Olive flounder
The intake of water is essential for all aquatic and terrestrial animals (Adolph, 1933). This requirement can be met either by the direct intake of water or by chemical reactions in the body (metabolic water).Dietary water, which is usually noted as ‘other dietary components’ may require more attention as it can influence the digestive process and uptake of dietary nutrients. Water is the least expensive ingredient in fish feeds and its appropriate use may provide a cost-effective method of improving fish growth and uptake of other dietary ingredients. Previous studies have shown an improvement in growth and nutrient retention attributable to a faster nutrient release from the stomach and increased digestibility (Grove et al., 2001).
Gastric evacuation is affected by various factors,including water temperature, fish size, diet composition, and meal size (Bromley, 1987; Naik et al., 2000; Sweka et al., 2004). Information on the gastric evacuation rates of fish is useful to assess appetite return and allow an estimate of the proper feeding regimen. The feeding rate is one of the most important factors in the growth of fish (Brett, 1979). The optimum feeding rates not only enhance the growth, survival, and feed conversion ratio, but also assist the minimization of food wastage, size variation, and ultimate production costs.
In Korea, the majority of fish farmers supplement dry pelleted diets with water during olive flounder production. However, limited information on the effects of the additional water in the dry pellets is available. In this study, we investigated the effects of the dietary moisture and feeding rate on the growth and gastric evacuation of young olive flounder.
Olive flounders were obtained from a local farm(Pohang, Korea). The fish were acclimated to rearing conditions for 4 weeks prior to initiating the feeding trial. After acclimatization, the fish (average weight, 106±1.4-g) were randomly allocated to 181,500-L cylindrical plastic tanks with 20 fish in each tank. Filtrated seawater was supplied at a flow rate of 20 L/min in each tank, and the water temperature was maintained at 22±1.6°C (mean±SD). The photoperiod was left under natural conditions during the feeding trial. All fish in each tank were collectively weighed at the beginning of the feeding trial and every 5 weeks thereafter.
Four levels of dietary moisture were prepared: 9%, 21%, 30%, and 40%. For the 9% moisture trial,commercial feed was used, while the 21%, 30%, and 40% moisture levels were obtained through mixing the appropriate volume of deionized water (15, 30,and 50 g/100 g feed) to the commercial feed (Jeilfeed, Daejeon, Korea) (crude protein: 49.4%, crude lipid:12.9%, ash: 9.2%). The moisture-containing pellets were stored at -20°C until use.
Three replicate groups of fish were fed the experimental diets containing 9%, 21%, 30%, and 40% moisture to satiation or 9% and 30% moisture at restricted feeding rates (95% satiation) twice a day(09:00 and 16:00) for 15 weeks. Satiation feeding was determined by the point of cessation of the voluntary feeding activity of the fish. Uneaten food was removed and weighed after the fish had stopped eating and deducted from the feed consumption. The feed allowance of fish in the restricted feeding group was determined based on the average consumption of fish fed diets with the same moisture levels in the satiation feeding groups.
Following the feeding trial, the gastric evacuation rates were measured using 14 fish from each tank.The fish were fed the experimental diets to visual satiety, while the other conditions were maintained in the feeding trial. The fish were anesthetized with 100 mg/L MS222 (Sigma, St. Louis, MO, USA) and killed for dissection of the gut. The feed contents,expressed as wet weight (g) of feed×100/fish (g), in the stomach and intestine (Brett and Higgs, 1970)were collected at 0 (immediately after feeding), 3, 6,12, 18, 24, and 30 h after feeding. The contents of the stomach and intestine were removed by gentle pressure.
>
Sample collection and analytical methods
At the end of the feeding trial, three fish from each tank were collected and stored at -75°C for chemical analysis. The samples of diet, dorsal muscle, and the contents of the fish stomachs were analyzed for proximate composition. Crude protein was determined by the Kjeldahl method using the Auto Kjeldahl System (VAP50OT/TT125; Gerhardt, Konigswinter,Germany). Crude lipid was determined by ether extraction using a Soxhlet extractor (Velp SER 148;Usmate, Milano, Italy). The moisture levels were determined by oven-drying at 135°C for 2 h. Ash was determined using a muffle furnace at 550°C for 6 h.
