Ciliates have been recognized as important components in the marine microbial food webs and oligotrich ciliates usually occurred dominantly among the planktonic ciliate community(Froneman and Perissinotto, 1996; Quevedo and Anadon,2000). According to Lynn (2008), the oligotrichs were divided into two subclasses; the Oligotrichia (oligotrichs sensu stricto) and the Choreotrichia, while the halteriids were affiliated with the stichotrichs. To date, about 1,490 species of oligotrichs (sensu lato) have been described worldwide but the validity is not confirmed. Of them, about 120 oligotrichids and 80 aloricate choreotrichids (not including tintinnids) have been recorded, only about 60% have been described, or redescribed using protargol staining method (Agatha, 2004a,2004b, 2011; Agatha and Struder-Kypke, 2007). Studies on marine planktonic ciliates have been carried out in Korea since 1980’s, only 4 oligotrichids and 10 aloricate choreotrichids have been reported (Table 1) (Suzuki and Han, 2000;Jeong et al., 2004; Moon et al., 2004; Kim et al., 2005, 2008,2010b; Ma and Choi, 2005; Lee et al., 2006).
The ecological roles of Korean oligotrichs (especially, loricate tintinnids) have been investigated in many studies (Yoo and Park, 2001; Kim and Lee, 2003; Yang and Choi, 2003;Moon et al., 2004; Kim et al., 2007; Lee et al., 2007; Han and Lee, 2010; Lee and Kim, 2010). They are frequently collected and abundant in the surface layer of coastal water and easily identified due to their large size and conserved lorica (Yoo et al., 1988; Yoo and Kim, 1990). Aloricate oligotrichs are more abundant than loricate tintinnids in microzooplankton community (Vaque et al., 1997; Ota and Taniguchi, 2003).However, most ecological studies do not include them because they are not easy to handle due to their limited taxonomic information, fragility and small size of body (Yoo and Park,2001; Kim and Lee, 2003; Lee and Kim, 2010). The taxonomic information of these taxa in Korea is urgently needed to investigate other related studies. Therefore we describe here four aloricate oligotrichs with their abundance including water temperature and salinity.
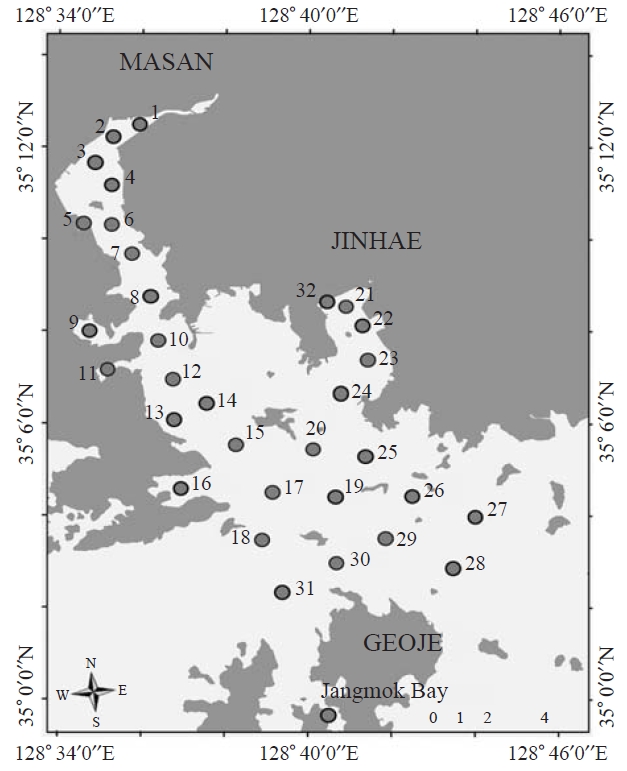
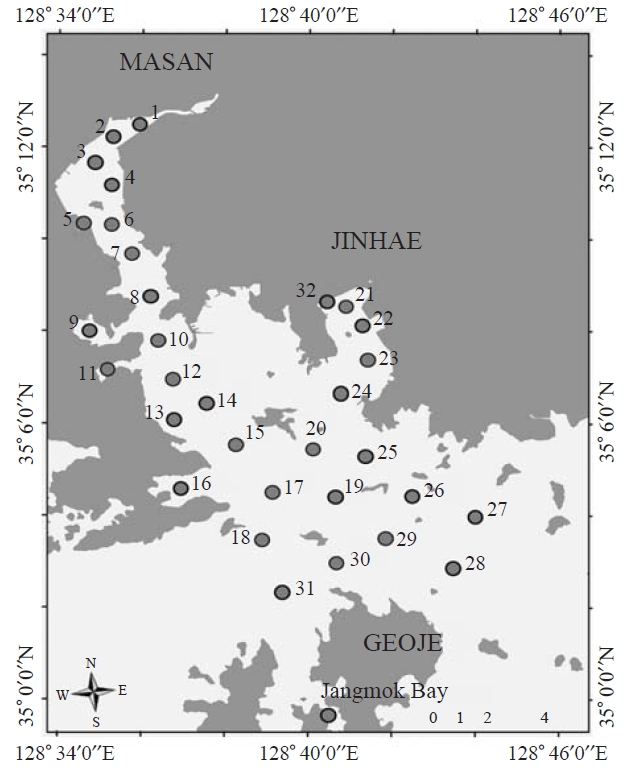
Ciliate samples were seasonally collected from Masan Bay
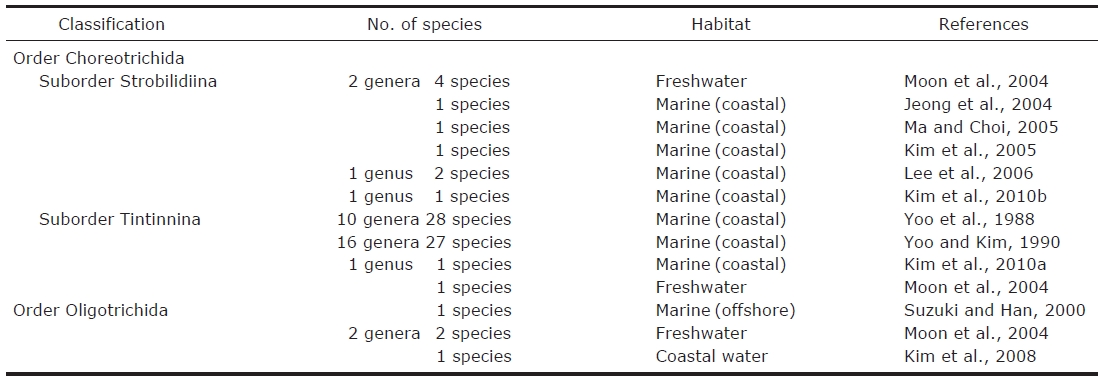
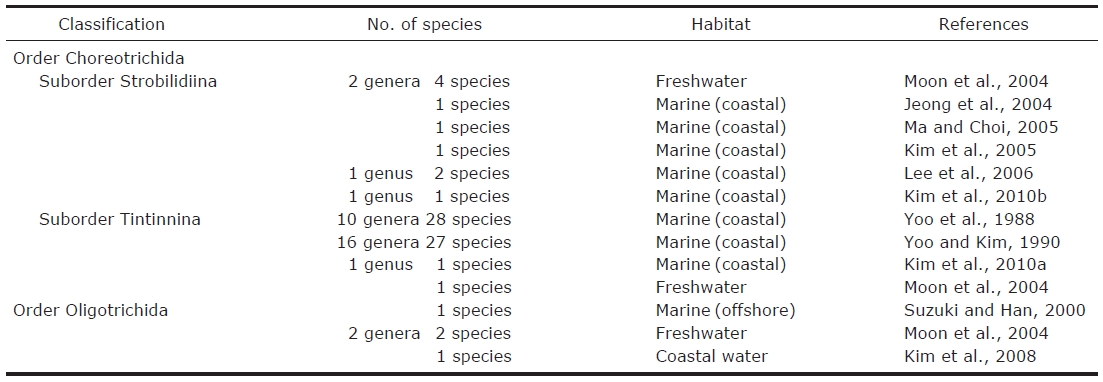
Number habitat and references of oligotrich ciliates (Oligotrichida and Choreotrichida not including tintinnids) described taxonomically in Korea
using Van Dorn sampler at 0.5 m depth of selected sampling sites (32 sites) from April 2006 to November 2008 (February,May, August, and November). In Jangmok Bay, ciliates were sampled by vertical towing from 5m depth to surface using a 20 ㎛ mesh plankton net (30 cm diameter) in June 2009(Fig.1 ). Samples were fixed immediately by Bouin’s solution (at final concentration 10%, v/v) in which most of the ciliates were well preserved. Water temperature and salinity were measured
>
Morphological investigations
In order to observe the morphological characteristics of ciliate cells, the fixed cells were stained and mounted using the quantitative protargol staining and the modified Wilbert’s method (Wilbert, 1975; Montagnes and Lynn, 1987; Foissner et al., 1999). Ciliate cells were identified and counted under ×400-1,000 magnification using a light microscope (Zeiss Axioskop II, Goettingen, Germany).
