Marine gastrotrich fauna of East Asia is still poor, in spite of serial taxonomic studies accomplished during the past 10 years and earlier. Twenty eight species of 11 genera have been recorded in five families: Cephalodasyidae, Lepidoda-syidae, Thaumastodermatidae, Chaetonotidae, and Xenotri-chulide. In Japan, 10 species of seven genera (Saito, 1937; Sudzuki, 1976, 1979; Chang et al., 2002; Lee and Chang, 2011); in Korea, 17 species of six genera (Chang et al., 1998a, 1998b; Chang and Lee, 2001; Lee and Chang, 2002, 2003, 2004, 2006, 2007; Lee et al., 2009); and in the Russian Far East, only one species,
Herein, we describe a new gastrotrich species of the genus
Gastrotrichs were collected from the shell-sandy bottom at about 6-7m in depth around Saeseom islet, which is located 150 m off Seogwipo Harbor at the southern coast of Jeju Is-land, South Korea.
General sampling methods and extraction process are same with those of Lee et al. (2009), because the specimens were obtained from the same locality with the same collection data as for
Type specimens have been deposited in the National Insti-tute of Biological Resources (NIBR), Incheon, Korea and also in the authors' collection at the specimen room of the Department of Biology, Daegu University (DB).
Terminology mostly follows Ruppert (1991) and Clausen (2000). Abbreviations used include: fpo, female gonopore; Lt, total length, from anterior tip of head to posterior tip of pedicles including adhesive tubes; mpo, male gonopore; Ph-JIn, junction between pharynx and intestine; TbA, anterior adhesive tubes; TbD, dorsal adhesive tubes; TbDL, dorsola-teral adhesive tubes; TbP, posterior adhesive tubes; TbV, ven-tral adhesive tubes; TbVL, ventrolateral adhesive tubes; U, percentage unit of Lt, used for the location (U-) from anterior to posterior, or for the relative length (-U).