The data were subjected to a one-way analysis of variance (ANOVA), and when significant (
>
Growth performance and body composition
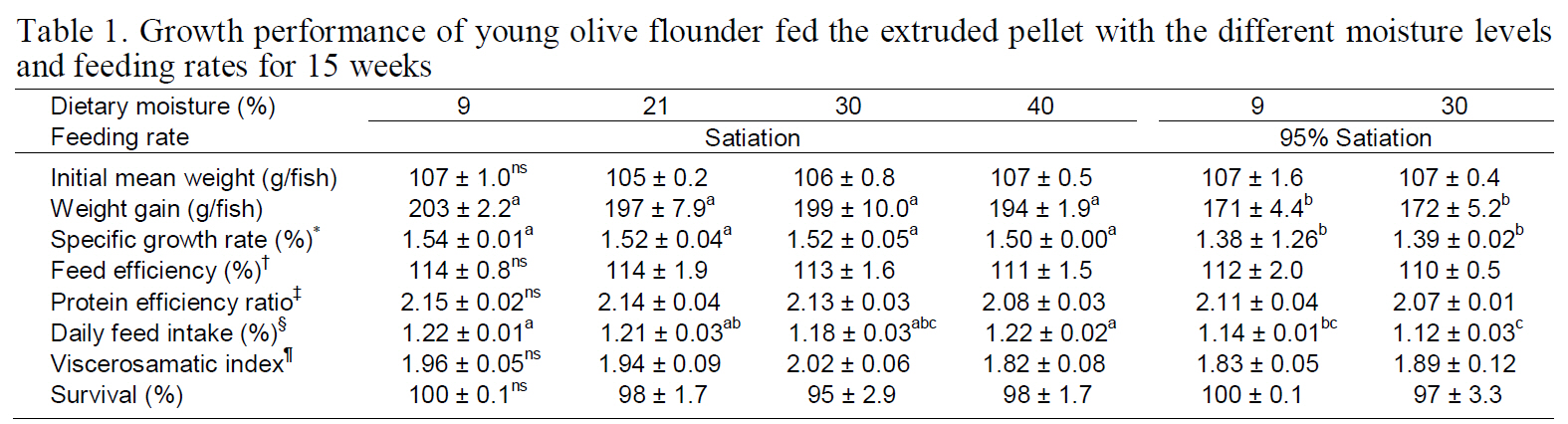
The growth performances of fish receiving the 9%, 21%, 30%, and 40% moisture diets at a different feeding rate for 15 weeks are shown in Table 1. The dietary moisture levels did not significantly affect the weight gain or the specific growth rate. However, the weight gain and specific growth rate of the fish fed 9% and 30% moisture diets to 95% satiation were significantly lower than those of fish receiving 9-40% moisture diets to 100% satiation (
Growth performance of young olive flounder fed the extruded pellet with the different moisture levels and feeding rates for 15 weeks
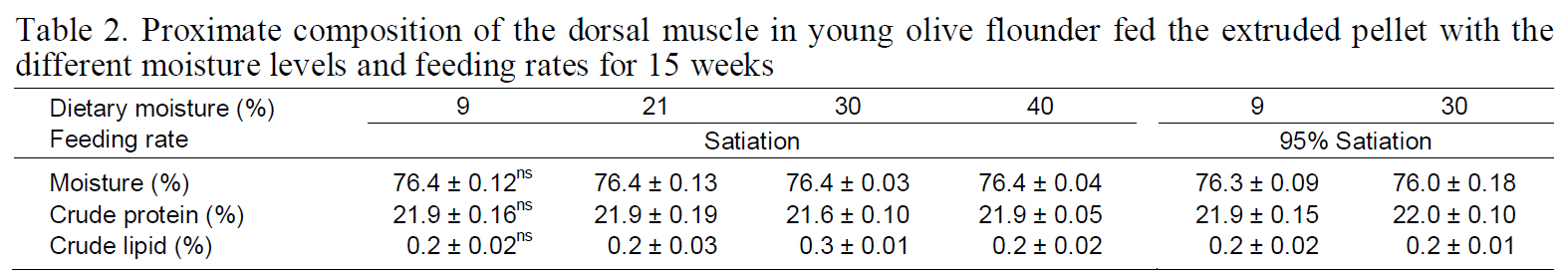
Proximate composition of the dorsal muscle in young olive flounder fed the extruded pellet with the different moisture levels and feeding rates for 15 weeks
treatment groups.
