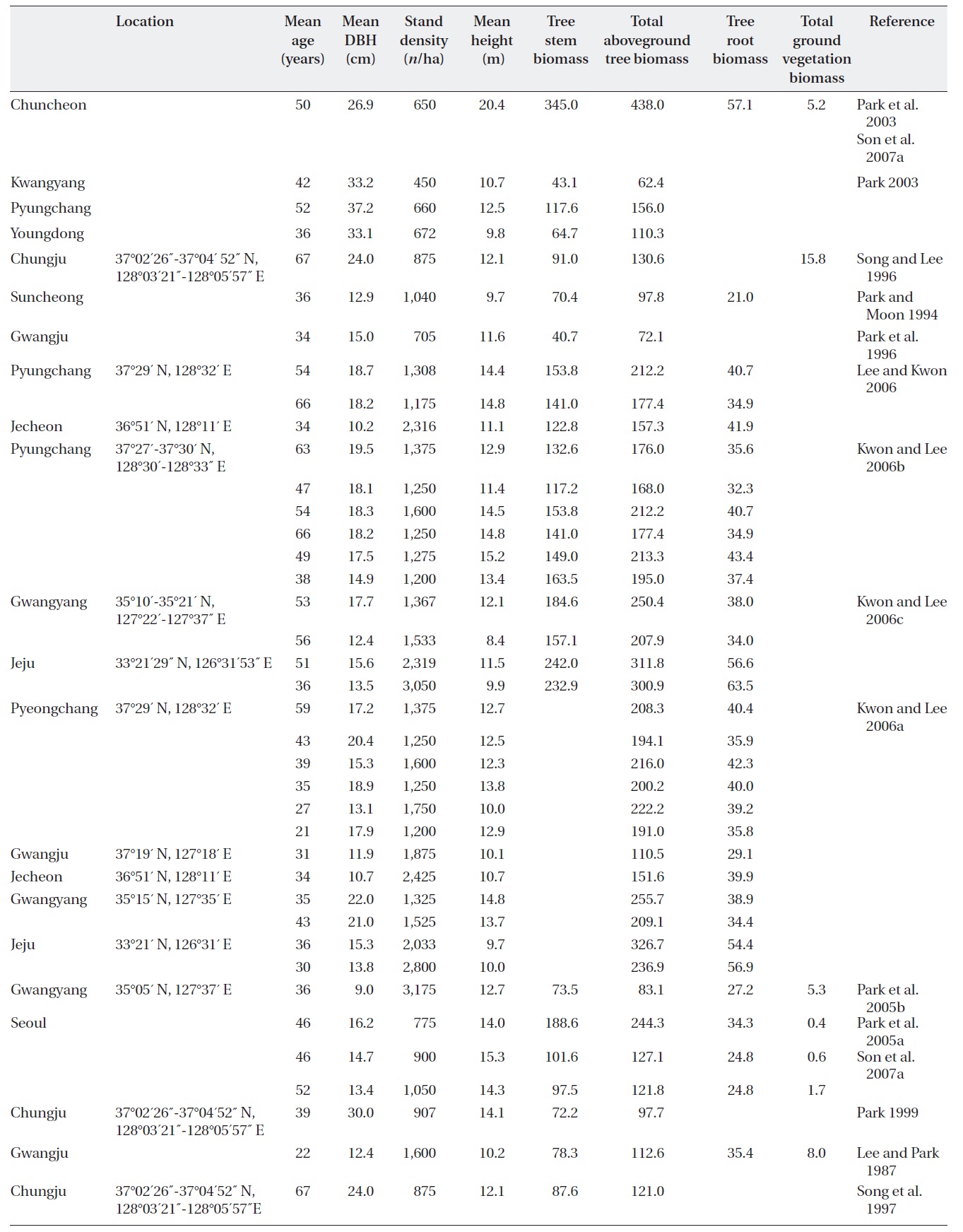
[Table 1.] Stand characteristics and biomass (Mg/ha) of Quercus mongolica forests in Korea.
