Colonization patterns and community changes in benthic macroinvertebrates in the Cheonggye Stream, a functionally restored stream in downtown Seoul, Korea, were studied from November 2005 to November 2007. Benthic macroinvertebrates were quantitatively sampled 15 times from five sites in the stream section. Taxa richness (59 species in total)increased gradually over the first year, whereas the density revealed seasonal differences with significantly lower values in the winter season and after flood events. The benthic macroinvertebrate fauna may have drifted from the upstream reaches during floods and from the Han River, arrived aerially, or hitchhiked on artificially planted aquatic plants. Oligochaeta,Chironommidae, Psychodidae, and Hydropsychidae were identified as major community structure contributors in the stream. Swimmers and clingers colonized relatively earlier in the upper and middle reaches, whereas burrowers dominated particularly in the lower reaches. Collector-gatherers colonized at a relatively early period throughout the stream reaches, and collector-filterers, such as the net-spinning caddisfly (Cheumatopyche brevilineata), predominated in the upper and middle reaches after a 1-year time period. Cluster analyses and multi-response permutation procedures demonstrated that the Cheonggye Stream shares more similarities with the Jungnang Stream than with the Gapyeong Stream. Detrended correspondence analysis and nonmetric multidimensional scaling demonstrated that physical environmental factors (depth, current velocity, dissolved oxygen, and pH) as well as nutrients (total nitrogen and total phosphorous), water temperature, and conductivity could affect the distribution of benthic macroinvertebrates in the study streams.
Stream restoration has become a popular management strategy for improving the physical and ecological conditions of regulated urban streams, as streams and rivers are the most vulnerable ecosystems that humans impact (Allan and Castillo 2007, Bernhardt and Palmer 2007). Stream restoration projects in urban areas have increased in number and importance in recent years, due to growing awareness of conservation, management,and other public benefits, including recreational,aesthetic, and economic factors (Brown 2000). A number of studies have demonstrated that a loss of habitat diversity caused by channelization negatively influences the abundance and richness of stream biota, including fishes and macroinvertebrates. Therefore, considerable efforts have been devoted to restoring these degraded streams to conditions closer to their natural state (Malmqvist et al. 1991, Quinn et al. 1992, Bis et al. 2000, Brown 2000, Muotka et al. 2002, Purcell et al. 2002, Moerke et al. 2004, Alexander and Allan 2006, Nakano et al. 2008).
The Korean peninsula has experienced rapid environmental degradation beginning in the 1960s, at which time the Korean government launched a land development program (Bae et al. 2005). However, the land development trend slowed when the Korean government began promoting nature conservation and restoration in the 1990s. The “Cheonggye-cheon (Cheonggye Stream) restoration project” in downtown Seoul is a symbolic event in this regard. This project was planned as a city redevelopment program in an old city district in Seoul to provide clean water flow passing through the center of the city. However, this project triggered nation-wide biomonitoring and stream restoration efforts in Korea, and hundreds of urban or suburban streams are currently undergoing restoration procedures throughout Korea. Stream restoration is currently one of the most pressing social and political issues in Korea.
Benthic macroinvertebrates are good indicator organisms for biomonitoring water quality and instream environments of stream ecosystems (Rosenberg and Resh 1993). Numerous biomonitoring studies have been conducted in natural and regulated streams and rivers, but changes in the colonization and communities of benthic macroinvertebrates in restored urban streams have not been studied thoroughly thus far. Creating an artificial stream channel, as in the functionally restored Cheonggye Stream, has provided a unique opportunity in this regard.
