The green algal genus Codium occurs in various marine habitats, from estuarine to fully marine and from soft to hard substrata. Codium is cosmopolitan, except for the Arctic and Antarctic Oceans. Species range from crustose forms to erect highly branched or flattened forms. Nevertheless,most species have a dark green color, spongy texture, and multi-axial thalli up to approximately 1 m in length (Graham et al. 2009). Currently, Codium consists of 130 species, 14 subspecies, three varieties, and three forms (Guiry and Guiry 2011). The major morphological characters used for identification are shape of thalli,utricles, and gametangia as well as branching patterns(Yoshida 1998). Codium thalli are composed of numerous utricles with an interior medulla, consisting of an entangled mesh of siphon structures. The most apparent and variable attributes are external morphology in which thalli spread out over the surface as mats, finger-like, or flattened branches (Verbruggen et al. 2007). Some species are broadly expanded with a velvety and smooth texture.
In Korea, a carpet-like Codium has been reported as Codium latum Suringar (Kang 1968, Oh et al. 1987, Sohn 2000). Kang (1968) described its features based on specimens from Busan and Jeju, Korea (Fig. 1A) as follows:broad and flat, or long-spread thallus, with a silky texture,and cylindrical utricles with swollen and rounded apex. Sohn (2000) described the morphology and analyzed trnL gene sequences from C. latum collected from Jeju Island, Korea and Tateyama, Japan. Lee and Kang(2001) recorded C. latum in “A Catalogue of the Seaweeds in Korea.” Sixteen species were recently reported by Lee(2008), including a new carpet-like species, Codium ta-
petum Y. Lee, collected from Jeju Island. Lee (2008) noted that C. tapetum was previously identified as C. latum, and characterized C. tapetum as having thalli with a broadly expanded frond with a short stipe, and sub-pyriform utricles with a slightly swollen apex.
In Japan, Shimada et al. (2004) conducted phylogenetic studies on 18Codium species, including carpet-like species, and concluded that rbcL gene regions are valuable for distinguishing between Codium species. Shimada et al. (2007) published a new carpet-like species as C. tenuifolium from Japan (Fig. 1B), rejecting the name C. pseudolatum, although they found C. pseudolatum named by Yamada in a herbarium specimen known as a nomen nudum. Because the name Codium pseudolatum was previously published by Nizamuddin (2001) from the Arabian Sea. C. tenuifolium is characterized by thin blades, clavate utricles with a tumid head, and variable reproductive structures at the utricle base (Shimada et al. 2007). Currently, there are three carpet-like Codium species, C. latum, C. tapetum, and C. tenuifolium, reported from Korea and Japan (Suringar 1867, 1870, Okamura 1915, Schmidt 1923, Kang 1968, Oh et al. 1987, Sohn 2000, Shimada et al. 2004, Shimada et al. 2007, Lee 2008).