Terminology and classification for the ciliate taxonomy are according to Montagnes and Lynn (1991) and Agatha (2004a).
Korean name: 1*폐구환소모충목, 2*폐구환무각소모충아목, 3*이종렬소모충과, 4*단이종렬소모충속, 5*원추단이종렬소모충
Class Oligotrichea Butschli, 1889
Subclass Oligotrichia Butschli, 1889
1*Order Choreotrichida Small and Lynn, 1985
2*Suborder Strobilidiina Jankowski, 1980
3*Family Strobilidiidae Kahl in Doflein and Reichenow,1929
4*Genus Rimostrombidium Jankowski, 1978
5*Rimostrombidium conicum (Kahl, 1932)
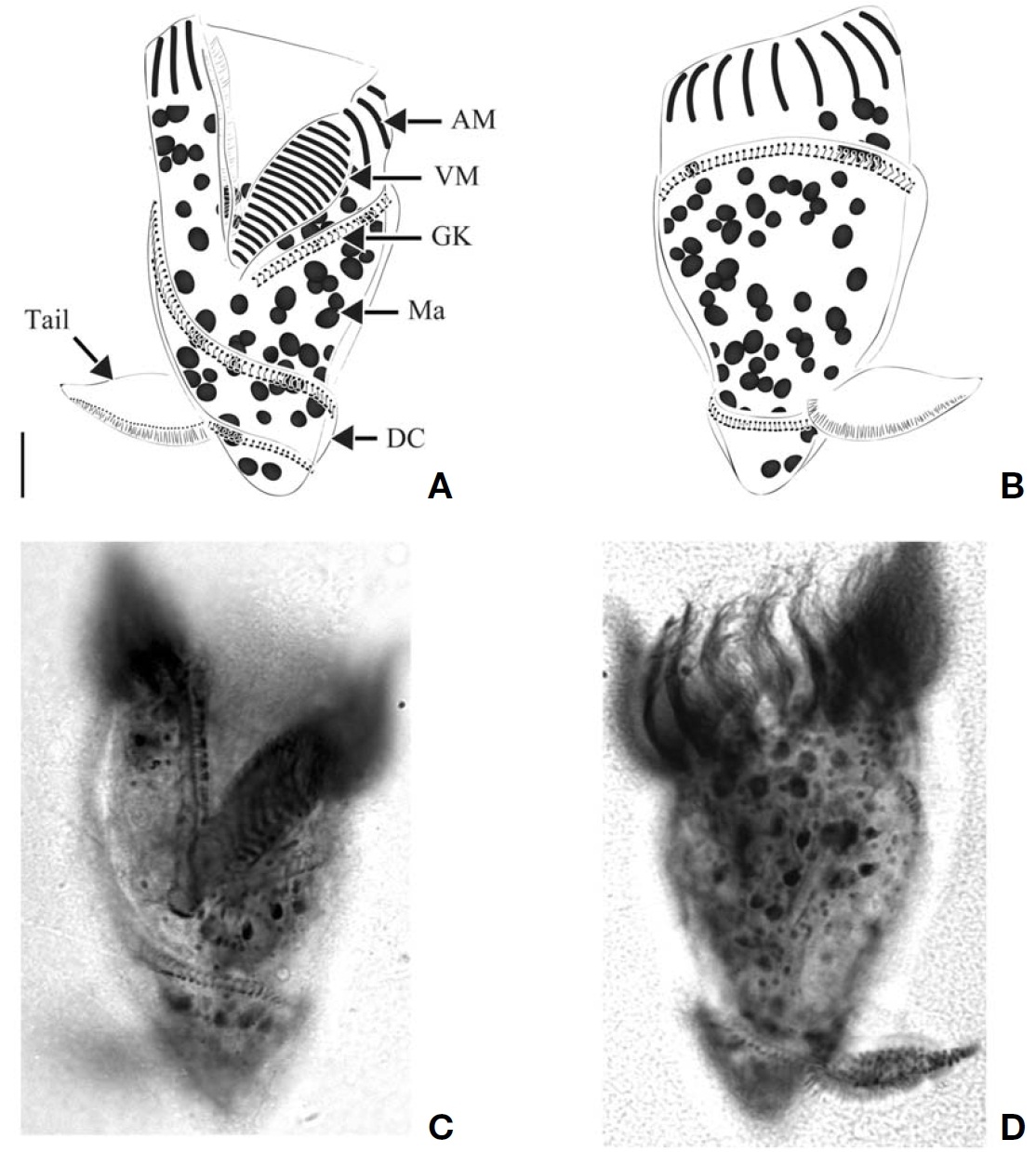
(Table 2,Fig.2 )
Strobilidium conicum Kahl, 1932: 506.