The objective of this study was to assess the colonization patterns and community changes in benthic macroinvertebrates in the Cheonggye Stream with regard to chronological changes in community structure, functional groups, and indicator species or species groups, via a 2-year field investigation that was undertaken since the initial flow after construction.
>
Historical background of the Cheonggye-cheon restoration project
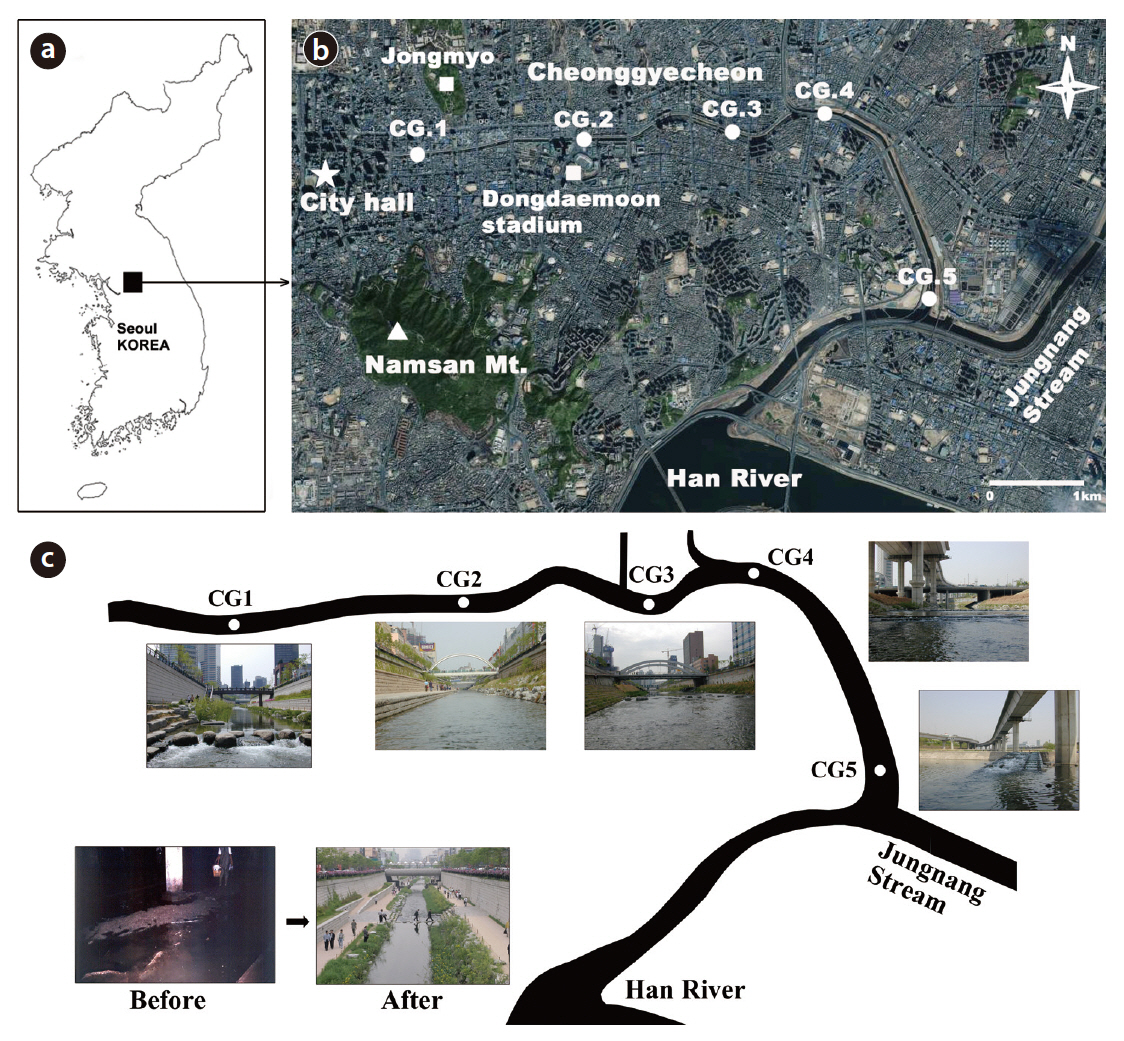
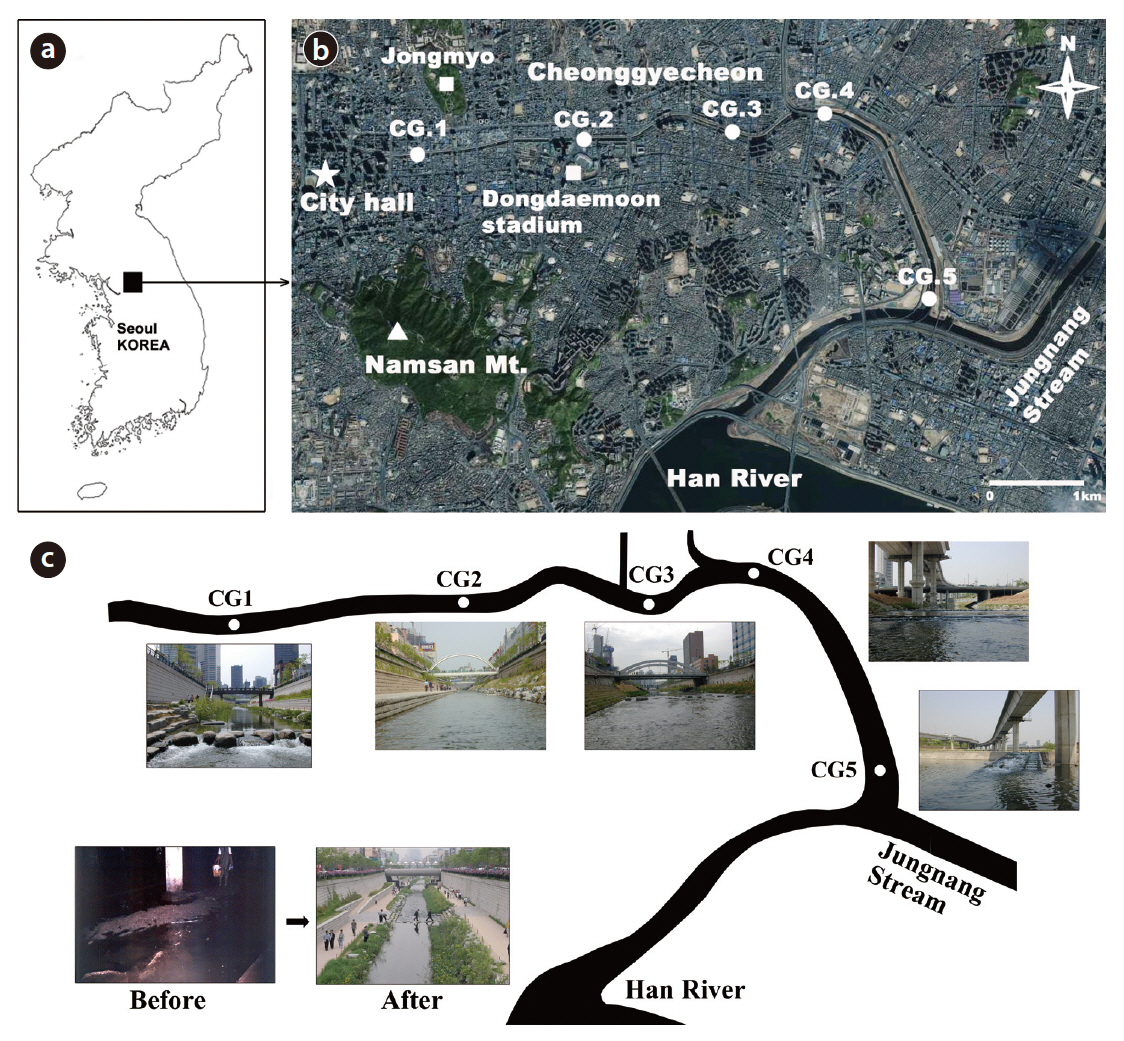
The Cheonggye Stream originates from the Inwangsan (Mt.) and Bukaksan (Mt.) areas located north of Seoul. The main stream follows the west-east axis through downtown Seoul and merges into the lower reach of the Jungnang Stream, which empties into the Han River (Fig. 1a and 1b). The total length of the Cheonggye Stream is approximately 11 km. The area includes urban structures and many office buildings.
The Cheonggye-cheon area is one of the most populated areas in Seoul, as this area has historically been a commercial and business center of the city since the beginning of the Joseon dynasty in 1392 (Figs. 1b and 2a). Between 1955 and 1977, an approximate 6-km stretch of the mainstream channel was cement paved from Gwangtong-gyo (bridge), near Seoul city hall, to Sindap-cheolgyo (a railroad bridge), approximately 6 km downstream from city hall (Fig. 2b). Additionally, a 5.6 km-long, 16-m wide elevated highway extending from Gwanggyo to Majang-dong, throughout the cement-paved section of the stream, was completed over the stream in August 1971, after 4 working years (Fig. 2c). In July 2003, Mr. Lee Myung-bak, then-Seoul mayor and current President of Korea, initiated a project to remove the cement-paved road and the elevated highway and to restore the stream. The stream was opened to the public on October 1, 2005 and was lauded as a major success (Fig. 2d).
Prior to the restoration effort, the main stream was covered completely by a road structure, with an elevated highway over the road, and an intercept sewage system under the road. During the 2-year restoration project conducted from July 2003 to September 2005 (budget, 386,000 million won), a 5.8-km section of the stream extending from Taepyeong-no, near city hall, to Sindap-cheolgyo was artificially reconstructed. As the Cheonggye Stream is an intermittent stream, and is normally dry, it requires additional flow to maintain a maximum 40-cm depth of water throughout the year. Approximately 120,000 t of water per day is supplied from the Han River (98,000 t) as well as from the subway groundwater system (22,000 t) to maintain the water level (http://cheonggye.seoul.go.kr).
The restored Cheonggye Stream consists of a series of pool-riffle sequences. These riffles are 10-20 m long, with a mean depth and current velocity (20 cm above the bottom) of 0.24 m (standard deviation [SD], ± 0.09) and 1.14 m/s (SD, ± 0.30), respectively. Riparian plants were artificially planted, but some trees, including
Five study sites along the restored section of the
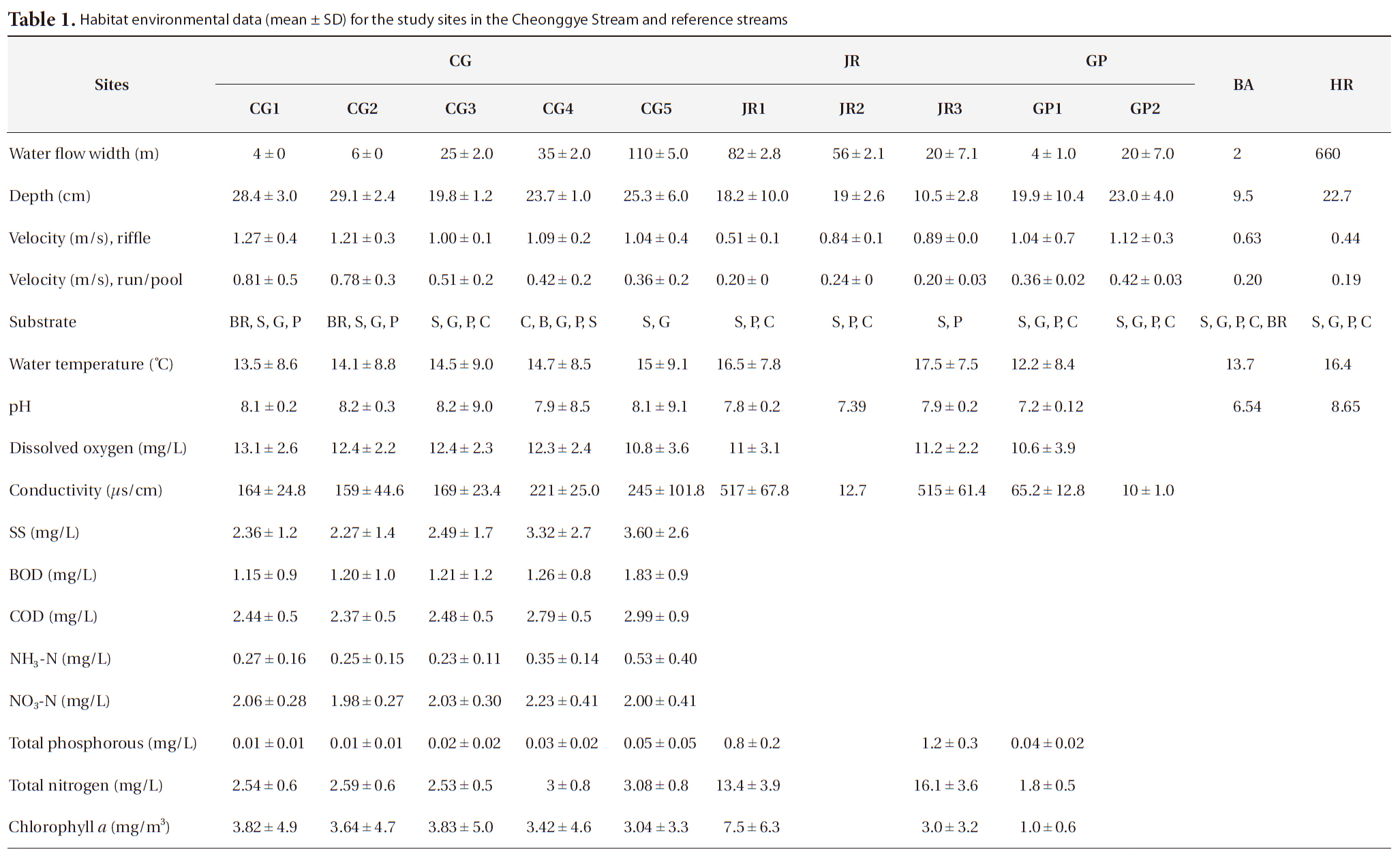
Cheonggye Stream, situated 1 km (Supyogyo, CG1: N 37°32´57.1˝, E 127°3´23.6˝), 2.8 km (Ogansugyo, CG2: N 37°32´57.1˝, E 127°3´23.6˝), 4.4 km (Biudanggyo, CG3: N 37°34´3.7˝, E 127°1´47.5˝), 5.5 km (Gosanjagyo, CG4: N 37°34´10.9˝, E 127°2´29.2˝), and 8 km (Jungnang influence, CG5: N 37°33´7.4˝, E 127°3´13.1˝) from the uppermost initial flow (hereafter referred to as CG1-CG5, respectively), were selected for sampling. The CG1-CG4 sites were located inside the past cement-paved section of the Cheonggye Stream. These study sites represented habitat types in the upstream (CG1 and CG2), mid-stream (CG3 and CG4), and downstream reaches (CG5) of the Cheonggye Stream (Fig. 1c).