During a survey of green algal diversity, we encountered Codium species with carpet-like thalli from several localities on Jeju Island, Korea. In this study, we confirmed the taxonomic position of the carpet-like species, currently named C. tapetum Y. Lee from Korea (2008) and C. tenuifolium Shimada, Tadano and J. Tanaka from Japan (2007). We examined morphological characteristics and sequenced rbcL gene using our recent specimen collections.
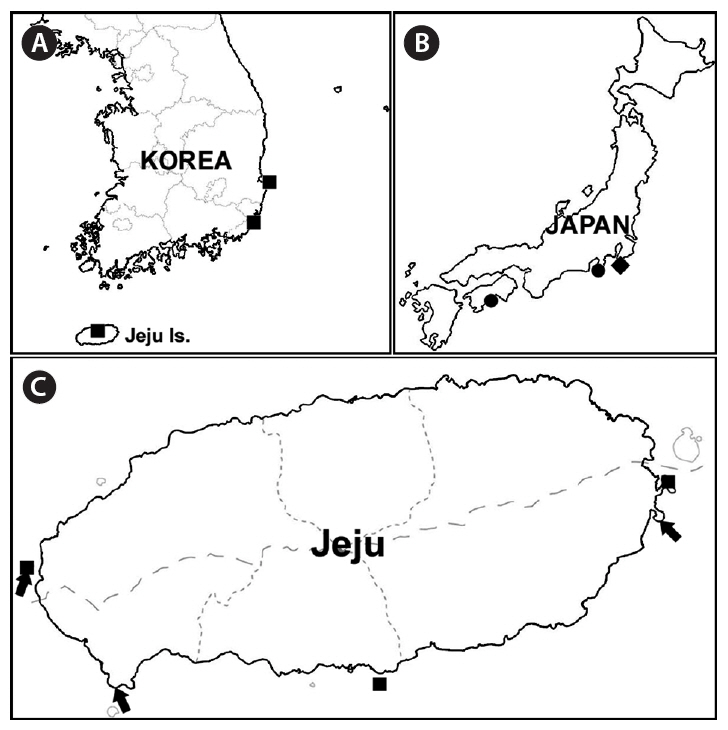
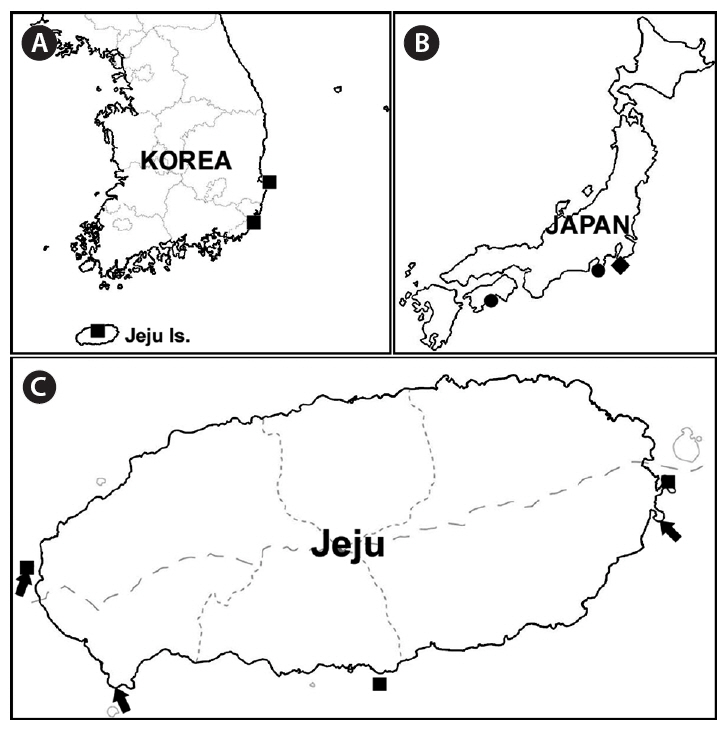
The distributions of carpet-like Codium species from Korea and Japan can be seen on the map (Fig. 1A-C). Six samples of carpet-like Codium were collected from Jeju Island (Fig. 1C): 10TE001, 10TE002, April 6, 2010, Shinyang, J. C. Kang; 10TE003, June 6, 2010, Chaguido, Y. D. Koh; 10TE004 (gametangia), 10TE005 (gametangia), June 16, 2010, Sangmo-ri, E. G. Han; 10TE006, August 13, 2010, Sangmo-ri, E. G. Han. The samples were stored in silica gel for molecular studies, and DNA vouchers were prepared from two fresh samples. The remaining samples were preserved in formalin-seawater for morphological observations. Habit photographs were taken with a D-80 camera (Nikon, Tokyo, Japan), and internal morphology was photographed using QIMAGING 1394 camera (QImaging, Surrey, BC, Canada) with a BX50 microscope (Olympus, Tokyo, Japan). DNA vouchers and specimens were stored at the herbarium of Jeju National University (JNUB), Korea.
DNA was extracted from dry samples using the DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) following the manufacture’s protocol. Extracted DNA was stored at -20°C and was used to amplify rbcL gene. Polymerase chain reaction (PCR) amplification was performed using rbcL primers designed by Shimada et al. (2004). The forward primer was EadF (5′?GACTAACTTATTATACACCGG-3′; 21mer), and the reverse primer was EadR (5′?CACCAGTTTCAGATTGCGATT-3′; 21mer). PCR amplification was conducted in a total volume of 20 μL with AccuPower PCR PreMix (Bioneer, Daejeon, Korea). PCR was conducted with an initial denaturation at 95°C for 5 min, followed by 35 cycles of amplification (denaturation at 94°C for 1 min, annealing at 52°C for 1 min, and extension at 72°C for 2 min), with a final extension at 72°C for 7 min. The PCR products were purified using the AccuPrep PCR Purification Kit (Bioneer).
To confirm the taxonomic position of the carpet-like Codium from Jeju, maximum likelihood (ML) analysis was performed, using PAUP 4.0 (Swofford 2002). Six rbcL sequences of Codium were aligned with other Japanese
Codium species. Two Bryopsis species were used as the outgroup. We performed a likelihood ratio test using MODELTEST (version 3.7) (Posada and Crandall 1998) to determine the best available model for our sequence data. The best model was a general time reversible (GTR) model with a gamma correction for among-site variation (Γ) and the proportion of invariable sites (I). ML trees were estimated using a heuristic search with 100 random addition sequence replicates, and tree bisection and reconnection (TBR) branch swapping. ML bootstrap analyses were performed with 1,000 replicates to test node stability.