Order Macrodayida Remane, 1925 [Rao and Clausen, 1970]
1*Family Planodasyidae Rao and Clausen, 1970
2*Genus Crasiella Clausen, 1968
3*Crasiella clauseni n. sp. (Fig. 1)
Material examined. Holotype (DBG1601), 14 paratypes (NIBRIV0000245017, 0000245018, DBG1602-13), mounted in glycerin, Korea, Jeju Island, Seogwipo, Saeseom islet, 33 ? 14'13?N, 126 ? 33' 33?E, 24 Apr 2006, leg, Lee J, Chang CY, Kim SH, in sublittoral sandy sediments containing shell crumbles, 6-7m deep.
Etymology. The proposed specific name is in commemora-tion of the late Dr. Claus Clausen who established the genus
Diagnosis.
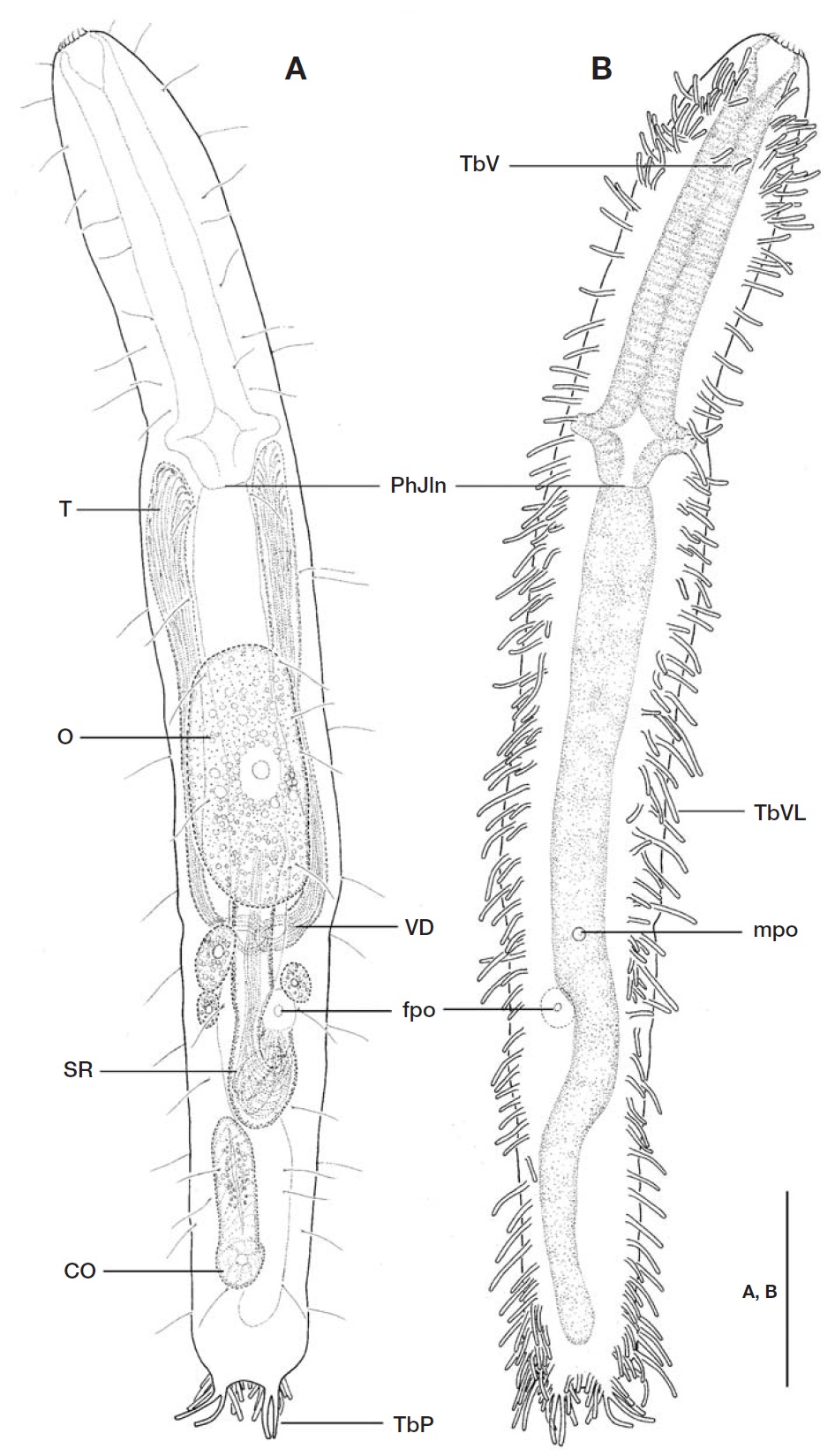
Description. Holotype: Body (Figs. 1, 2) elongate, oblong, dorso-ventrally flattened, with small bilobed caudum, Lt 385㎛ long. Lateral sides of body nearly parallel, weakly con-stricted in the level of posterior pharyngeal region and a slightly narrowed distal head region. Head indistinct; lacking shallow lateral pits and ciliary tufts. Widths of anterior part of head/PhJIn/trunk/base of caudal lobes 28/41/42/17 ㎛ at U05/U33/U58/U96, respectively.
Dorsal surface somewhat uneven, without cuticular arma-ture like hooks, scales and plates.
Numerous hairs (ca. 12-18 ㎛ long) irregularly arrayed in 2-3 columns per side on subdorsal and lateral surfaces along nearly whole body length (U09-U90); several sensory hairs(ca. 5 ㎛ long) implanted sporadically on anteroventral sur-face at U01-U03.
Locomotor ciliation covering entirely ventral surface from rear of oral opening to base of caudum.