We sampled two source streams, the Bukaksan (Mt.) Stream and the Han River to evaluate potential sources of benthic macroinvertebrate colonizers. The Bukaksan Stream is a headwater stream originating from the Bukaksan (alt. 343 m above sea level, located approximately 3 km north of the Cheonggye Stream) and connected to the main Cheonggye Stream. Most branch streams of the Cheonggye Stream inside the urban area of Seoul, including the Bukaksan Stream, are channelized with cement-paved road structures, and the water flows only when precipitation is sufficient (rainy season). The Han River site, located approximately 5 km upstream from the confluence of the Jungnang Stream, is the site to which the source water is supplied.
We also sampled two reference streams, the Jungnang Stream (three sites) and the Gapyeong Stream (two sites) to compare benthic macroinvertebrate communities. The Jungnang Stream is a nearby urban stream to which the Cheonggye Stream merges, and the Gapyeong Stream is a natural stream located approximately 60 km from Seoul. The sampling sites are as follows: Jungnang Stream (JR): JR1 (Nowon-gyo: N 37°32´57.1˝, E 127°3´23.6˝), JR2 (Ihwa-gyo: N 37°35´22.4˝, E 127°4´20.9˝), and JR3 (above Cheonggye Stream confluence: N 37°40´35.4˝, E 127°3´10.5˝); Gapyeong Stream (GP): GP1 (Seungcheon-sa: N 37°56´3.8˝, E 127°29´12.5˝) and GP2 (Seonbawi: N 37°56´13.6˝, E 127°29´21.8˝).
Field investigations were conducted from November 2005 to November 2007 at five study sites in the Cheonggye Stream. The first year’s (November 2005-November 2006) sampling was monthly, whereas sampling was conducted seasonally during the second year (February 2007, April 2007, August 2007, and November 2007). Reference streams (Jungnang and Gapyeong) were sampled in April 2006 and April 2007. The two source streams (Bukaksan and Han River) were sampled once in October 2006 to determine the origin of the Cheonggye Stream fauna.
Benthic macroinvertebrates were sampled quantitatively using a Surber sampler (50 × 50 cm; mesh 0.2 mm; 1 riffle, 1 run/pool, and 1 margin habitat sample per site; total sampling area 7,500 cm2 per site); the samples from three habitat types were pooled for statistical analyses. Marginal sampling was taken within a 1-m area from the bank at which the flow was very slow, and aquatic macrophytes were present. However, these marginal sampling data were not used for comparisons with reference stream sites because the bank structures of the artificial and natural streams were fundamentally different. The samples were preserved with Kahle’s solution and transported to the laboratory for sorting and identification. Benthic macroinvertebrates were identified to the species level using available references (McCafferty 1981, Wiederholm 1983, Yoon 1995, Merritt and Cummins 1996, Kawai and Tanida 2005). Other sampling and habitat investigation methods were conducted in accordance with general protocols (Ward 1992, Williams and Feltmate 1992, Merritt and Cummins 1996, Allan and Castillo 2007). Benthic macroinvertebrates were assigned to a habitat orientation group and a functional feeding group (Merritt and Cummins 1996).
Benthic macroinvertebrate communities were analyzed using quantitative sampling data in relation to the environmental data. We initially evaluated responses of macroinvertebrate taxa and groups using a site (block) in time approach and analyses of variance (ANOVA) with the SAS program (Sokal and Rohlf 1994). Logarithmic transformations were applied to satisfy the ANOVA assumptions. ANOVA was used to test for differences in mean abundance, mean species richness, and mean species diversity in the macroinvertebrate community (SAS Institute 2001). An ANOVA was used to compare the chronological changes and distributional differences in macroinvertebrate communities (Smith and Smith 2001).
The averaged data were examined for the multivariate analyses with PCORD ver. 4.28 (McCune and Grace 2002). The main matrix for each sample had high beta diversity, moderate-to-extreme row and column skewness, and a high coefficient of variation among the sums of the tricolumns(species) in the matrix. We deleted rare species that occurred in less than 5% of the samples and applied a logarithmic transformation.
We used detrended correspondence analysis (DCA) and nonmetric multidimensional scaling (NMS). DCA was used to determine the number of factors structuring the complex macroinvertebrate community and to qualitatively summarize the overall distribution of species assemblages across the gradients of different areas (Hill and Gauch 1980). Sorensen’s distance measure was used in species space. NMS (Kruskal 1964, Mather 1976, Clarke 1993) is an iterative method based on rank distances between sample units. It is useful for ecological gradient studies because of its general robustness and lack of assumptions about the distribution or type of data. Therefore, NMS was used to determine the number of factors in the complex macroinvertbrate community structure and to qualitatively summarize the overall distribution of species assemblages across the gradients of Cheonggye Stream changes. We examined 13 variables to identify which had the highest correlation coefficients. The variables used included two categorical variables (date and site) and 11 quantitative variables (water flow width, depth, velocity of riffle, velocity of run/pool, water temperature, pH, dissolved oxygen [DO], conductivity, total phosphorous, total nitrogen, and chlorophyll
Simple repeated-measure designs, which are common in ecology, require a multiresponse permutation procedure (MRPP) (McCune and Grace 2002). The MRPP analysis assigned the observed values randomly to different sites (blocks). We could use MRPP because our research fit the analysis of block differences well. This procedure is useful for analyzing ecological data because it does not require assumptions of normality and constant variance (Biondini et al. 1988, McCune and Mefford 1999).
Cluster analyses were conducted using presence/absence data from the five sites in the Cheonggye Stream (quantitative data) and from the reference sites separately for April 2006 and April 2007. Normalized Euclidian distances were computed for the most distant site pairs (complete linkage) (Wilkinson 1988).