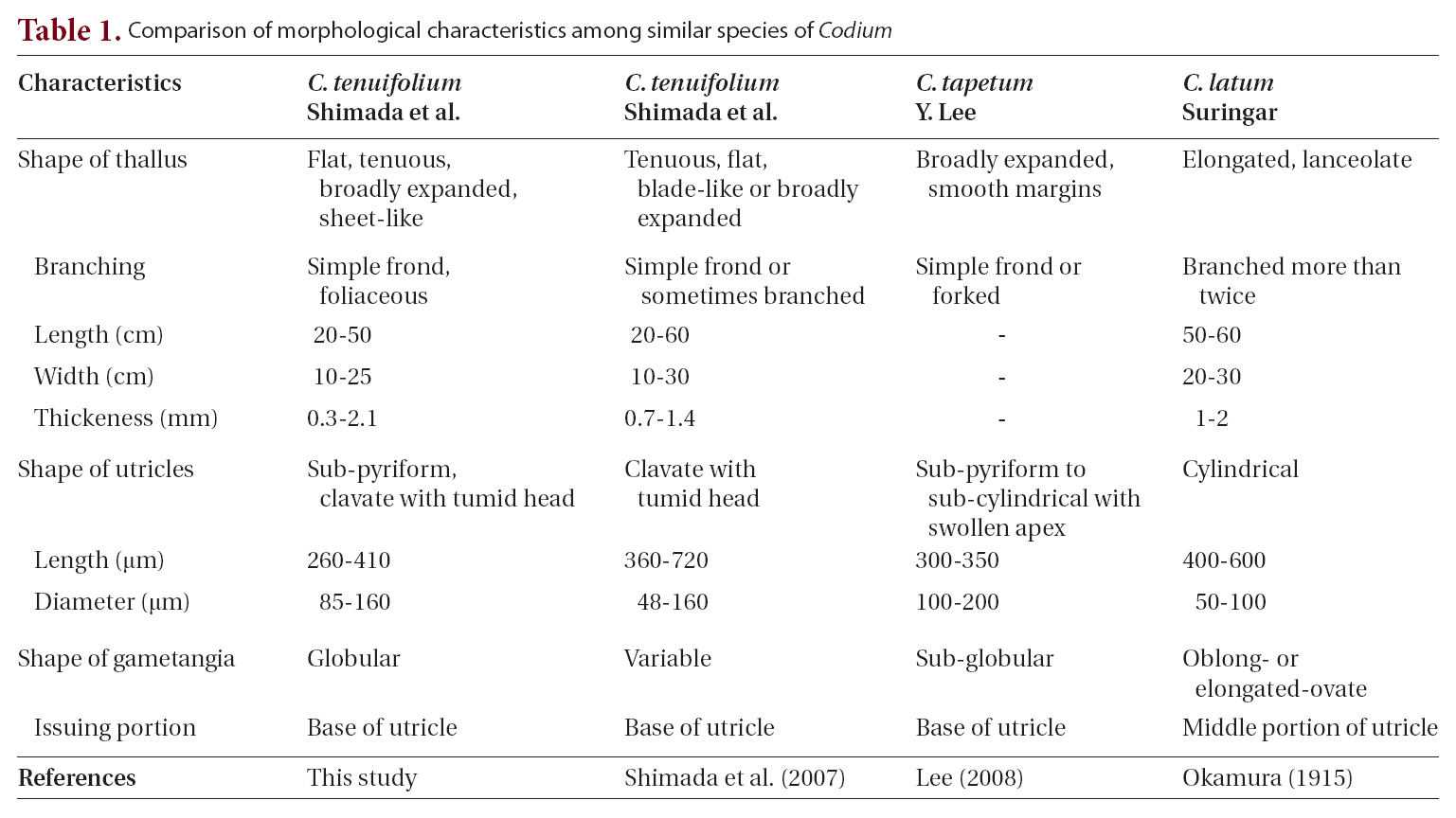
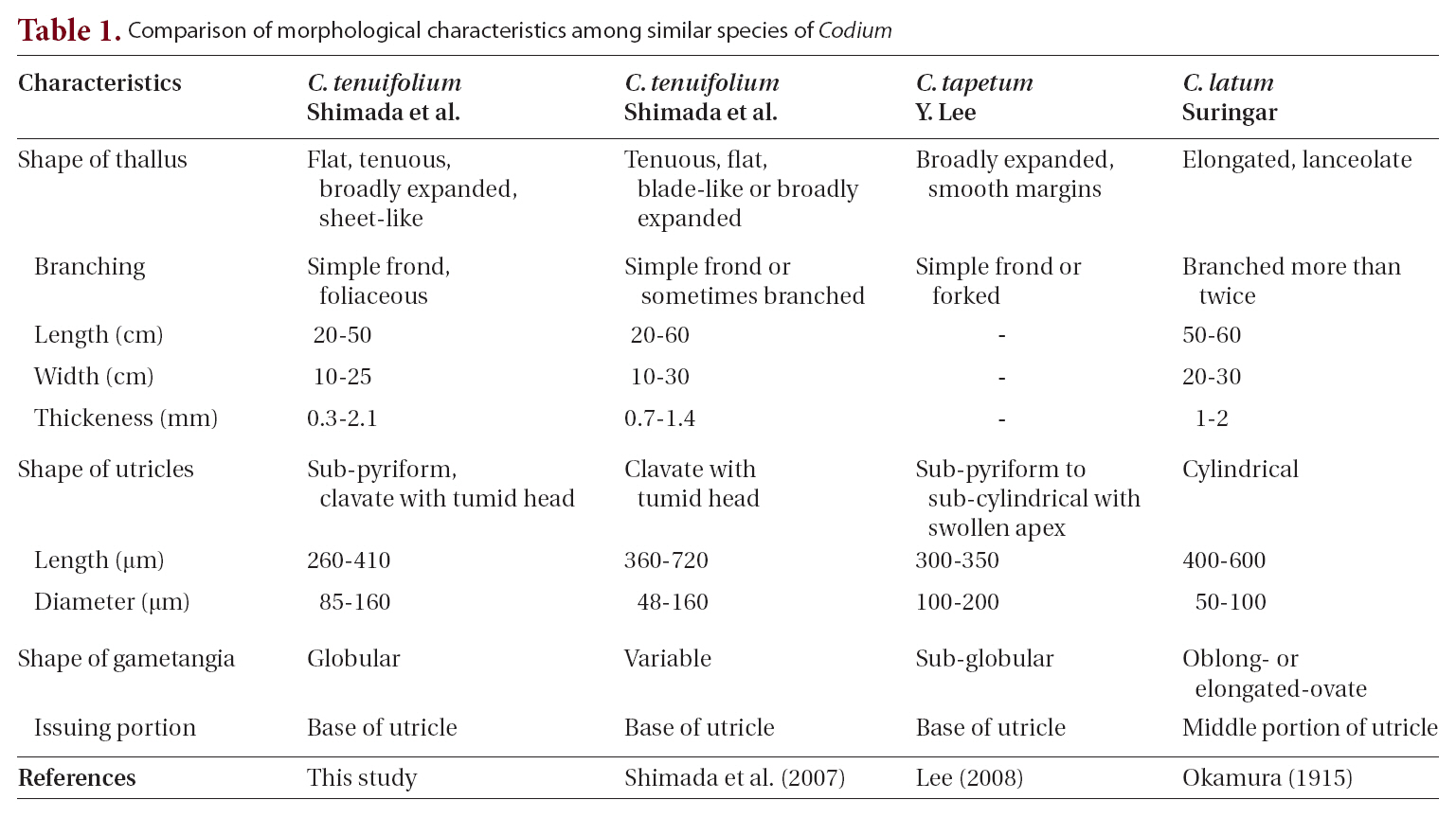
Thalli arise from a discoid and concave holdfast attached to a shell or rock surface. Thalli have a stipe less than 1 cm long and 3-5 mm in diameter. The stipe is either terete or slightly compressed. The thallus is broadly expanded from a stipe and has a foliaceous and bi- or trifurcate shape, rather than branched, and is dark green in color (Fig. 2A-D). Thalli are 20-50 cm long, 10-25 cm wide, and 0.3-2.1 mm thick. The thallus has thin, fragile,
sheet-like, and velvety texture.