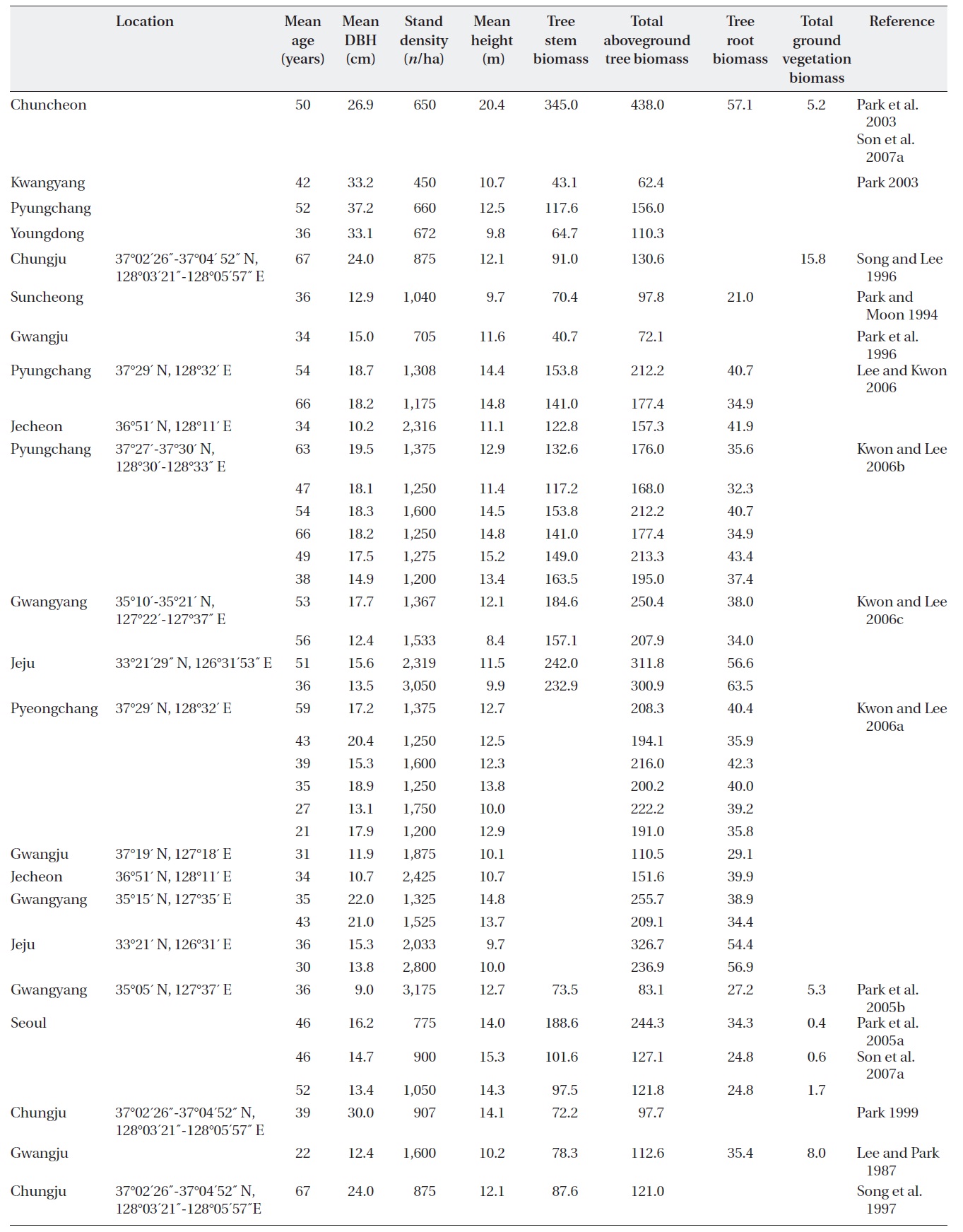
Stand characteristics and biomass (Mg/ha) of Quercus mongolica forests in Korea.