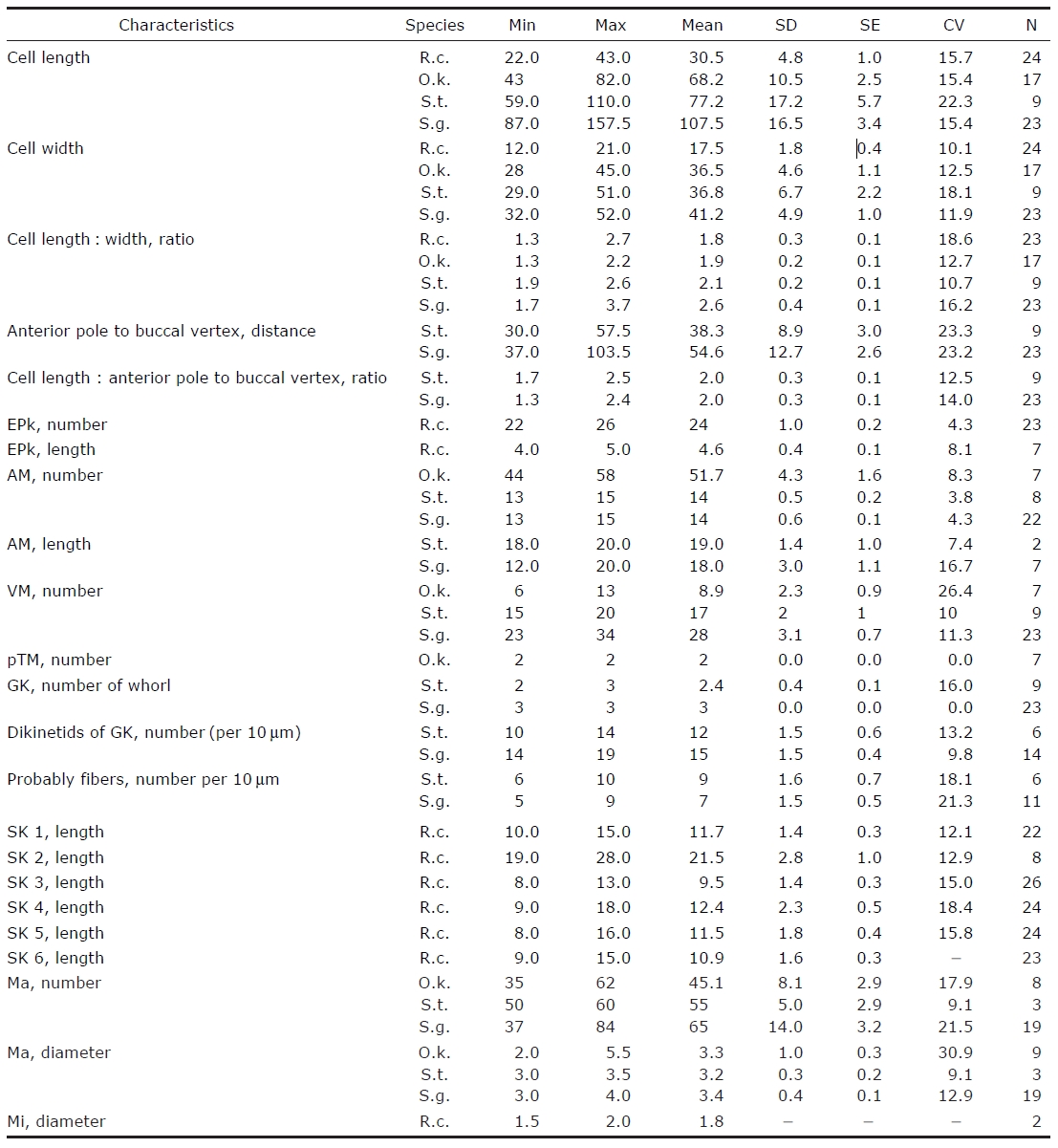
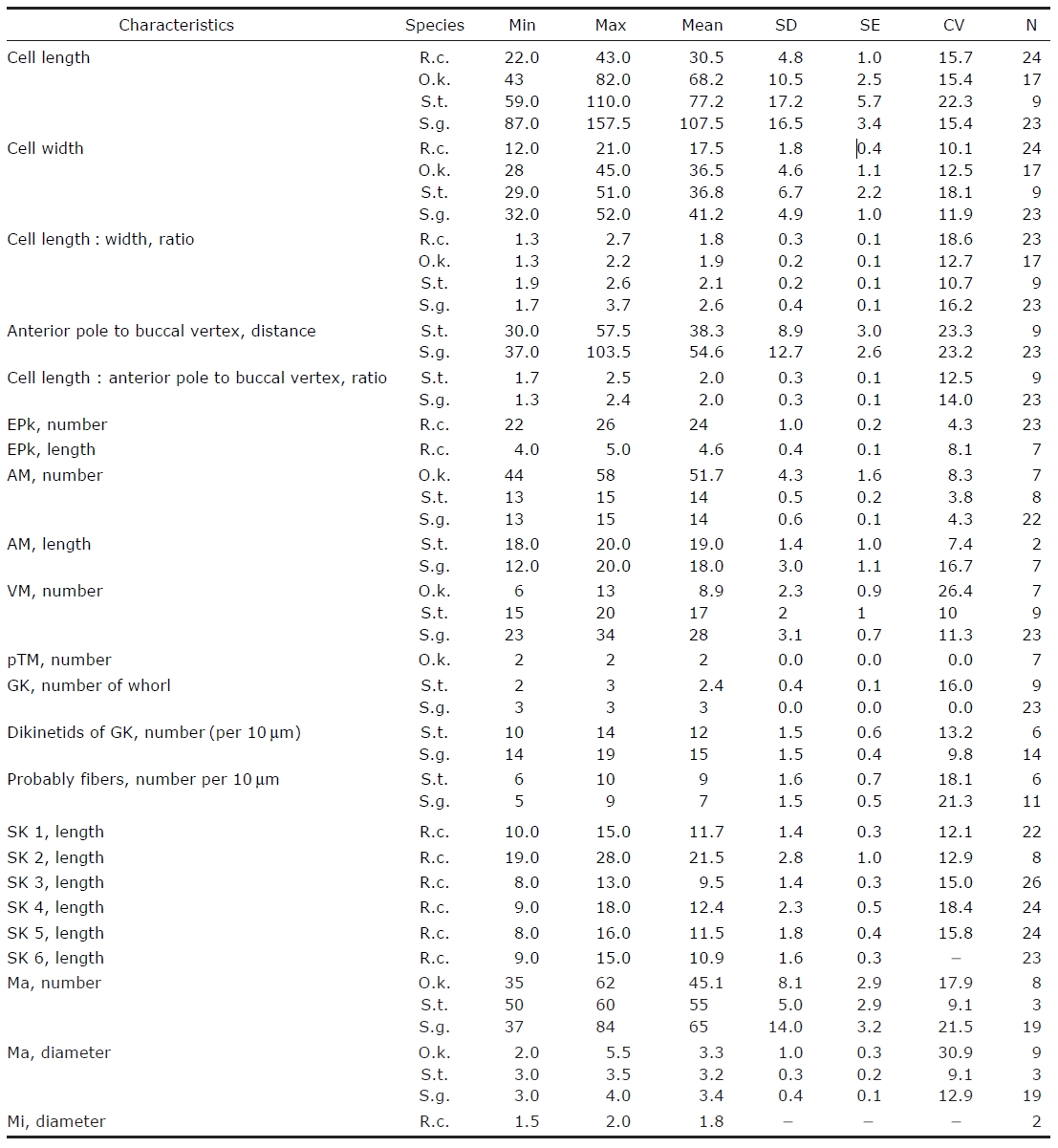
Morphometrical characterization of Rimostrombidium conicum (R.c.) Omegastrombidium kahli (O.k.) Spirotontonia turbinata(S.t.) and Spirotontonia grandis (S.g.)
Rimostrombidium conicum Petz and Foissner, 1992: 160;
Agatha and Riedel-Lorje, 1998: 14.