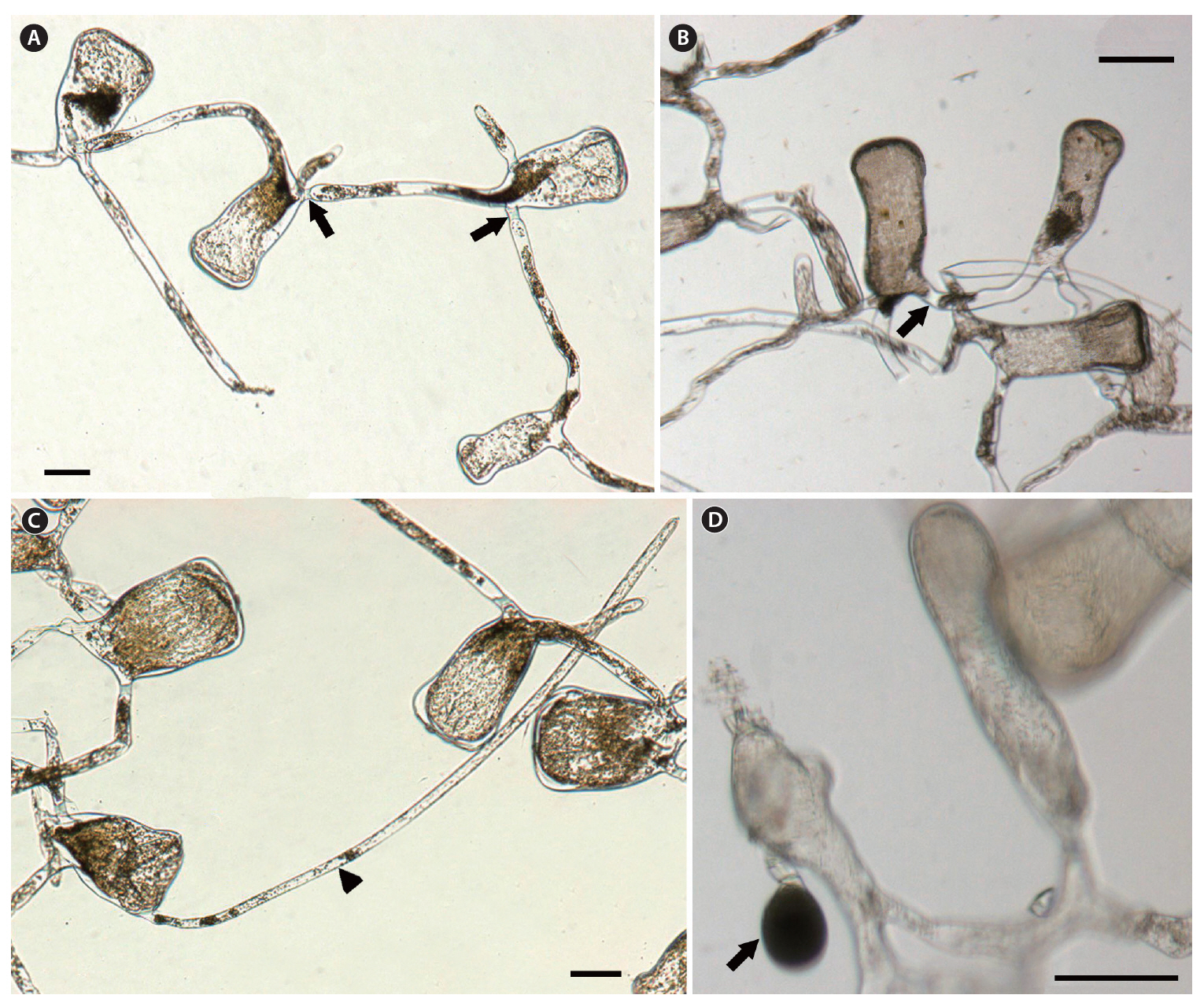
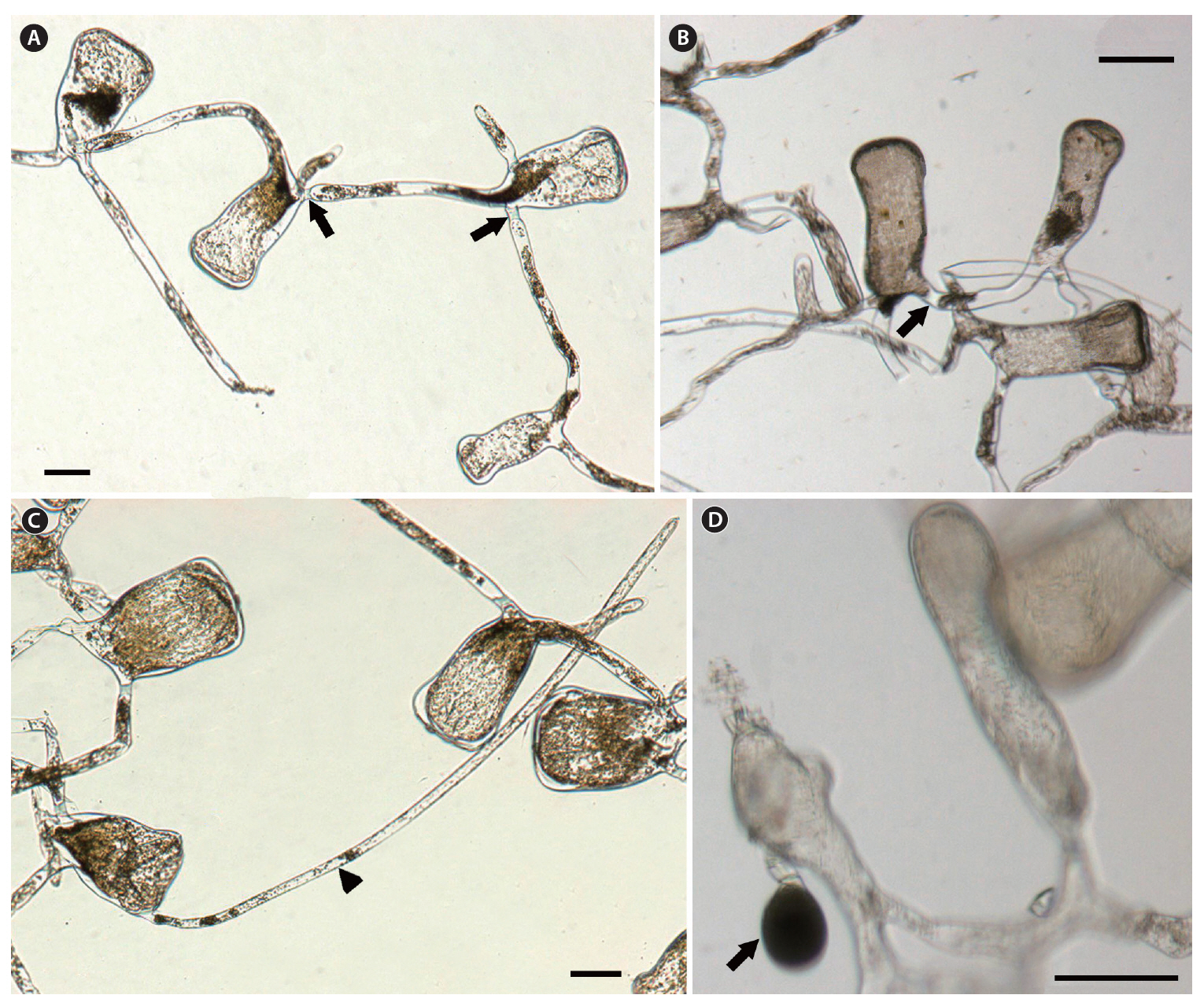
The thallus is composed of numerous concatenate utricles with entangled siphons, creating the medullary filaments. Two or three siphons are linked to each utricle base, and extend towards nearby utricle bases (Fig. 3A & B). The siphon diameter is 28-41 ㎛, and typically has an H-shaped septum (Fig. 3A & B) on one side. The utricle is cylindrical, sub-pyriform and clavate, and the utricle base is not as thick as the apex. The utricle body is approximately 85 ㎛ in diameter, and the apex is 110-160 ㎛. The utricle apex is flat and rarely concave, and becomes slightly more tumid and swollen compared to the basal and middle sections (Fig. 3A-C). Utricles are 260-410 ㎛ long, and the length is three times longer than the width. Utricles are generally hairless, but hairs up to 700-1,000 ㎛ long can grow on the apical section of the utricle (Fig. 3C). Globular gametangia dangle from the short pedicel, and are tinged with a dark green color. They develop irregularly on the siphons, following initiation at the utricle bases (Fig. 3D).