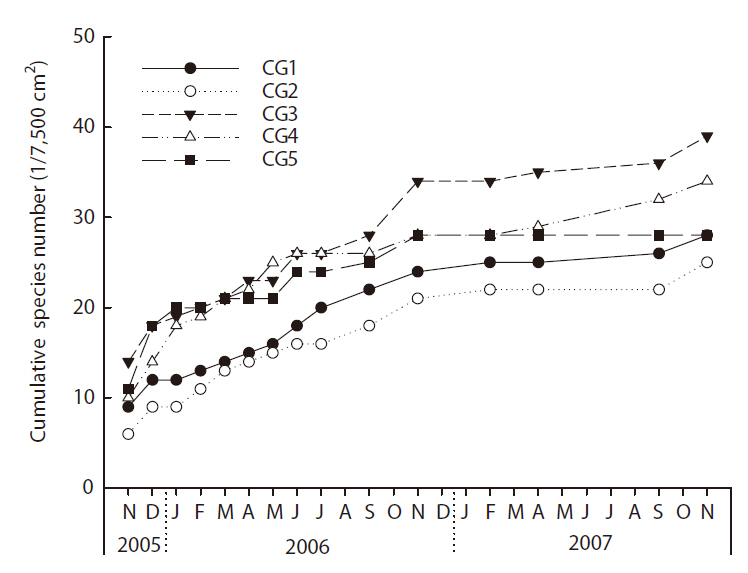
The general habitat environments at the study sites are shown in Table 1. As the Cheonggye Stream was constructed in the form of a straightened channel with assembled bedrock pieces and an artificial water supply, water flow width, depth, and velocity remained almost constant throughout the year. However, during sudden heavy rainfall (approximately > 100 mm per h), the stream water overflowed over the walkways beside the stream banks in the upper and mid-stream reaches immediately after a rainfall. In general, the upper stream reach (CG1 and CG2) was relatively steeper and narrower, with a faster current velocity than the lower stream reach. Yearly average water temperature was relatively lower and DO levels were relatively higher in the upper sampling sites, but the differences between the sites were not so profound. Conductivity, nutrient (total nitrogen and total phosphorus), and pH levels were somewhat higher than that observed in other natural reference streams. Attached algae (
>
Benthic macroinvertebrate faunas and communities
Fifty-nine species of benthic macroinvertebrates were sampled from the Cheonggye Stream study sites over the study period (Appendix 1). In the reference streams, 10 and 66 species were detected in the Jungnang and Gapyeong streams, respectively. Twenty-two species were detected from the Bukaksan Stream and the Han River, from which the benthic macroinvertebrate fauna may have originated.
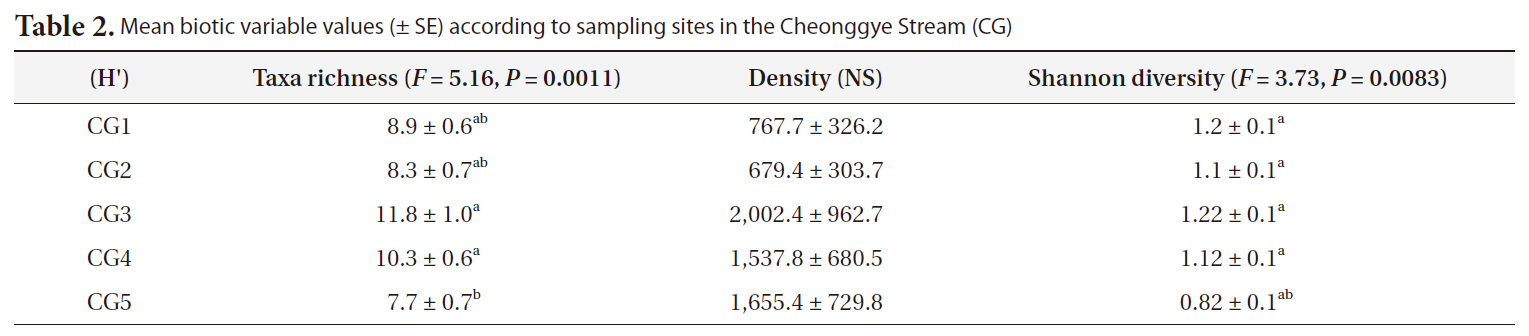
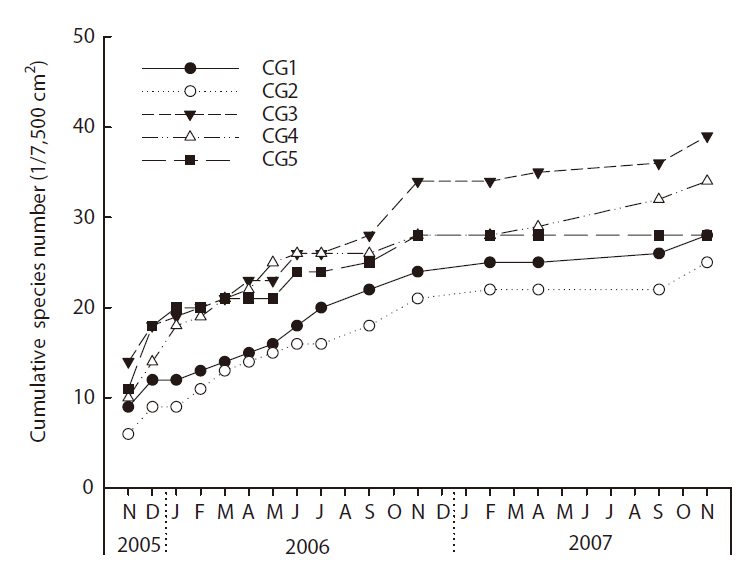
The cumulative species number at the study sites, and at the middle and lower stream sites (CG3-CG5) in particular, increased very rapidly during the initial 1-month period, but the rate of increase gradually slowed over the following years (Fig. 3). Larger numbers of benthic macroinvertebrate species were detected in the middle stream sites (CG3 and CG4) than the upper (CG1 and CG2) and lower study sites (CG5).