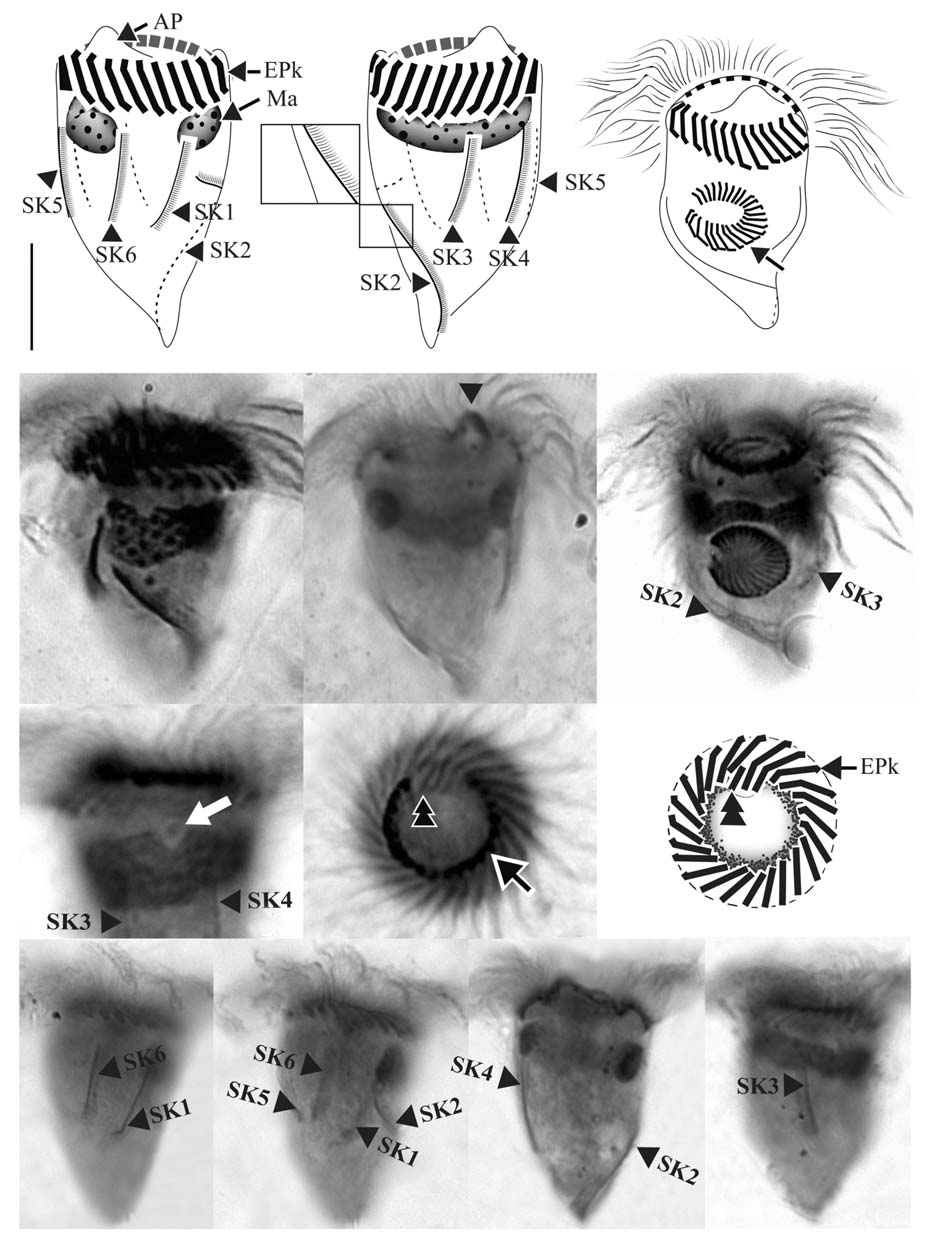
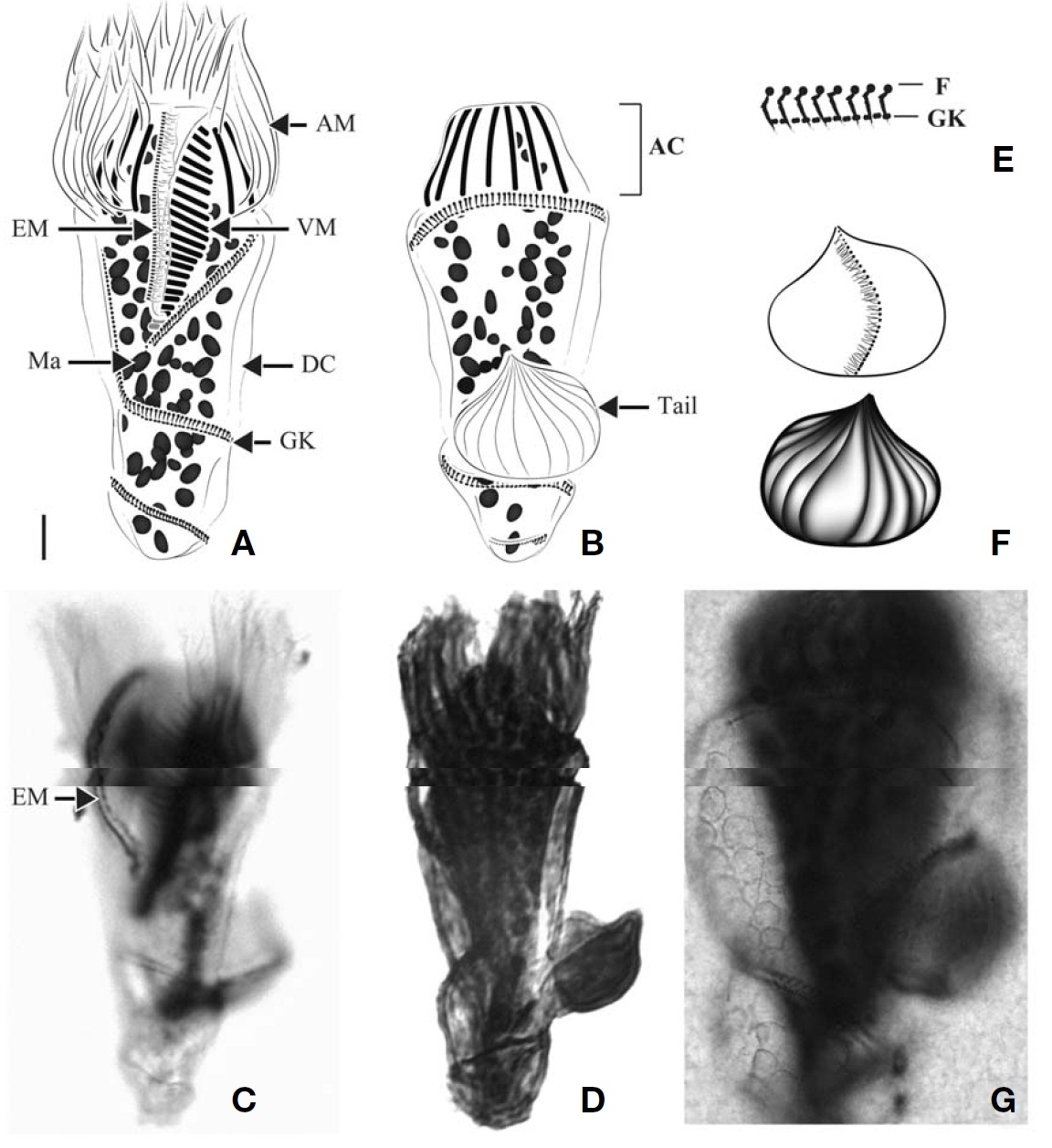
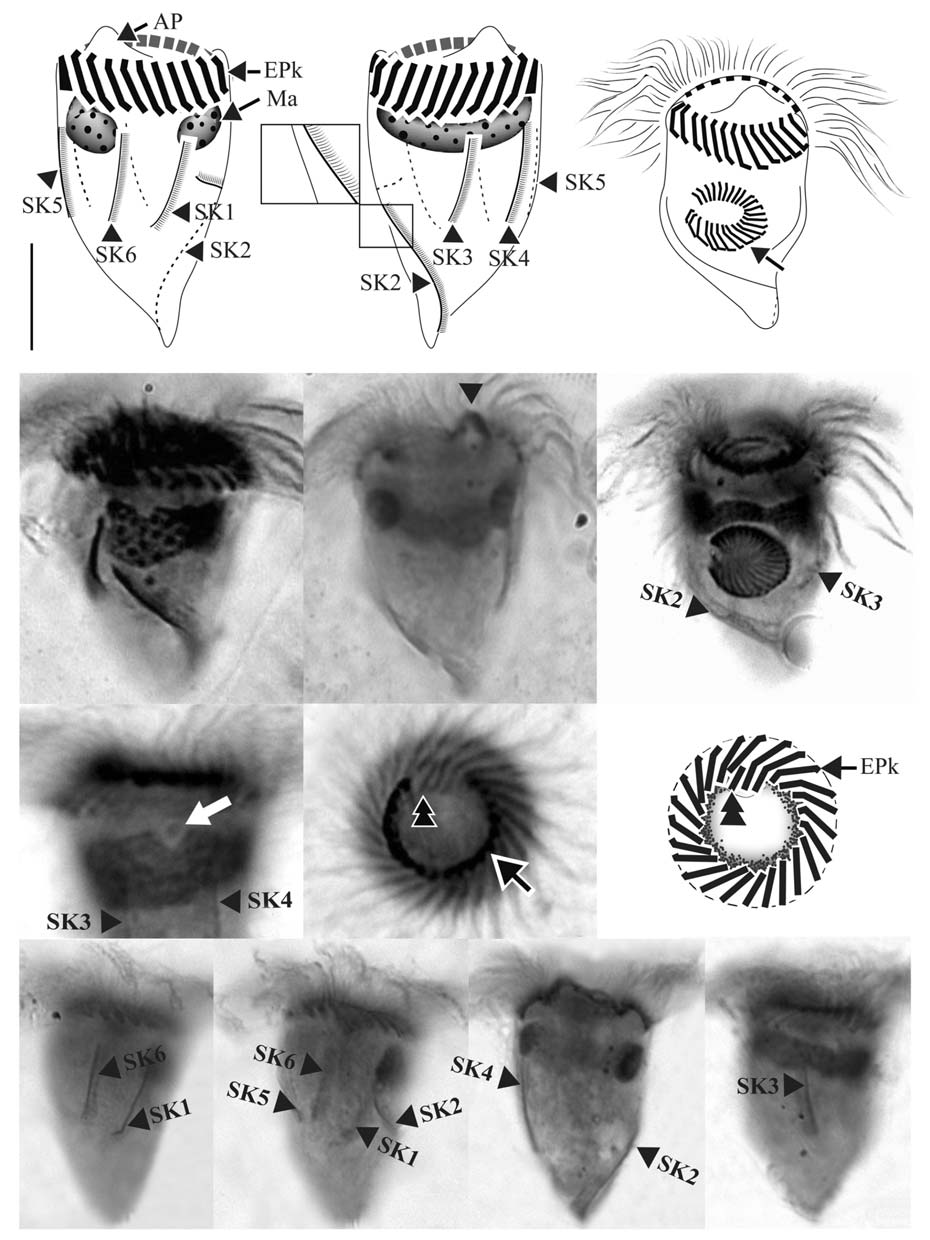
Diagnosis. Body asymmetrically conical shaped; cell size 22-43 μm in length and 12-21 μm in width after protargol preparation;22-26 EPks; one separated IPk; 6 SKs, SK2 longer
than the other kineties, and spirally extending to posterior cell end, SK1, 3, 4, 5, and 6 not extending to posterior end and slightly curved or straight; one Ma C- or horseshoe-shaped and horizontally located underneath EPZ.