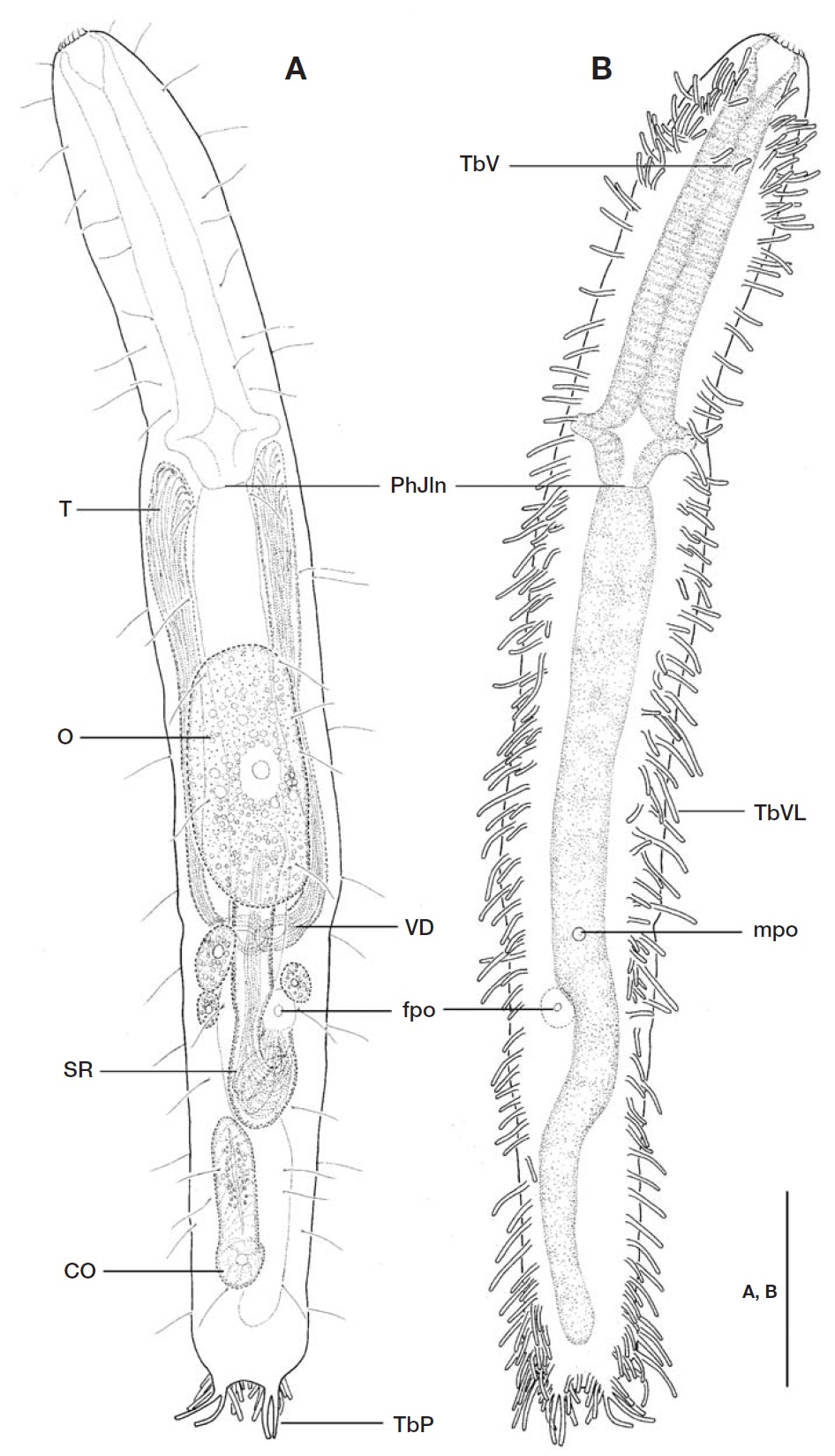
Adhesive tubes: TbA and TbVL not obviously separated, 119-123 tubes per side, somewhat slender, with different sizes(ranging from 5 ㎛ to 19 ㎛ long), extending over nearly whole body length (U3-U95) from rear of oral opening to base of caudal lobes. Adhesive tubes in pharyngeal region representing different arrangements of horizontal and longi-tudinal rows; foremost 2 pairs located rather ventrally at U03 and U04, respectively; 6 horizontal rows per side situating in anterior pharyngeal region from U05 to U13, each row composed of 2-6 tubes (5, 6, 5, 4, 2, 3 tubes in left side and 4, 4, 4, 3, 2, 2 in right, respectively) in different sizes (ca. 6-11 ㎛ long); followed by 14-16 tubes per side, positioned at irregular intervals, forming a longitudinal row in remaining pharyngeal region at U15-33. About 82-85 adhesive tubes per side, ca. 5-19 ㎛ long, arranged irregularly between PhJIn to front of caudal lobes, crowded near posterior end of trunk at level of anus (U94). A pair of TbV (ca. 7 ㎛ long), not foot-type, implanted mid-ventrally in anterior pharyngeal re-gion at U10. TbD (L) absent. TbP comprising 9-10 tubes per side, forming a small caudal lobe, consisting of 2 long distal tubes (15 ㎛ long), 3 lateral (ranging ca. 7-10 ㎛ long) and 4-5 medial ones (ranging ca. 5-8 ㎛ long); lateral and medial TbP gradually decreased in size towards base of caudal lobe.