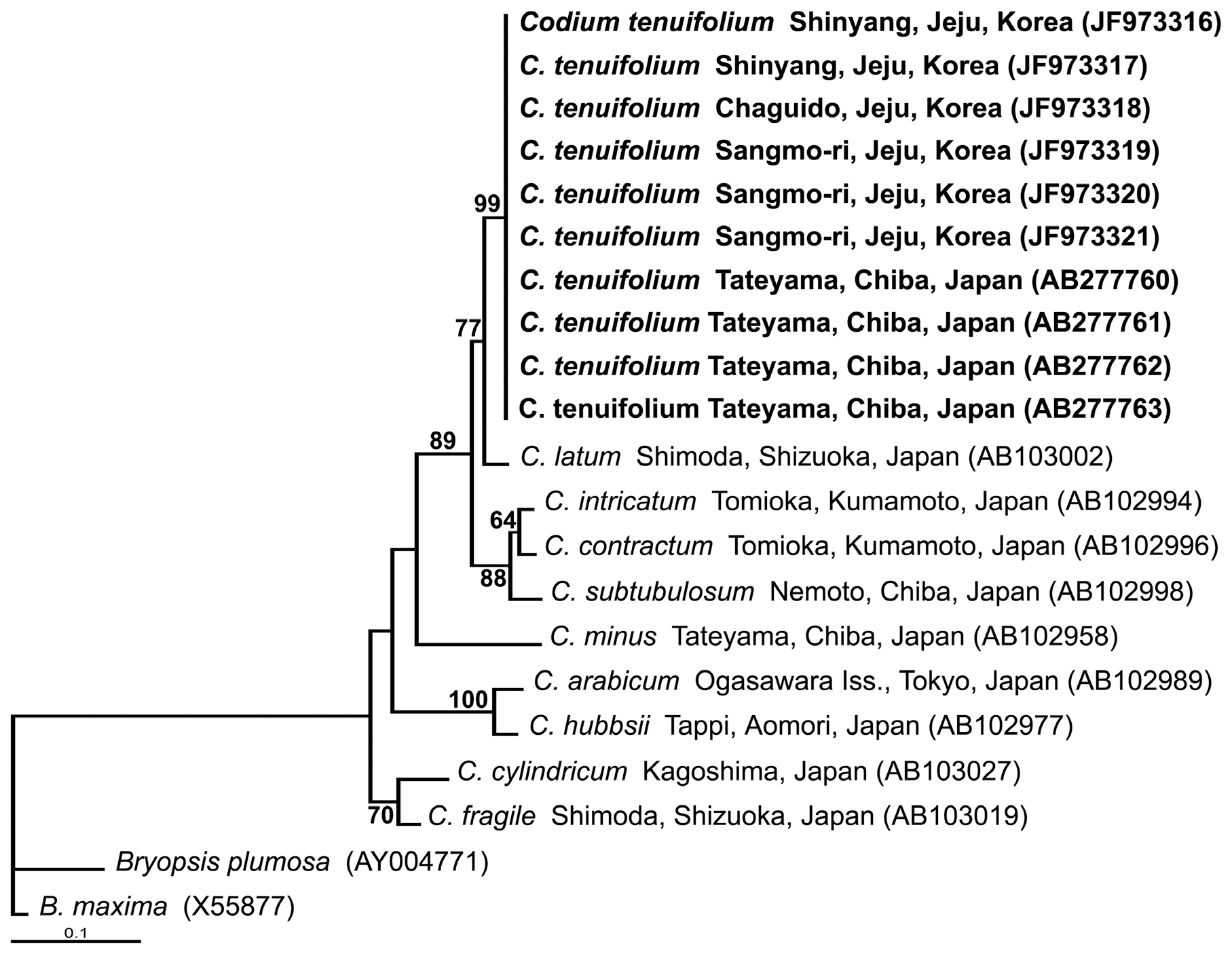
We sequenced six carpet-like Codium specimens from Jeju for rbcL, and aligned these with four Japanese C. tenuifolium specimens, and nine Codium species from GenBank. In total, 602 bp of rbcL were aligned: 143 sites (23.8%) were variable and 108 sites (17.9%) were phylo-
genetically conserved. There were excesses of adenine (28.9%) and thymine (30.9%) at all codon positions. Six carpet-like Codium specimens from Jeju were identical to four C. tenuifolium specimens from Japan. The C. tenuifolium sequence divergence differed by 2.82% from C. latum, 5.0% from C. intricatum, 5.3% from C. subtubulosum, 5.3% from C. contractum, and 9.1% from C. minus.