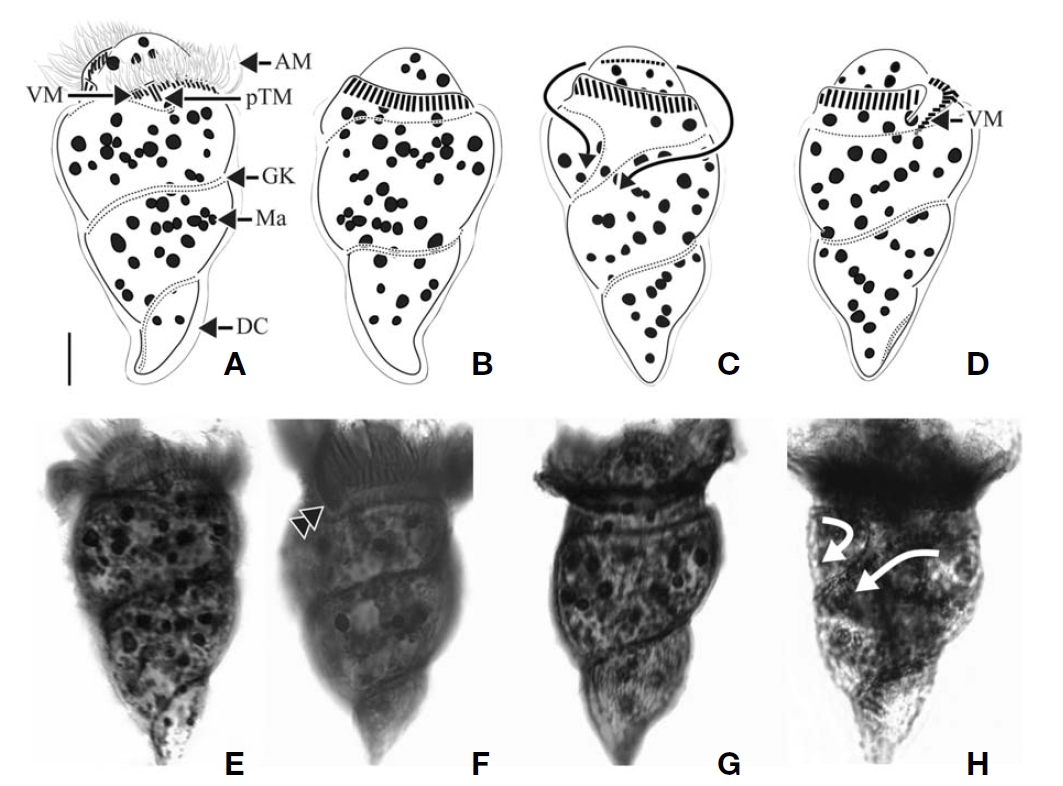
Morphological description of Korean population. Body asymmetrically conical shaped with conspicuous protuberance at anterior pole of cell, bluntly tapering in posterior region.Body size 22-43 μm in length and 12-21 μm in width(average 31×18 μm, n=24) after protargol impregnation(Fig.2 A, B, D, E).
EPZ closed and surrounding anterior end of cell, consisting of 22-26 polykinetids (average 24, n=23). Two elongated EPks extending into eccentric oral cavity. One IPk lying into oral cavity and separated from EPks. A black circle observed below anterior part of EPZ, having gap into oral cavity (“C”shaped position), probably fibres (Fig.2 H, I).
Six SKs composed of short cilia. Except for SK2, SKs not extending to posterior pole of cell and having similar lengths:SK1, 12 μm; SK3, 10 μm; SK4, 12 μm; SK5, 12 μm; and SK6, 11 μm on average. SK2 longer than the other kineties(average 22 μm in length, n=8), slightly sinistrally spiraled and extended to posterior pole of cell. SK1 and SK6 less or not sinistrally curved than SK3-5 slightly dextrally curved or longitudinal (Fig.2 J-M).
Single Ma C- or horseshoe-shaped and contained numerous globular nucleoli, horizontally located underneath EPZ with ventrally opening. One ellipsoidal Mi positioned on dorsal side of Ma (Fig.2 G).