Taxa richness differed significantly among the five Cheonggye Stream sites (
Taxa richness and density following 15 samping tri-
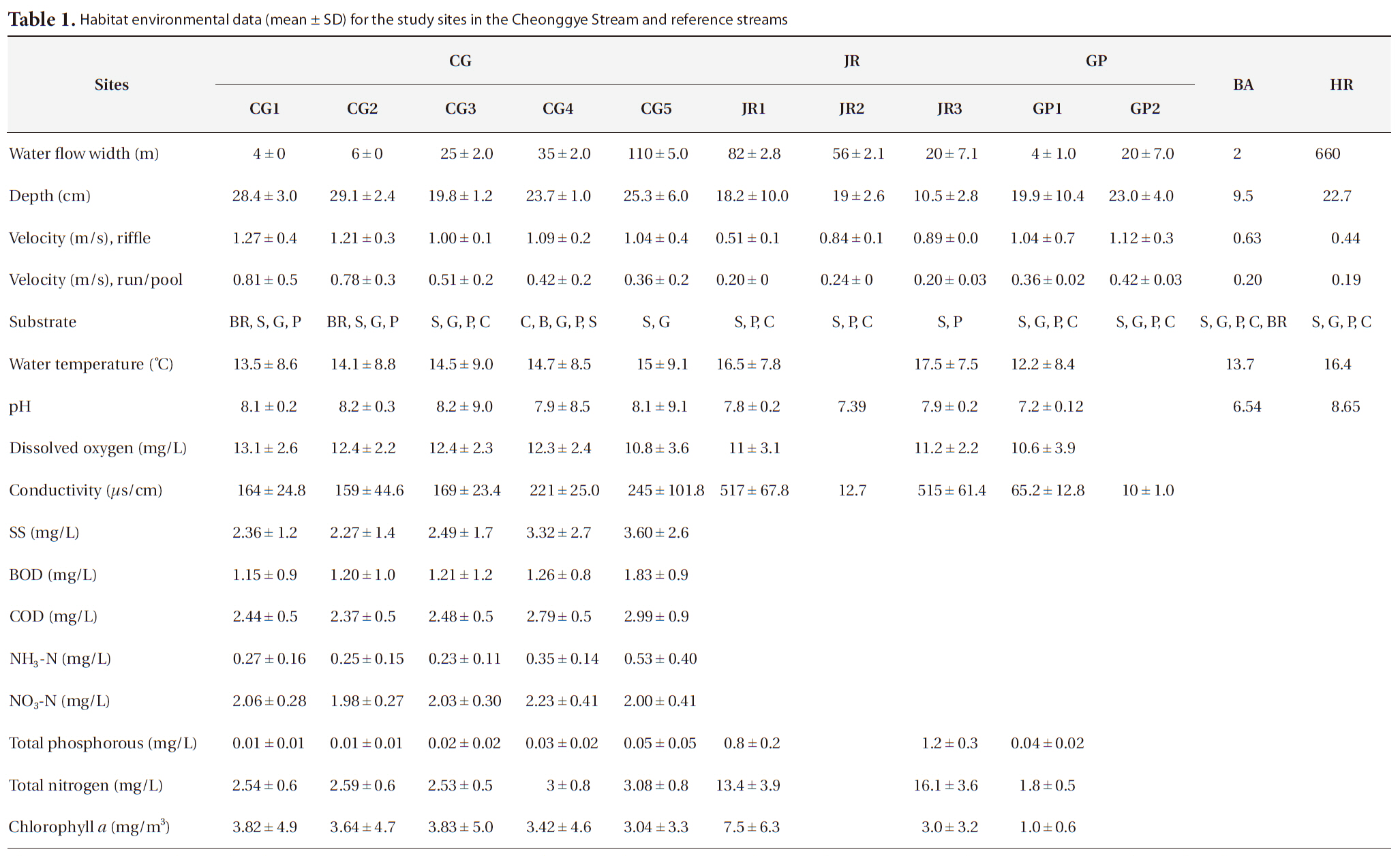
Habitat environmental data (mean ± SD) for the study sites in the Cheonggye Stream and reference streams
als over 2 years (November 2005-November 2007) at the five Cheonggye Stream sites differed significantly (
>
Colonization patterns and community changes
At the first sampling trial in November 2005, aquatic
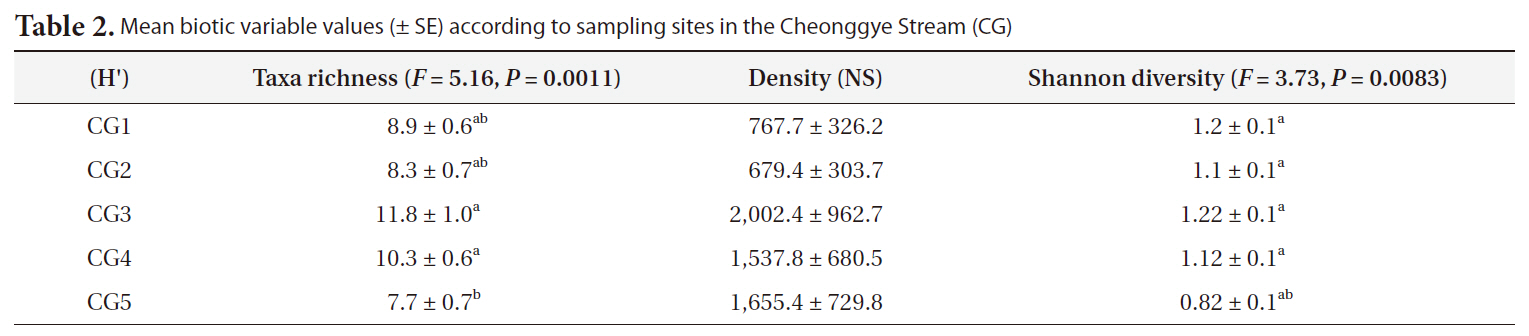
Mean biotic variable values (± SE) according to sampling sites in the Cheonggye Stream (CG)
insect taxa such as the Ephemeroptera, Trichoptera, Chironomidae, and Odonata colonized rapidly, and Plecoptera, Hemiptera, and Coleoptera occurred subsequently over the first 6-month period. Other benthic macroinvertebrates, including Oligochaeta, Gastropoda, Hirudinea, and Turbellaria were also