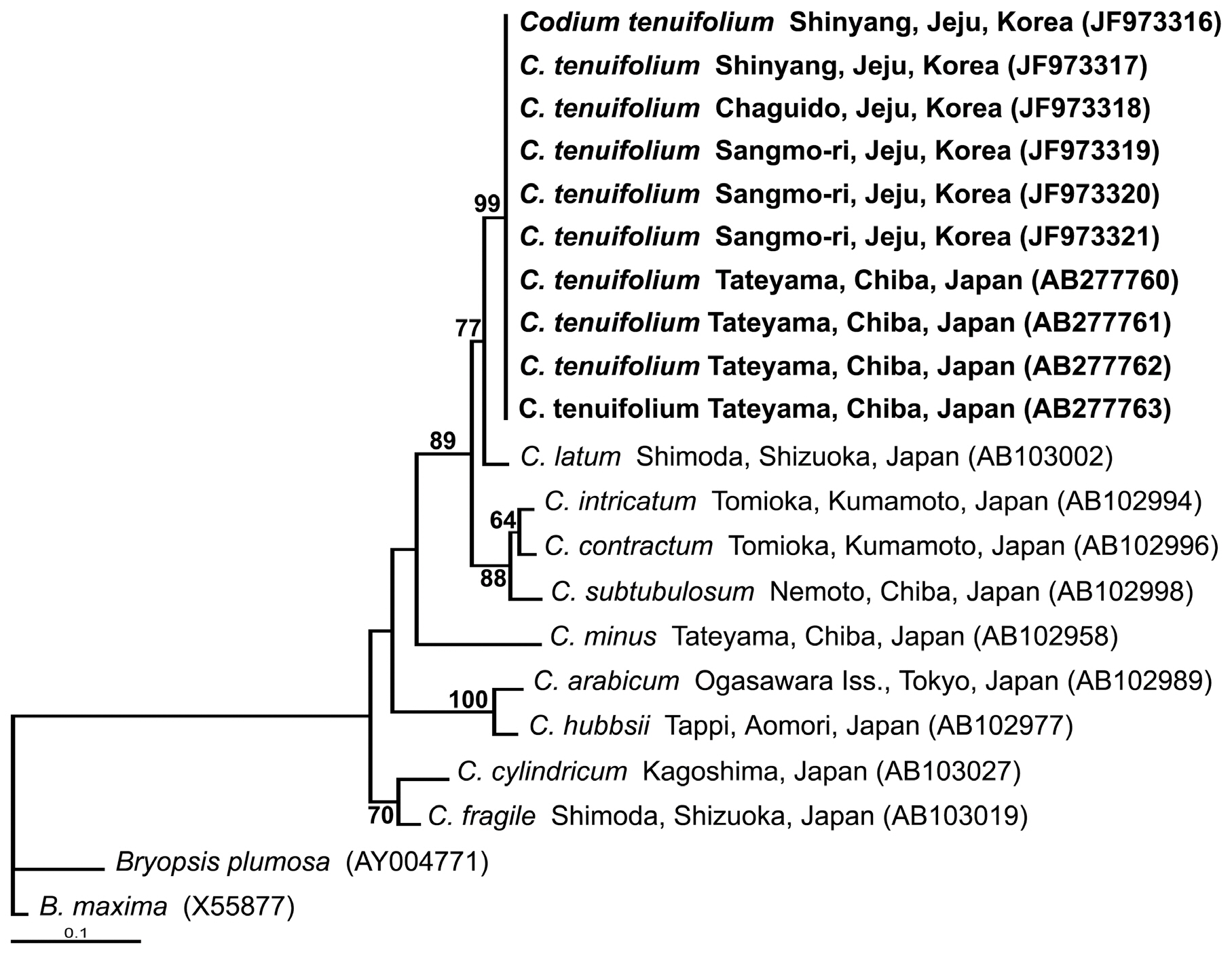
In the phylogenetic tree (Fig. 4), all carpet-like Codium specimens from Jeju and C. tenuifolium from Japan were associated on the same node. This clade had strong support (99%) and formed a sister group with C. latum from Japan with lower support (77%). The C. tenuifolium and C. latumclades were grouped with C. intricatum, C. contractum, and C. subtubulosum, with bootstrap values of 89% (Fig. 4).