Oral primordium developing between SK2 and SK3
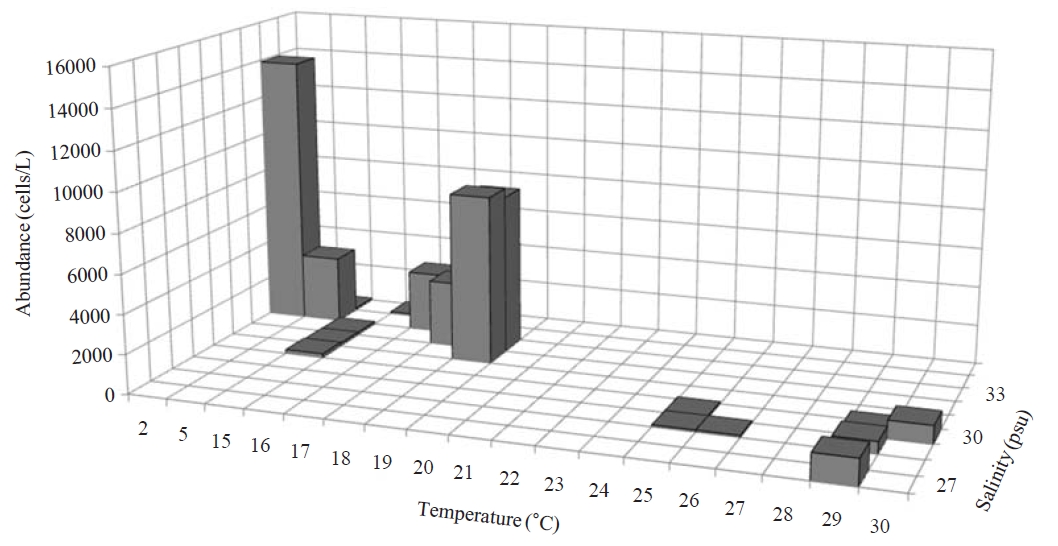
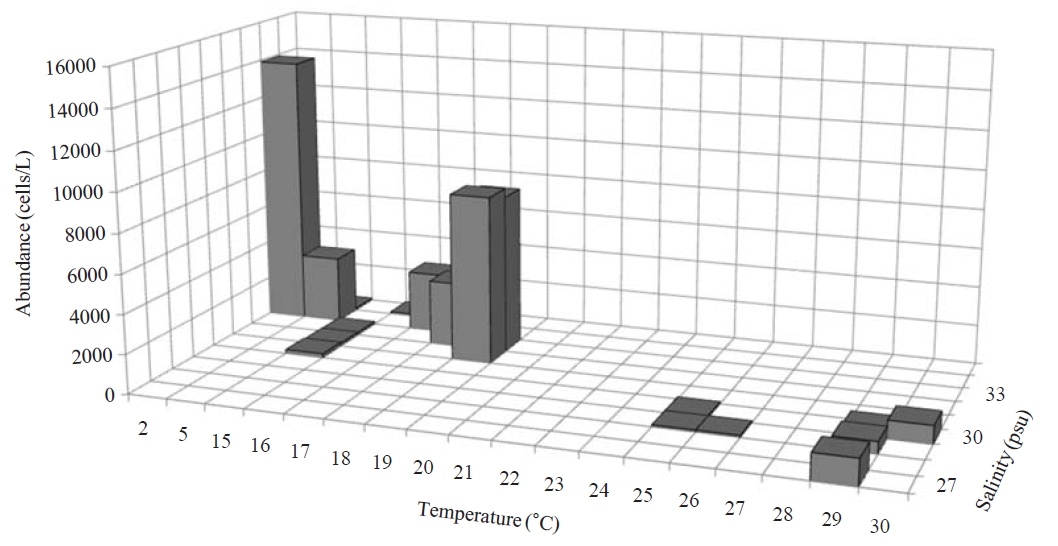
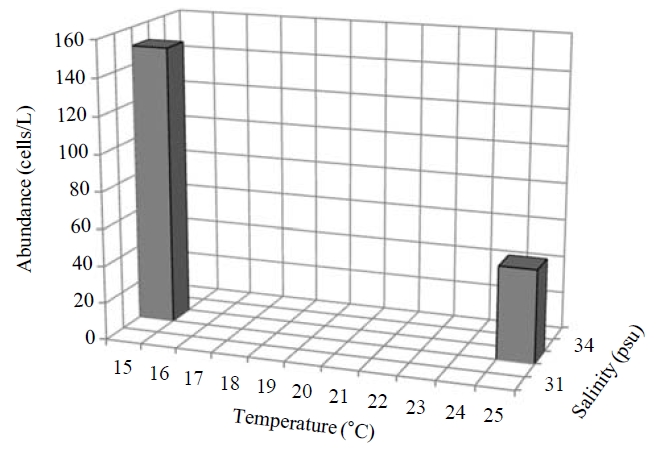
Occurrence and ecological characteristics. Maximum abundance(13,900 cells/L) of this species was observed at M4 site in Masan Bay in February 2008, where water temperature of 2.13℃ and salinity of 34.21 psu were recorded. This species showed highest abundance in cold water (2-5℃) but also occurred at high temperatures (? 15℃) in low abundances(100-8,600 cells/L) (Fig.3 ).
Comparison with other populations. German population was collected from the brackish and coastal waters of the North Sea and with maximum abundance of 100,000 cells/L in August while the maximum abundance of Korean population occurred in February (about 13,900 cells/L) (Agatha and Riedel-Lorje, 1998). The morphology of Korean population is similar to German population (e.g., cell size, shape, number of SKs and EPKs, shape, and location of Ma), except for the arrangement of somatic kineties. SK1 of German population is the longest (vs. SK2 the longest in Korean population).German population has dextrally spiral SK2 which does not reach a posterior pole (vs. SK2 extending to posterior pole in Korean population) (Agatha and Riedel-Lorje, 1998).
1*Order Oligotrichida Butschli, 1889
2*Family Strombidiidae Faure-Fremiet, 1970
3*Genus Omegastrombidium Agatha, 2004
4*Omegastrombidium kahli Song et al., 2009
(Table 2,Fig.4 )
Omegastrombidium kahli Song et al., 2009: 336-337.
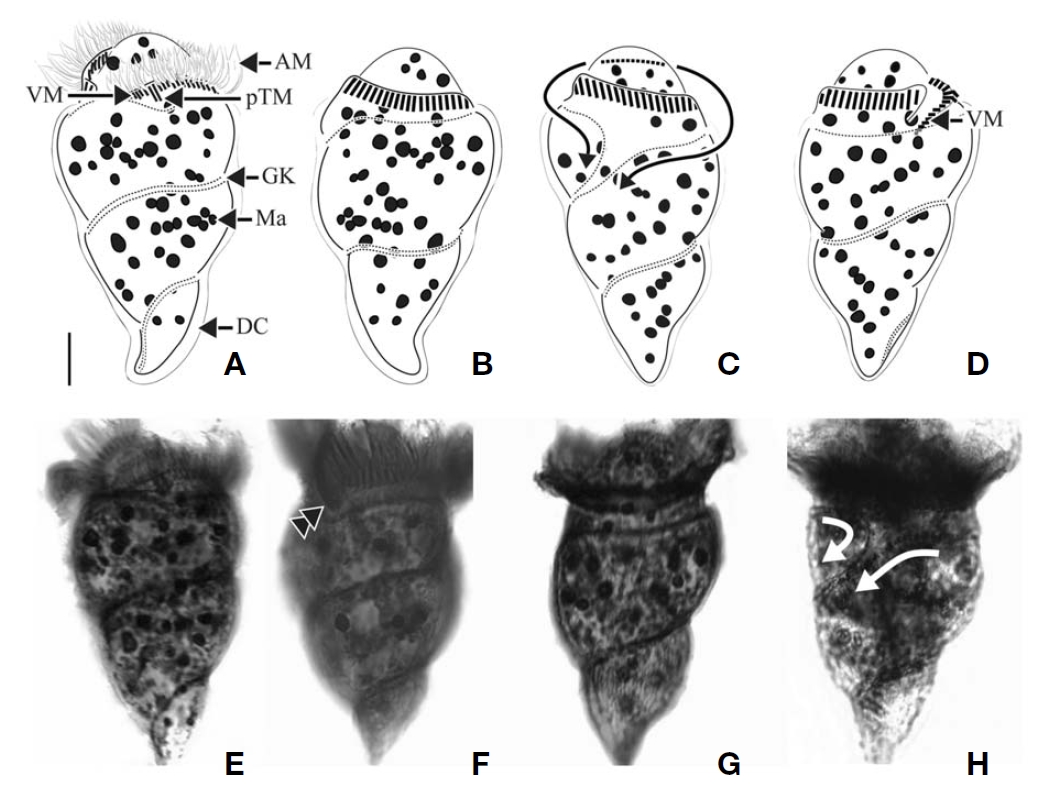
Diagnosis. Body obconical with screw-like appearance; cell size 43-82 ㎛ in length and 28-45 ㎛ in width after protargol preparation; 44-58 AM, 6-13 VM, and 2 pTM; dextrally spiral GK, performing about 2 whorls; 35-62 ellipsoidal Ma,about 2-5.5 ㎛ in diameter.