detected during the initial time period. A large number of Chironomidae, Oligochaeta, and Baetidae (57.6%, 26.9%, and 9.1%, respectively) occurred throughout the study period, whereas other aquatic insects and benthic macroinvertebrate taxa, including Hemiptera, Plecoptera, Odonata, Coleoptera, Gastropoda, Hirudinea, and Turbellaria remained at relatively lower abundance throughout the entire study period.
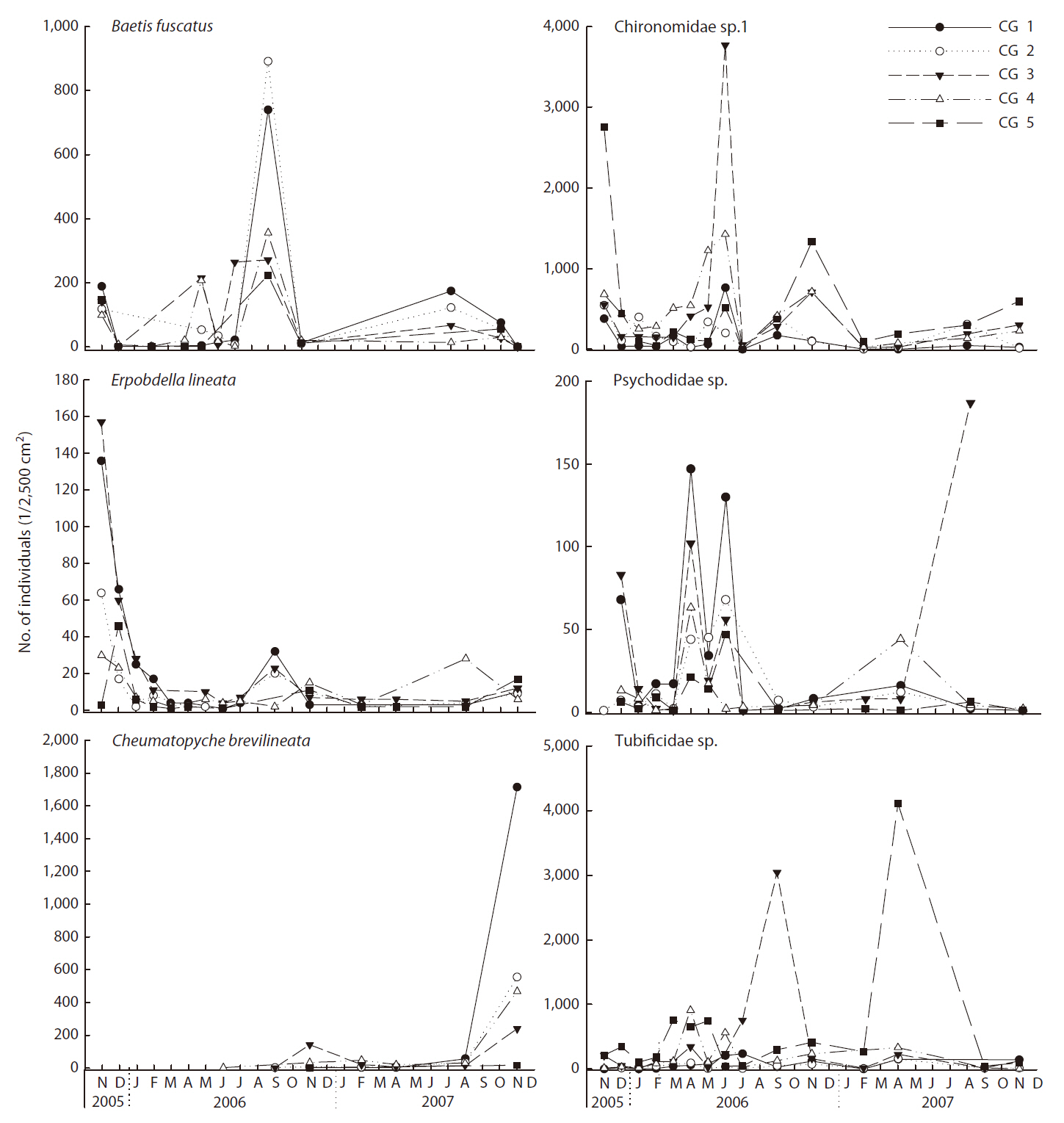
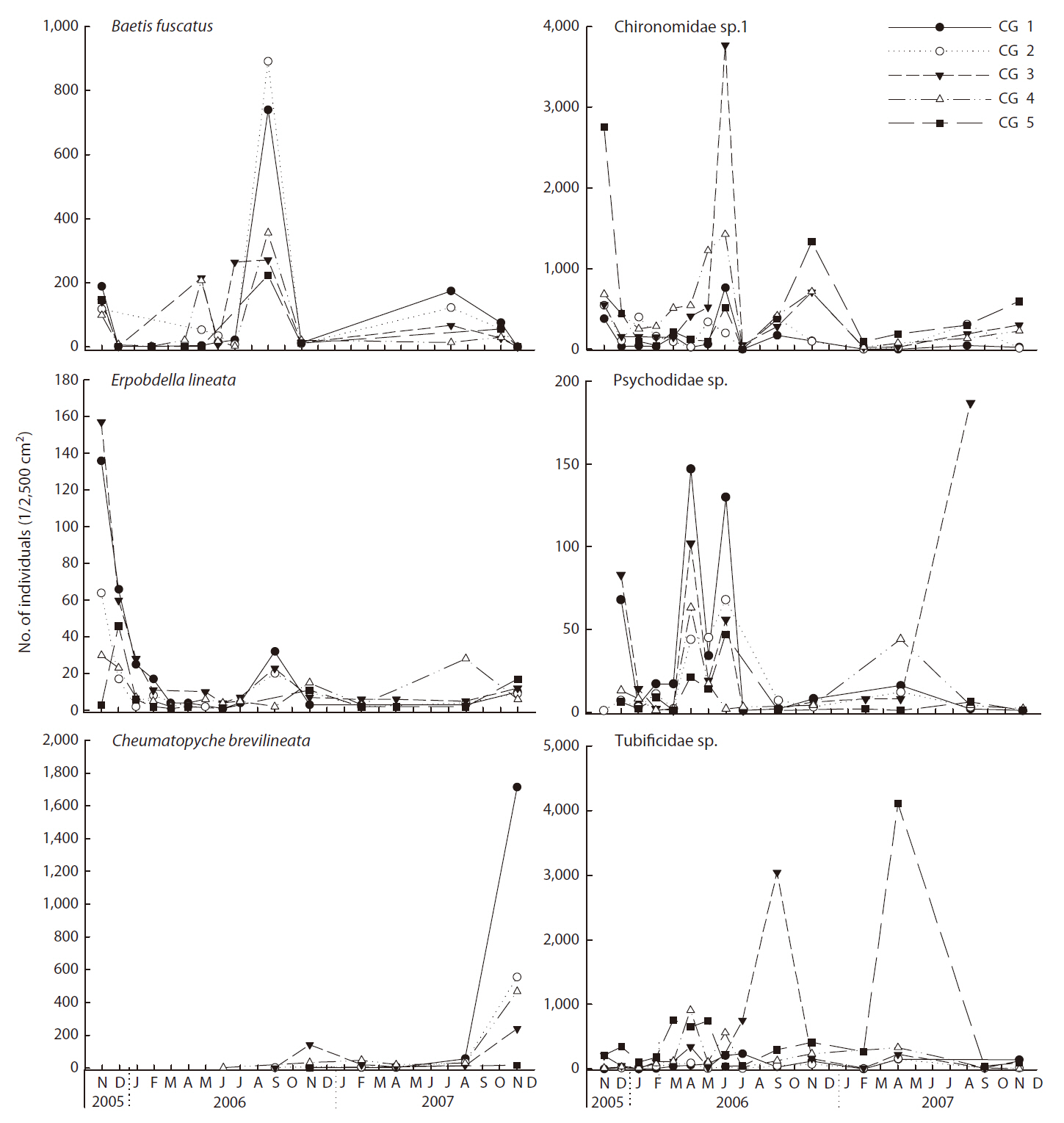
Dominant species in the Cheonggye Stream differed depending on site and time (Fig. 5). A chironimid species (Chironominae sp. 1) and a tubificid worm (Tubificidae sp.) occurred abundantly throughout the study sites, but
they were relatively more abundant in the lower stream sites and emerged in mass at certain sampling trials (e.g., November 2005 and November 2006 at CG 5 and June 2006 at CG3 for Chironominae sp. 