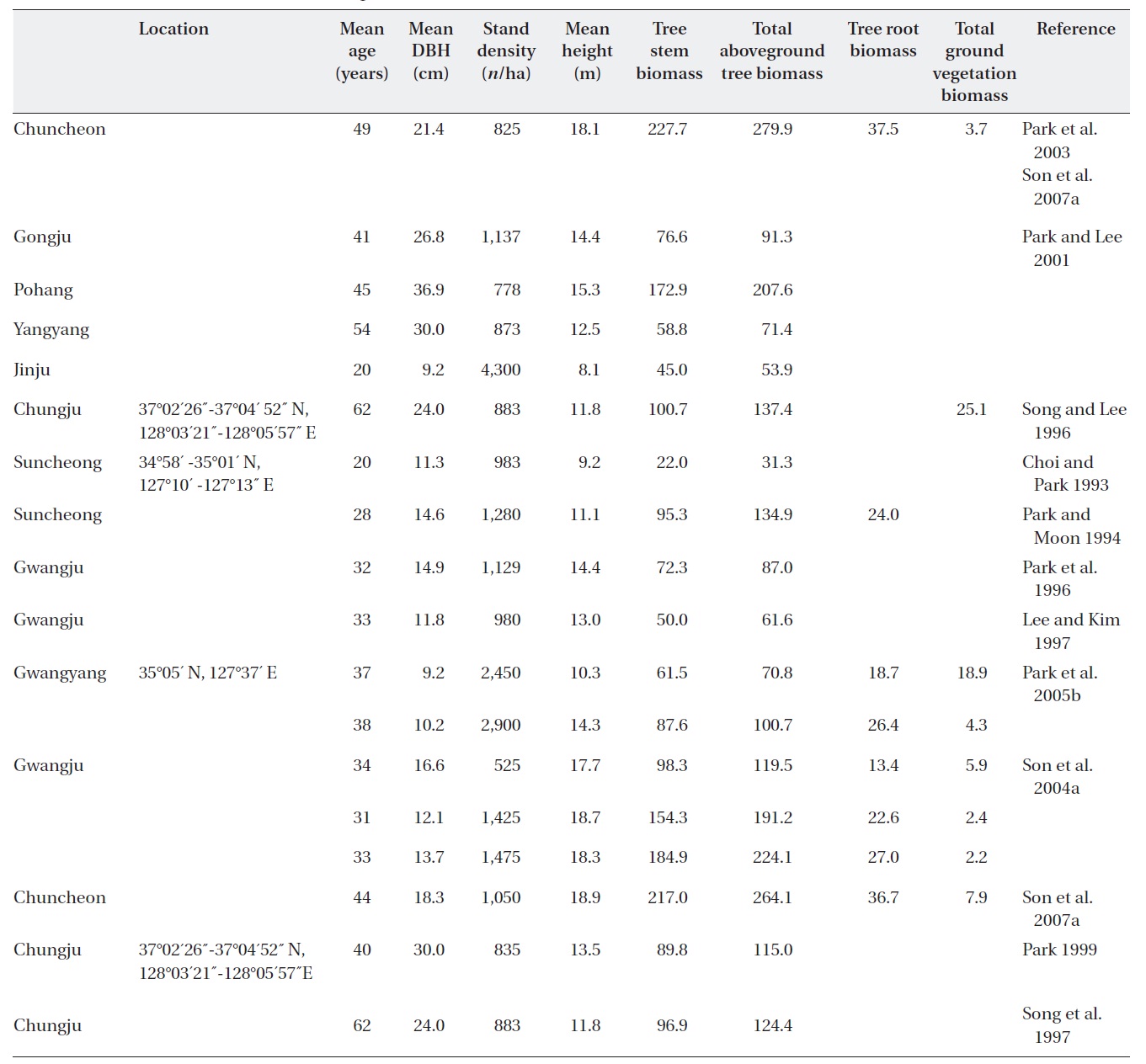
[Table 2.] Stand characteristics and biomass (Mg/ha) of Quercus variabilis forests in Korea.