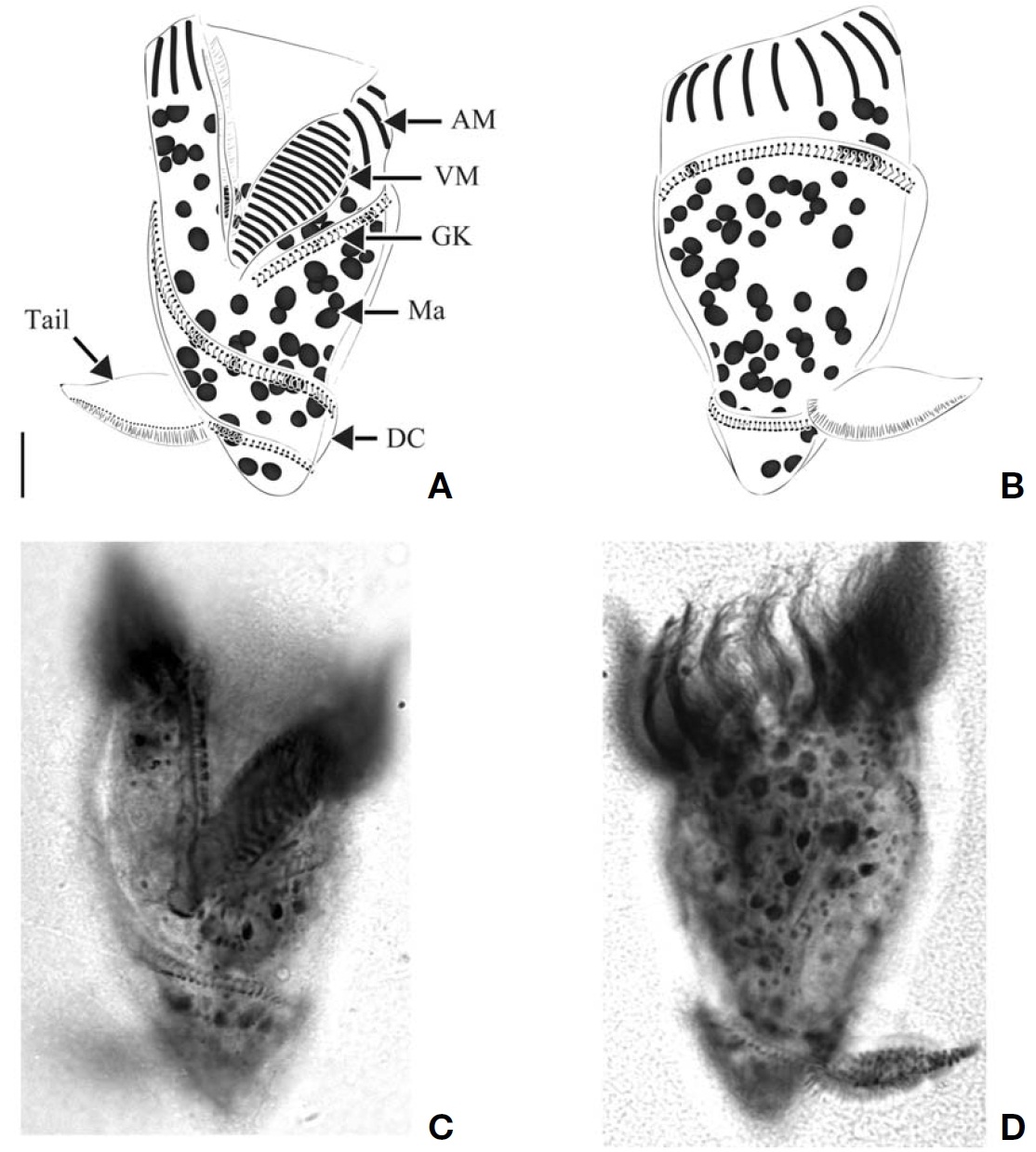
Morphological description of Korean population. Body dextrally screwed cone- or turban shell-shaped with conspicuous apical protrusion (AP) at anterior pole. Cell size 43-82μm in length, and 28-45 μm in width (average 68×37 μm,n=17) after protargol impregnation. Transparent distended cell surface (DC) covered from GK to posterior end of cell(Fig.4 A-D).
Oral polykinetids surrounding anterior pole with ventral opening, divided into AM and VM. Approximate 44-58 AM(average 52, n=7) and 6-13 VM (average 9, n=7). Two pTM positioned between AM and VM and obviously longer than AM and VM (Fig.4 A, F).
GK originated at anterior part of body, both ends facing on left side of anterior part and turning approximately 1.5-2 whorls (with dextrally spiraling) and then terminating at pos-
terior end on right lateral side (Fig.4 C, D). GK on ventral side obliquely extended to right side while GK on dorsal side positioned horizontally (Fig.4 A, B). No ventral kinety found.
About 35-62 (average 45, n=8) ellipsoidal Ma (2-5.5 μm in diameter), scattered throughout cell.
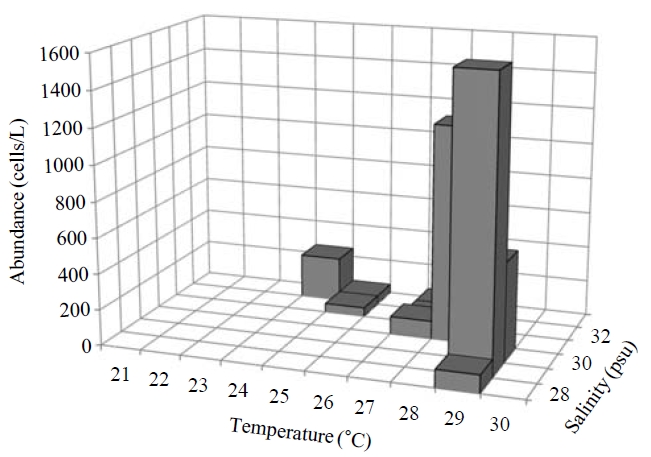
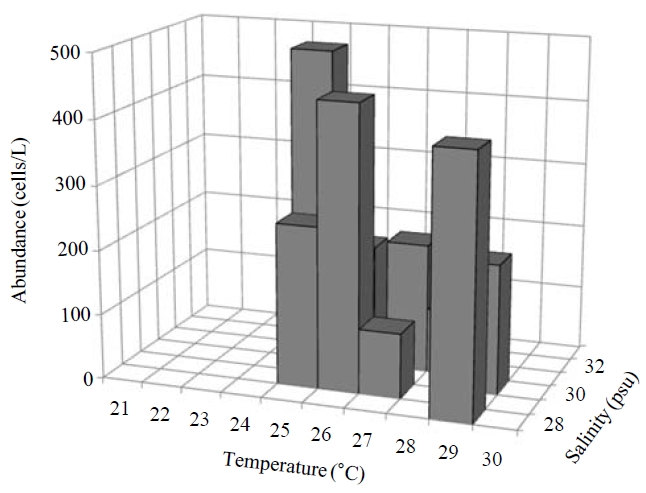
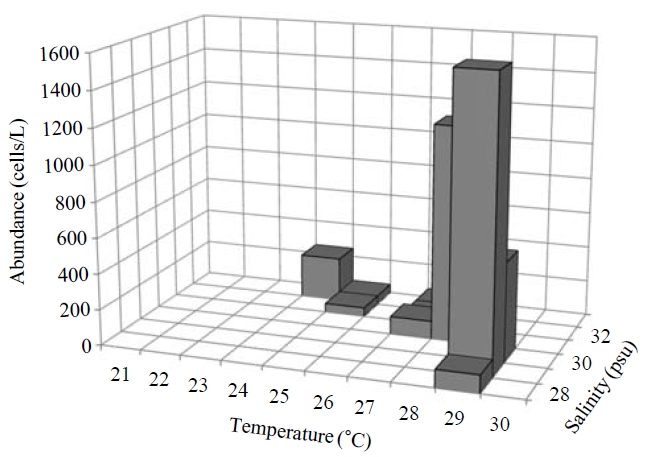
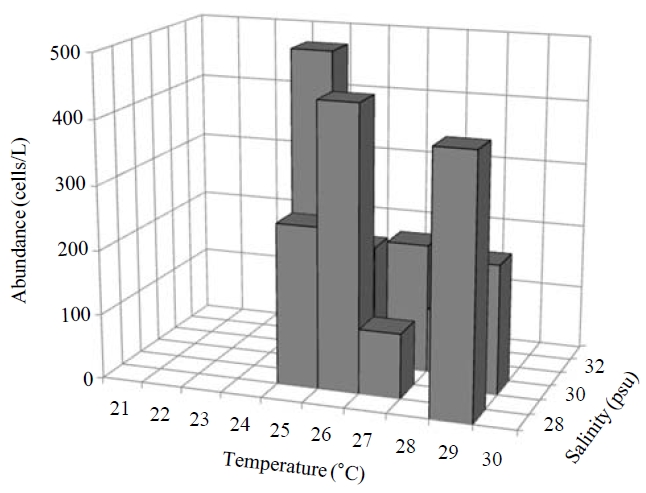
Occurence and ecological characteristics. Maximum abundance(ca. 1,600 cells/L) of this species was observed at M4 site in August, 2007 where water temperature of 29.13℃ and salinity of 28.69 psu were recorded respectively. This species was strictly found at high temperatures (24-29℃) period (Fig.5).