Digestive tract: Oral opening ornamented with corona of 15-18 short hook-like projections (ca. 2-3 ㎛ long) along its terminal edge; buccal cavity funnel-shaped and slightly cuti-cularized; pharynx 127 ㎛ long, with a pair of pharyngeal pores, opened laterally at U30; intestine broad and straight in its anterior region, then narrowing gradually towards anus,and curved near posterior half of seminal receptacle; anus situated ventrally at U92.
Reproductive system: Testes paired, situated along anteri-or intestinal region, their tips reaching just behind pharyngeal knobs at U30; vasa deferentia including spermatozoa, coiled and joined at the middle of intestine region at U63-65; mpo emerging from joint of vasa deferentia at U64. Ovum well-developed,ca. 31×67 ㎛, situated dorsally in mid-intestinal region at U44-U62, followed by 3 small oocytes. Seminal receptacle elongate, 77 ㎛ long, tube-shaped with its poste-rior part a little swollen and folded; posterior end folded, opened (fpo) ventrally on two-thirds of seminal receptacle at U70; situated dorsally from posterior part of ovum to anterior
end of copulatory organ at U57-U78. Copulatory organ (44㎛ long) tube-shaped with an internal duct, gradually nar-rowing to its anterior edge, located at U77-U89.
Variability. Body lengths of 12 adult specimens ranged 366 to 440 ㎛ (mean 396 ㎛, standard deviation 24), and maxi-mum width 41-58 ㎛, measured in glycerin mount.
The number of adhesive tube showed somewhat asymmet-rical array and individual variability. Especially, TbVL in the intestinal region ranged from 52 to 83 tubes. In contrast with ventrolateral tubes, the paired TbV was rather consistent, located typically in the anterior pharyngeal region, except for one specimen lacking the pair, and for four specimens with only one tube in one side.
Epidermal glands were not clear in holotype, however, in five paratypes numerous epidermal glands were irregularly scattered at dorsal, dorsolateral, and ventrolateral surfaces, along nearly the whole body length.
Three sub-adults were included among 15 specimens exam-ined, with the body sizes of 281 ㎛, 302 ㎛, 325 ㎛ long and 33 ㎛, 42 ㎛, 48 ㎛ wide, respectively. They showed fewer ventrolateral and posterior adhesive tubes than in adults:3-4 horizontal rows (vs. 5-7 in adults) in the anterior-most pharyngeal region; 21-39 ventrolateral tubes in the intestinal region (vs. 52-83 in adults); and 5-6 TbP per side (vs. 8-11 in adults).
Taxonomic affinities. To date, six species have been record-ed in the genus
Among them,
Other three congeneric species,
A key to the species of genus Crasiella Clausen
1. With pestle organ in the cephalic region ······································································
2. TbA and TbVL separated ··········································3 TbA and TbVL not separated ··················································································
3. TbA forming 2 longitudinal rows ···············································································
4. With 5-7 horizontal rows of adhesive tubes and a pair of TbV in the pharyngeal region ··············································································
5. TbA forming 1 diagonal row ··················································································
Korean name: 1*납작털배벌레과, 2*어리납작털배벌레속, 3*어리납작털배벌레