The approximate composition of the dorsal muscle in the fish fed 9%, 21%, 30%, and 40% moisture diets at different feeding rates for 15 weeks are shown in Table 2. The moisture, crude protein, and crude lipid content of the dorsal muscle did not significantly differ among the groups.
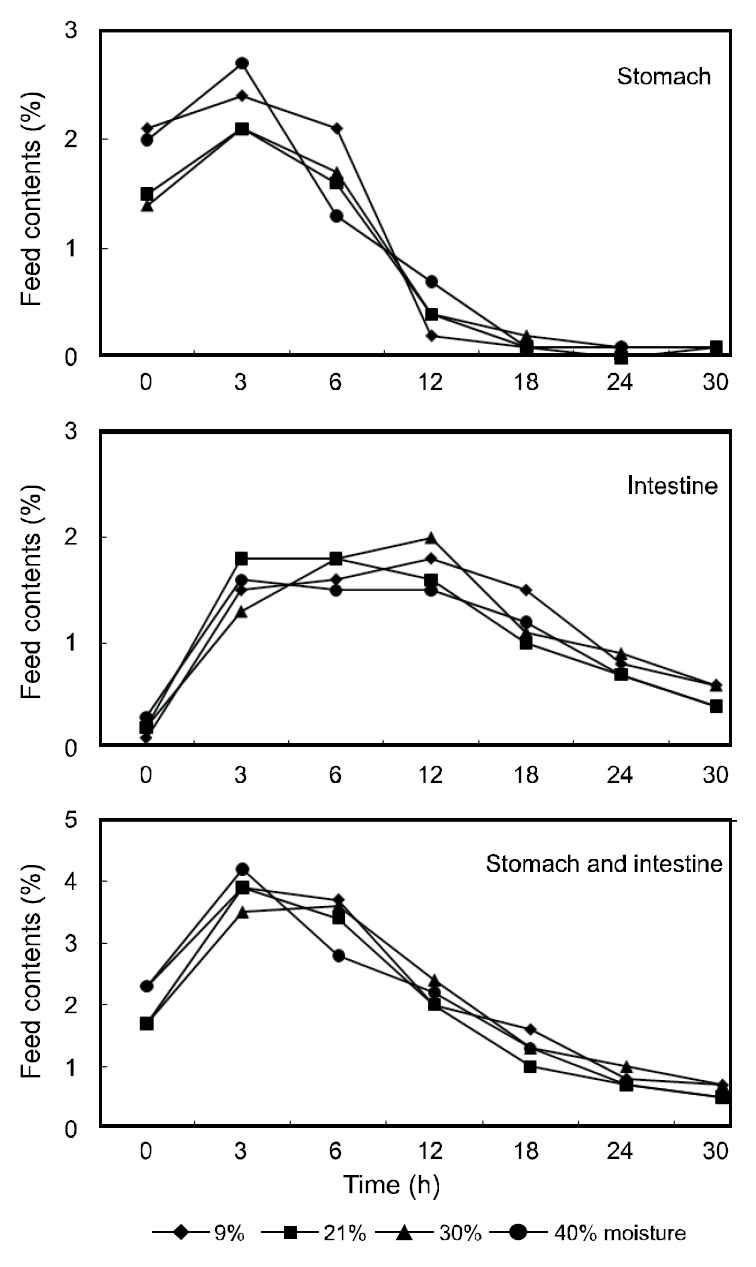
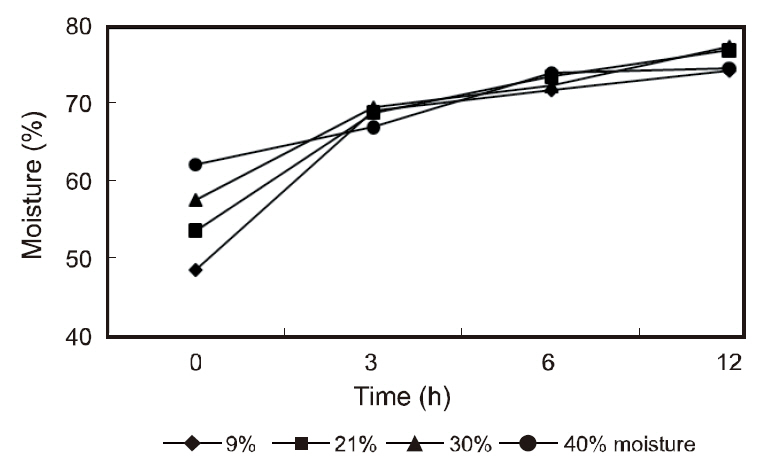
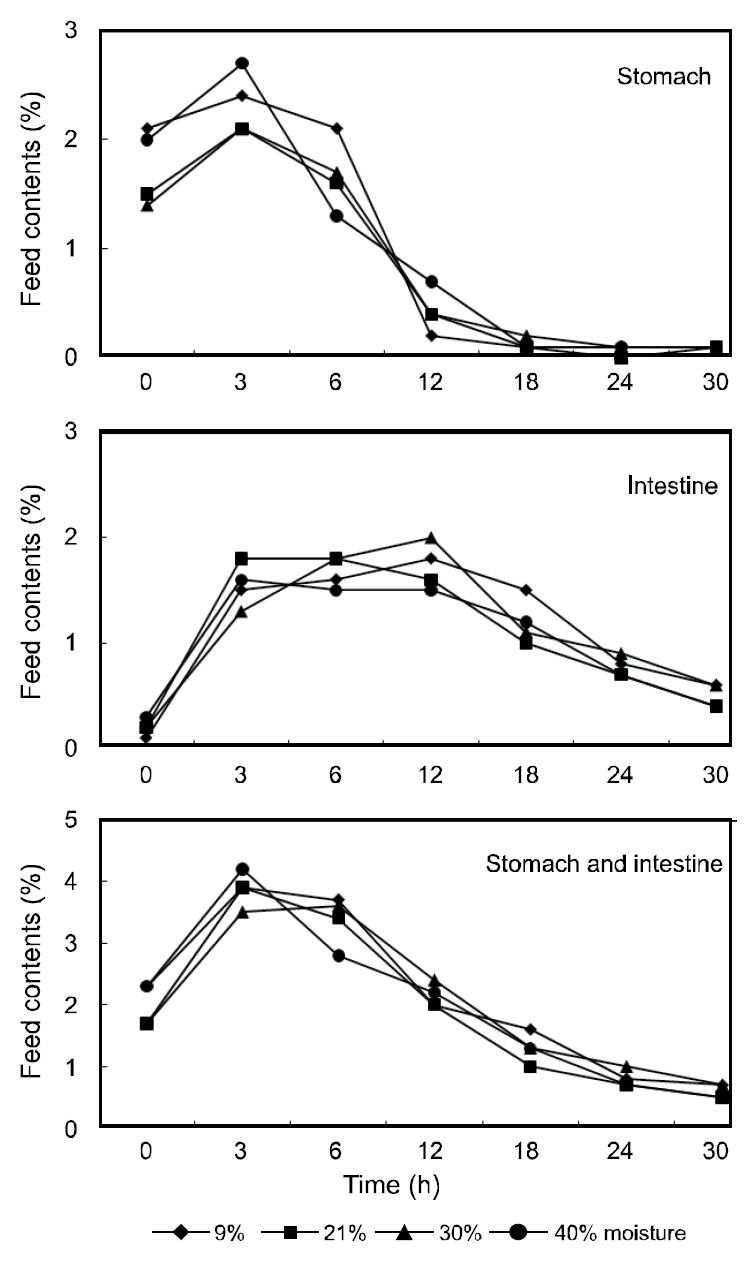
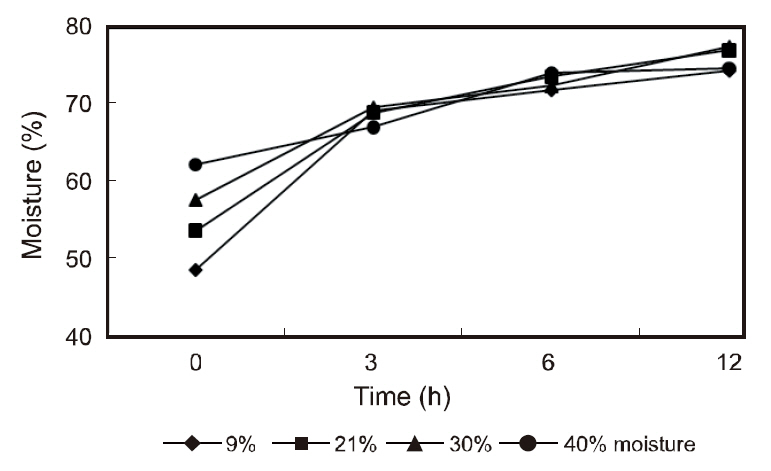
The gastric contents of the fish receiving the various moisture diets are shown in Fig. 1. The stomach contents peaked within 3 h of feeding and gradually decreased thereafter with the stomachs of the fish being completely evacuated within 18 h. The contents of the intestine peaked at 3-12 h post-feeding and began to decline at 30 h until the intestine was almost completely evacuated. The moisture of the stomach contents in the various diets are shown in Fig.2. The moisture of the stomach contents reached approximately 70% within 3 h post-feeding and gradually increased to approximately 75% within 12 h. No considerable differences were observed in the gastric evacuation and stomach moisture content in the fish receiving the different moisture diets.