1; April 2006 at CG3 and April 2007 at CG5 for Tubificidae sp.). A baetid mayfly (B. fuscatus) occurred more abundantly in the upper stream sites (CG1 and CG2) and peaked during the spring and summer. A psychodid fly (Pshchodidae sp.) occurred more abundantly during the spring. A leech species (
val from the initial flow.
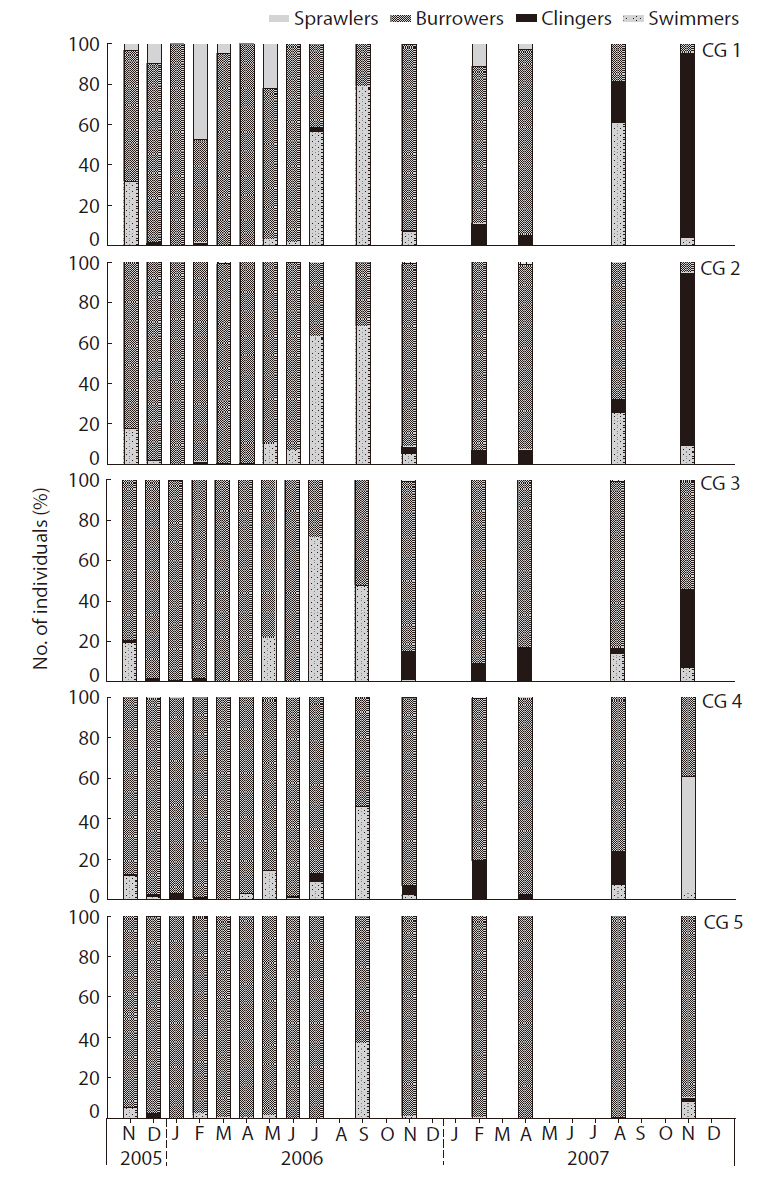
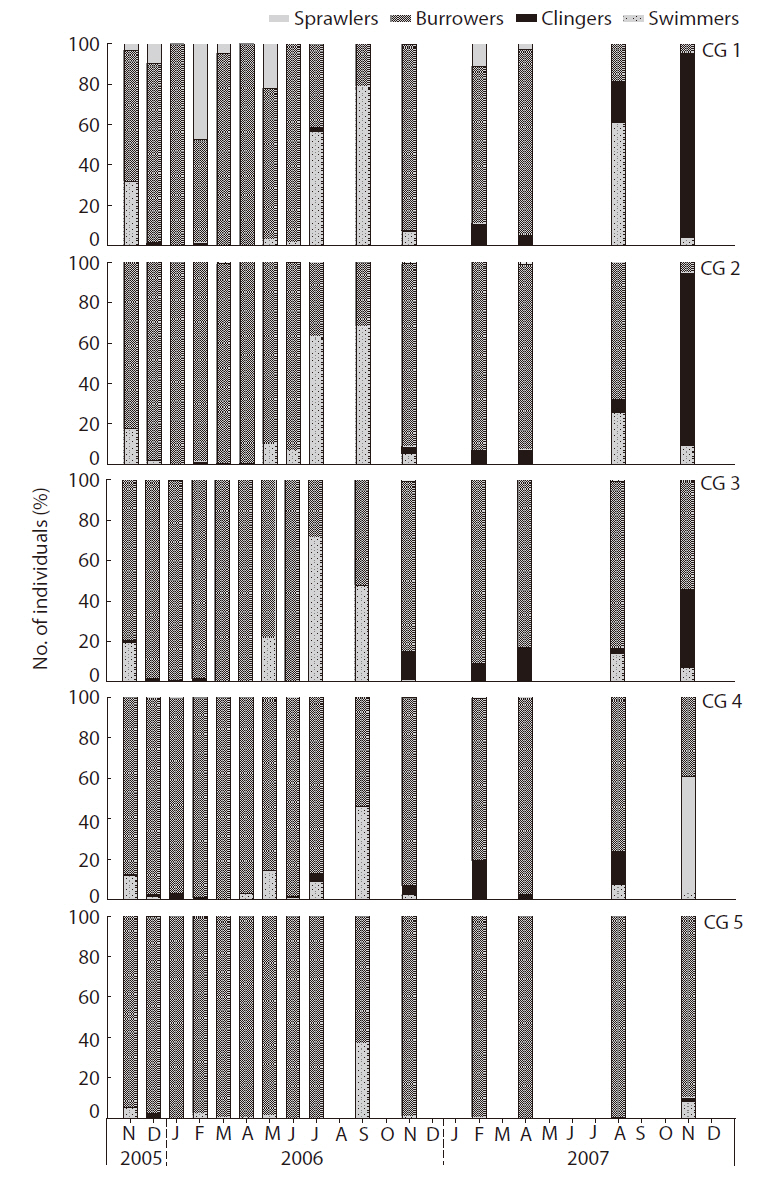
The habitat orientation groups changed with changes in density of the major benthic macroinvertebrate taxa, in particular the dominant species (Fig. 6). The swimmers (e.g.,
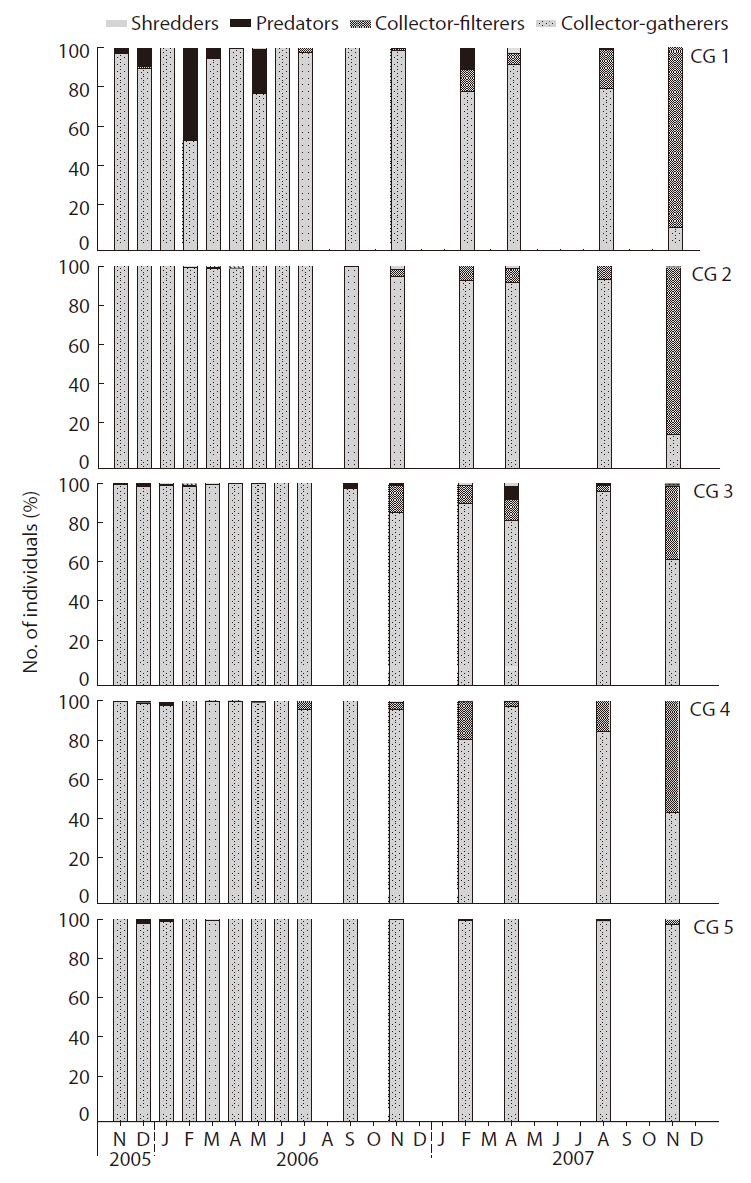
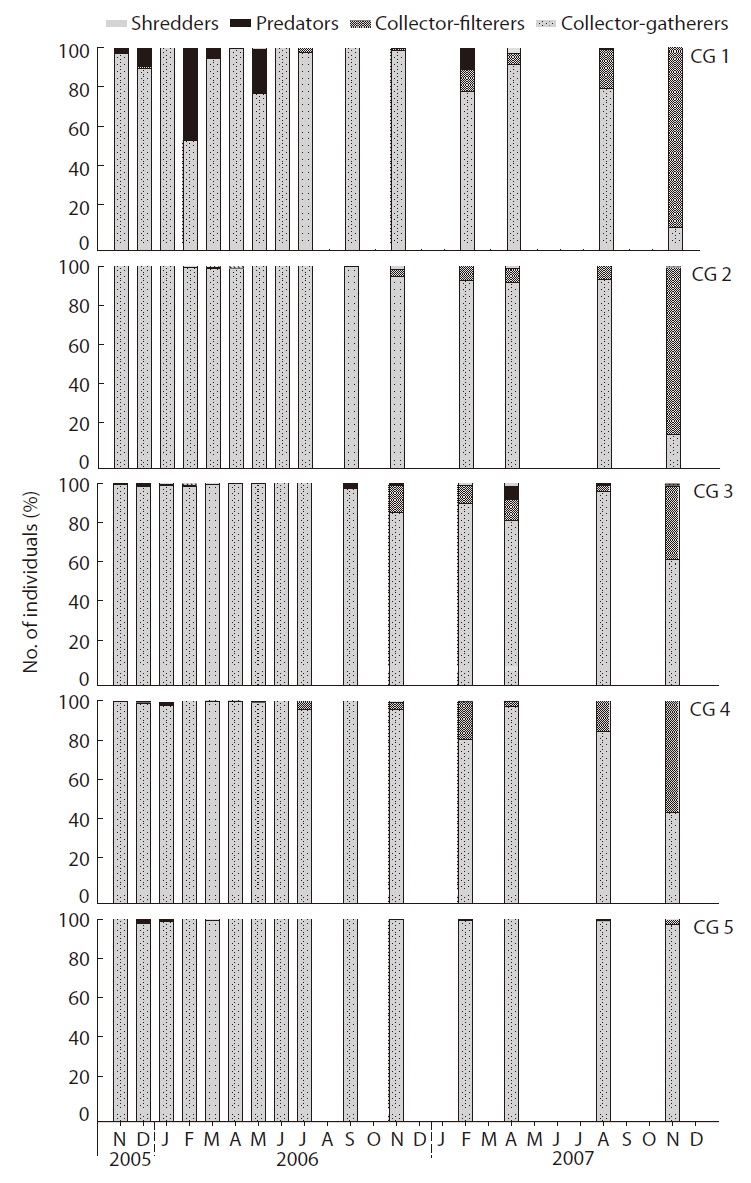
Collector-gatherers and collector-filterers colonized first, followed by the scrapers, predators, and shredders (Fig. 7). Collector-gatherers (Chironominae sp. 1, Tubificidae sp., and
were quite scarce at all stream sites throughout the study period.