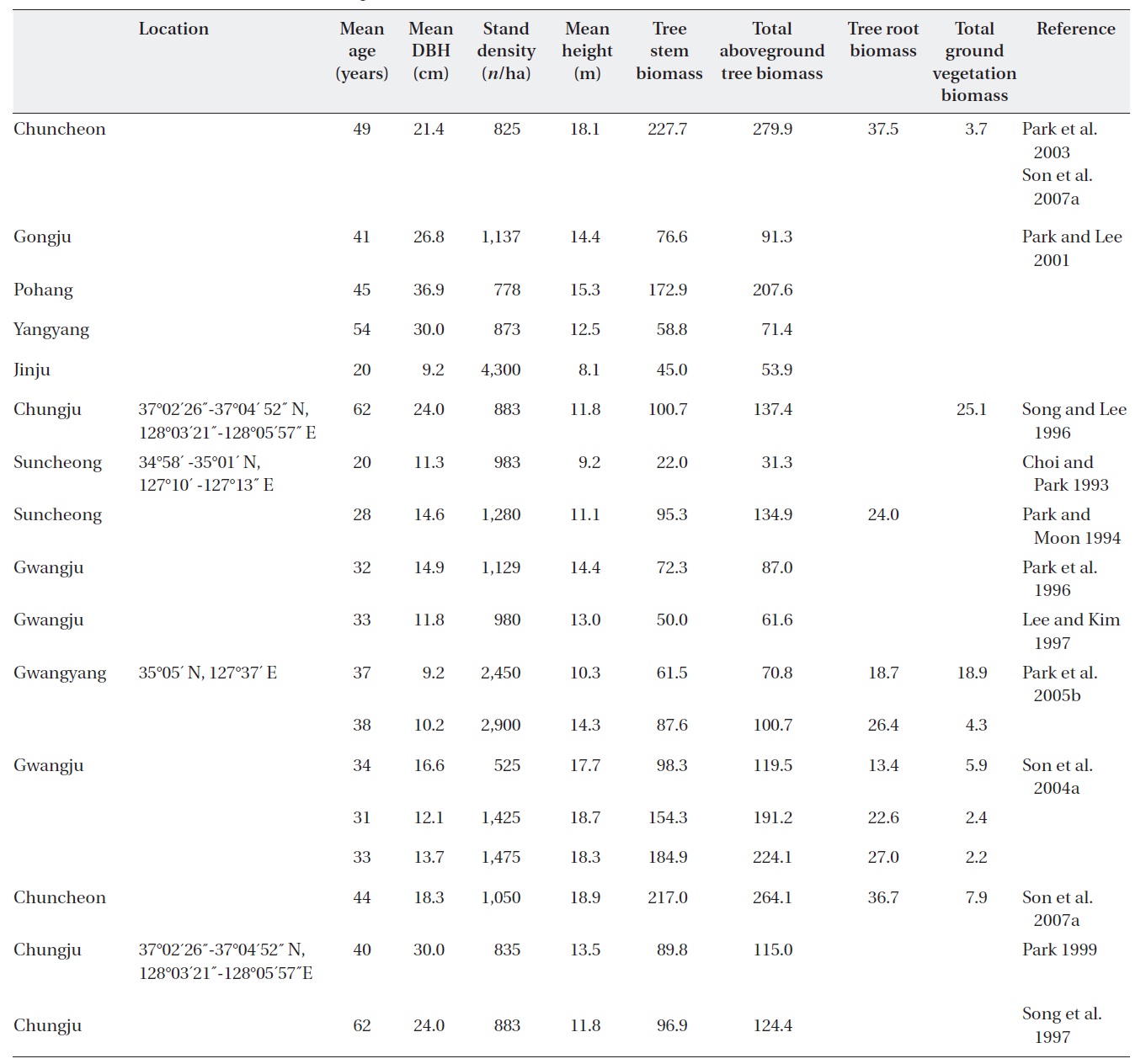
Stand characteristics and biomass (Mg/ha) of Quercus variabilis forests in Korea.
and 1.02 Mg/m3 for oak species (
The IPCC (2003) provides tables of the default values for the BEF of temperate broadleaf forests and the R of temperate oak forests. The IPCC (2006) also provides default values for the BCEF of temperate hardwoods forest. For a better understanding of applicability of the biomass expansion factors in the Korean case, the mean BCEF, BEF and R values for
using the default values.
Recent studies indicate that the relationships between BEF and stand age are heteroscedastic and non-linear. For example, Li et al. (2010c) reported that BEF is expressed as a logarithmic equation of stand age for natural Japanese red pine (
Ground vegetation is an important biomass component in oak forest ecosystems. The EBEF was easily defined, and it is believed that the EBEF is a complementary method for estimating the ground vegetation biomass in