In this study, an increase in dietary moisture from 9% to 40% displayed no beneficial effects on the growth and feed utilization of olive flounder (106 g average initial weight). Similar nonsignificant differrences for the weight gain and feed efficiency were observed in the subadult flounder (332 g) fed diets containing 7-15% moisture under the rearing water temperature of 21°C, as well as in 646-g flounder fed between 8% and 23% moisture diets at the suboptimal water temperature of 12°C (Kim et al.,2011). Lee et al. (2000b) reported that dietary moisture content displayed no significant effects on the weight gain or feed efficiency of Korean rockfish
Marine fish may require dietary water for osmoregulation between the body and medium (Ruyet et al., 1982; Higgs et al., 1985), and the growth-promoting effect of moisturized diets could be attributable to a faster nutrient release from the stomach, leading to higher digestibility (Grove et al., 2001). Other reasons for the improved growth when receiving moist diets include an improved acceptance of soft versus hard feed particles (Stradmeyer et al., 1988). For dentex, fish fed 7% water diets displayed smaller stomachs than those fed a 40% water diet (Chatzifotis et al., 2005). The stomach volumes of the herring diet-fed rainbow trout
This study showed a similar stomach moisture content in fish fed diets of varying moisture levels within 3 h of feeding. Similarly, after feeding, the feed was hydrated, and within 2 h, the stomach digesta reached similar water content levels to dentex that had received 7%, 20%, and 40% water diets (Chatzifotis et al., 2005). Fish require moisturized ingested food prior to digestion (Hughes and Barrows, 1990) and dry pellets impose a demand for water during gastric processing. Fish hydrate their digesta through drinking during ingestion and stomach secretion (Kristiansen and Rankin, 2001). The ability of fish to moisturize their digesta varies on the fish species (Hughes and Barrows, 1990). A higher water content (70%) in the stomach digesta of olive flounder 3 h post-feeding was observed in this study compared to the 60% reported for dentex (Chatzifotis et al., 2005). Certain species not being able to moisturize their digesta may decrease their efficiency of digestion, resulting in a reduced growth performance (Hughes and Barrows, 1990).
The gastric contents of fish are affected by properties of the diet including its composition of ingredients and its moisture content (Poston, 1974; Grove et al., 1978; Sveier et al., 1999). However, the results of this study indicate that the gastric evacuation rates of olive flounder were not affected by the dietary moisture content. Similarly, Garber(1983) reported no significant differences in the evacuation rates between yellow perch fed 4.7% and 27.1% moisture diets. For dentex, no differences in the gastric evacuation rates were observed among fish fed diets containing 7%, 20%, and 40% moisture (Chatzifotis et al., 2005). In this study, at 30 h postfeeding,the intestine of the flounder was primarily evacuated. However, subadult flounder (848 g) fed diets containing 8% and 23% moisture under a rearing water of 12°C required 56 h to evacuate their intestines in previous studies (Kim et al., 2011). The variations in gastric evacuation time may be due to differences in the fish size, the dietary composition, or the water temperature employed during these studies.
Olive flounder in the restricted feeding groups showed considerably lower weight gain than that of fish of the satiated feeding group. Similar results have been reported in previous flounder studies (
The proximate composition of the dorsal muscle was not affected by the dietary moisture levels and feeding rates in this study, consistent with previous reports (
Taken together, this study demonstrates that the gastric evacuation of olive flounder is not affected by dietary moisture levels and that the addition of water into the diet displays no beneficial effects on the growth of olive flounder.