Based on the analysis of rbcL gene and a comprehen sive morphological study, we confirmed the name of the carpet-like Codium species from Jeju as Codium tenuifolium Shimada, Tadano and J. Tanaka (2007), instead of C. tapetum Y. Lee or C. latum Suringar. C. tenuifolium has the following characteristics: (1) tenuous, flat and broadly expanded erect thallus with a fragile and velvety surface, (2) simple, bi- or trifurcate fronds, (3) sub-pyriform utricles with a swollen and tumid head, and (4) globular gametangia borne from a short pedicel attached to a utricle base. Although C. tenuifolium, as originally described (Shimada et al. 2007), has variable type gametangia, we observed only globular shaped gametangia. Phylogenetic analysis of rbcL gene sequences of carpet-like specimens from Jeju indicated that they were identical to C. tenuifolium from Japan. C. tenuifolium is distinct from Japanese C. latum and the two are sister species. C. tenuifolium occurs in the low intertidal and shallow subtidal zones on Jeju Island, with erect fronds individually attached to a rock or shell. In Korea, carpet-like Codium were previously identified as C. latum after Kang’s description (1968). Lee (2008) reported a new carpet-like species, C. tapetum Y. Lee. He suggested that the new species has been known in Korea as C. latum. Comparing morphological features of C. tapetum with C. tenuifolium, we confirmed that both species have the same morphological characteristics (Table 1). Thus, C. tapetum Y. Lee (2008) should be regarded as a synonym of C. tenuifolium Shimada, Tadano and J. Tanaka (2007).
According to Okamura’s description in 1915, C. latum has a broad thallus that is flat, expanded, and lanceolate or band-like externally. Because C. latum has similar morphological features to C. tenuifolium, most carpet-like Codium species had been recognized as C. latum in Korea. However, we identified two species based on our comprehensive morphological observations. The diagnostic features distinguishing the two species are the shapes of the utricles and gametangia. The utricles of C. latum are cylindrical and slightly enlarged at the top, forming a roundish knob with thin apical walls (Okamura 1915). In C. tenuifolium, utricles have a tumid head that is more swollen than the basal and middle sections. The gametangia of C. latum extend from the middle portion of the utricle (Okamura 1915, Schmidt 1923), but those of C. tenuifolium develop at the utricle base. C. latum gametangia are oblong or elongate-ovate (Okamura 1915, Schmidt 1923), but C. tenuifolium are variable and may
be globular, subglobular, or oval (Shimada et al. 2007, Lee 2008).
The molecular phylogenetic analysis using rbcL gene sequences demonstrated that the carpet-like specimens from Korea corresponded with C. tenuifolium from Japan and revealed sufficient sequence divergence to separate C. tenuifolium from other species. The sequence divergences of six C. tenuifolium samples were identical to each other, and they had 2.82% sequence divergence with C. latum from Japan. Although C. tenuifolium is superficially similar to C. latum in having a thin and broad thallus, rbcL sequence data identified a sister relationship between the two. Shimada et al. (2007) confirmed that C. tenuifolium specimen is separate from C. latum specimen through analysis of rbcL gene sequences with 2.6% pair-wise distance.
Silva (1954) characterized Codium with two subgenera: the subgenus Tylecodium has prostrate or erect thalli showing applanate, pulvinate, or compressed cylindrical fronds without true branching; the subgenus Schizocodium has erect thalli consisting of cylindrical or compressed fronds that are dichotomously or polytomously branched. Silva (1962) raised the subsection Lata, which was established by Schmidt (1923), to the section Lata for C. latum,C. latum subsp. palmeri, C. laminarioides, and C. platylobium, and these taxa have applanate, thin
and broad erect thalli. Verbruggen et al. (2007) studied the phylogenetic relationships of Codium using rbcL and rps3-rpl6 sequences, and demonstrated that C. latum formed a distant clade with C. laminarioides and C. platylobium. According to the results of our study, C. tenuifolium would belong to the section lata based on morphological features and a molecular analysis that C. tenuifolium is tied with C. latumas a sister taxon (Fig. 4). Therefore, it is necessary to demonstrate whether or not C. tenuifolium belongs in the section Lata by re-examining morphological features and DNA sequences.
In conclusion, we confirmed that the morphology and rbcL gene sequences of a carpet-like Codium from Jeju, Korea were identical to those of C. tenuifolium from Japan. We propose that carpet-like Codium from Jeju is C. tenuifolium, and that C. tapetum should be regarded as a synonym of C. tenuifolium. C. tenuifolium is not only clearly distinguished from C. latum based on morphological characteristics of the utricles and gametangia, but plastid rbcL gene sequences do provide sufficient pair-wise differences to separate the two species. Since it remains to be demonstrated whether or not C. latum occurs in Korea, more studies are required on the entity of C. latum from Korea.