Comparison with Chinese population.
in morphology: marine species collected from coastal water in salinity about 30 psu; cell shape; the number of AM and VM; the presence and number of pTM, and the shape and number of Ma. However, cell size of Korean population is smaller than Chinese population (43-82×28-45 μm of Korean population vs. 60-80×40-50 μm of Chinese population).Number of Ma of Korean population is larger than that of Chinese population (35-62 vs. 20-30). Chinese population is vague about the appearance of an AP but Korean population shows a conspicuous protrusion at anterior end of the cell after protargol impregnation.
1*Family Tontoniidae Agatha, 2004
2*Genus Spirotontonia Agatha, 2004
3*Spirotontonia turbinata (Song and Bradbury, 1998)
(Table 2,Fig.6 )
Spirotontonia turbinata Song et al., 2009: 334.
Diagnosis. Body inverted triangle-shaped with tail on dorsal side of posterior cell end; body size, 59-110 μm in length and 29-51 μm in width after protargol impregnation; 13-15 AM and 15-20 VM; sinistrally spiral GK (about 2.5 whorls);no VK; ellipsoidal Ma, size 3-3.5 μm in diameter, and 50-60 in number.
Morphological description of Korean population. Body obconical to inverted triangle with bluntly pointed posterior end, slightly leftward inclined anterior portion of body. Body size, 59-110 μm in length and 29-51 μm in width (average 77×37 μm, n=9). Tail located above the second whorl of GK on dorsal side, easily detached from body during process of handling or staining. A kinety consisting of densely spaced basal bodies with fibers on front of tail. Body covered with transparent DC between whorls of GK (Fig.6 A-D).
Buccal cavity open on ventral side, extending slightly to right and terminating approximately 1/2 of way down cell.AM and VM positioned like question-marked (“?”) on ventral side. About 13-15 AM (average 14, n=8). VM extended along left side of buccal cavity and consisting of 15-20 membranelles(average 17, n=9). A row of EM lying on inner wall of buccal lip on right inner side of oral cavity, consisting of densely arranged monokinetids (Fig.6 A, C).
Sinistrally spiral GK orientating on left side of buccal cavity,extending to left lateral side along posterior margin of VK then spirally turning down to posterior of body with about 2 whorls (average of GK per 10 μm, 12). “L” shaped fibres located above GK and arranged along GK (average of F per 10 μm, 9). No VK detected.
Ellipsoidal Ma scattered in whole of body, size 3-3.5 μm in diameter (average 3 μm in diameter, n=3), 50-60 in number(average 55, n=3).
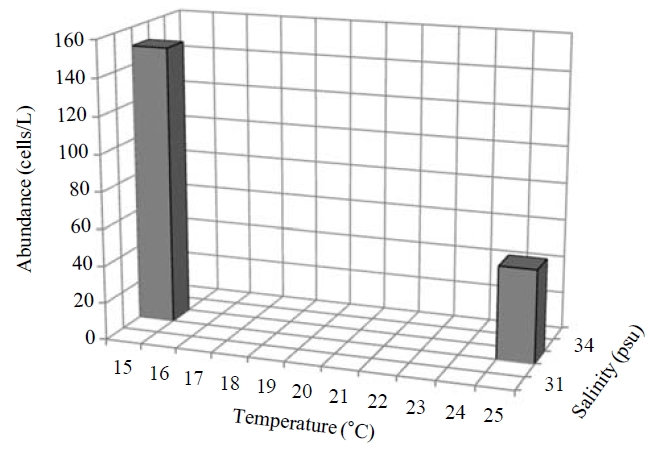
Occurence and ecological characteristics.This species were collected in November 2006 and in August 2008 from Masan Bay (water temperature of 15-23.27℃ and salinity of 30.30-32.6 psu) and in June 2009 from Jangmok Bay (water temperature of 17.45-23.27℃ and salinity of 30.30-31.51 psu). Maximum abundance of 150 cells/L showed at M20 and M30 sites in August 2006 where water temperature of 15C and salinity of 32.6 psu were recorded respectively (Fig.7).
Comparison with other populations.
4*Spirotontonia grandis (Suzuki and Han, 2000)
(Table 2,Fig.8 )
Tontonia grandis Suzuki and Han, 2000: 989-994.
Spirotontonia grandis Agatha, 2004a: 298.
Diagnosis. Body obconical with screw-like appearance; size 87-157.5 μm in length and 32-52 μm in width after protargol impregnation; a conspicuously apical collar (AC) located at anterior pole; tail on dorsal side; 13-15 AM and 23-34 VM;sinistrally spiral GK of three whorls, oriented below buccal area and terminated at posterior end of dorsal side; no VK;37-84 ellipsoidal Ma, about 3-4 μm in diameter.
Morphological description of Korean population. Body screwed barrel in shape with AC at anterior pole, posterior of cell bluntly pointed. AC region truncate- or dome-shaped.Cell size 87-157.5 μm in length and 32-52 μm in width (average 108×41 μm, n=23) after protargol impregnation. Tail broad leaf shaped, located above the second whorl of GK on dorsal side, easily detached from body during handling in staining process. A short kinety positioned across front view of tail, consisting of densely spaced basal bodies and fibers (Fig.8 F). Back view of tail wrinkled. Transparent DC covered between whorls of GK (Fig.8 A, B). Cortical platelets observed in several specimens, having elliptical shape and covering on DC surface (Fig.8 G).
Buccal cavity open, extending obliquely to right and terminating about 1/2 of way down cell. AM surrounding AC region, consisting of 13 to 15 AM (average 14, n=22). VM extending along left side of buccal cavity, consisting of 23-34 membranelles (average 28, n=23). A row of EM lying on
inner wall of buccal lip on right side of buccal cavity, consisting of monokinetids.
Sinistrally spiral GK, with a total of three whorls, located below posterior part of buccal cavity, extending to left lateral side along posterior margin of VK and turning around dorsal side horizontally, then sharply going down to proximal part on ventral side and then spirally going down two whorls,finally terminating at posterior end of dorsal side. F (probably fibers) parallel arrangement above GK; “L”-shaped in
stained (the number of kinetids per 10 μm GK=15 and F=7 on average). No VK detected.
Oval or irregular shaped Ma scattered in whole of body,3-4 μm in diameter (average 3 μm, n=19), and 37-84 in number(average 65, n=19).
Occurence and ecological characteristics. This species were collected from Masan Bay in August 2006-2008 and Jangmok Bay in June 2009. During the survey period, this species was found only in summer at high water temperature(¤17℃). In Jangmok Bay, this species and
Comparison with other populations.
Korean name: 1*개구환소모충목, 2*민소모충과, 3*오메가열소모충속, 4*다핵오메가열소모충 Korean name: 1*꼬리소모충과, 2*나선체꼬리소모충속, 3*원추나선체꼬리소모충, 4*큰나선체꼬리소모충