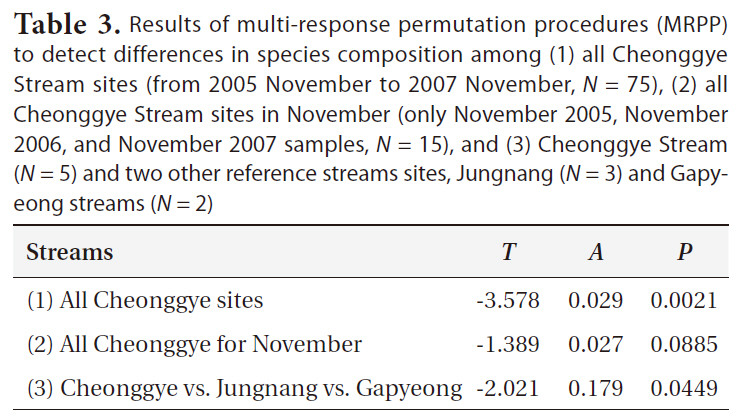
The MRPP confirmed that the benthic macroinvertebrate communities of all Cheonggye Stream sites differed significantly (
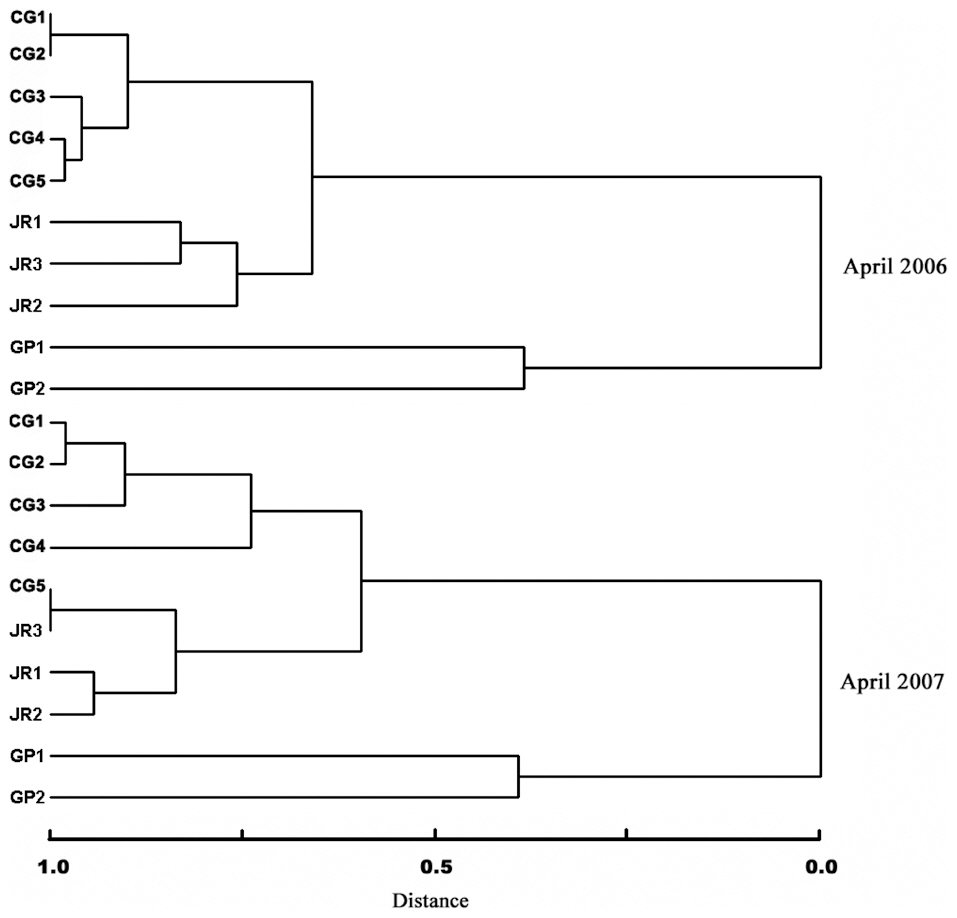
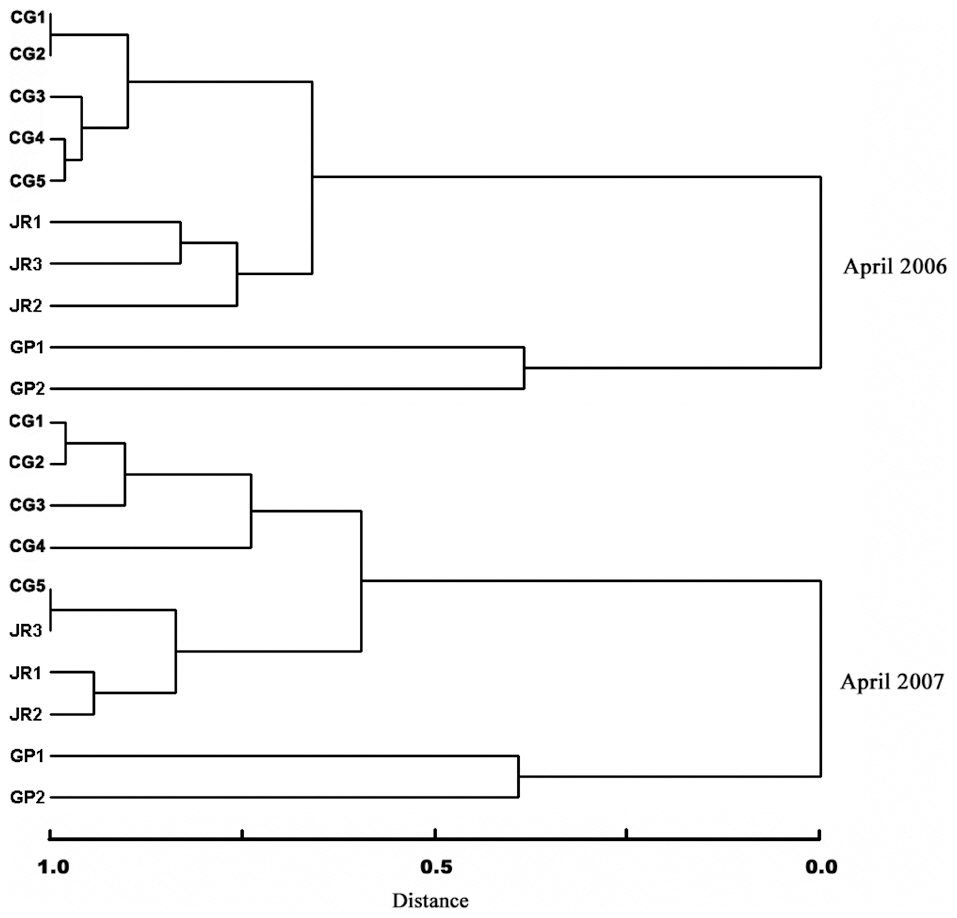
The species composition cluster analyses for the first year (April 2006) also demonstrated that the benthic macroinvertebrate communities between the sites of the Cheonggye Stream (CG1-CG5) were clustered more closely than between those of the Jungnang Stream (JR1-JR3) and Gapyeong Stream (GP1-GP2) (Fig. 8). The cluster analysis results for the second year (April 2007) were similar to those of the first year, except that the lower site of the Cheonggye Stream (CG5) was more closely clustered with the lower site of the Jungnang Stream (JR3).
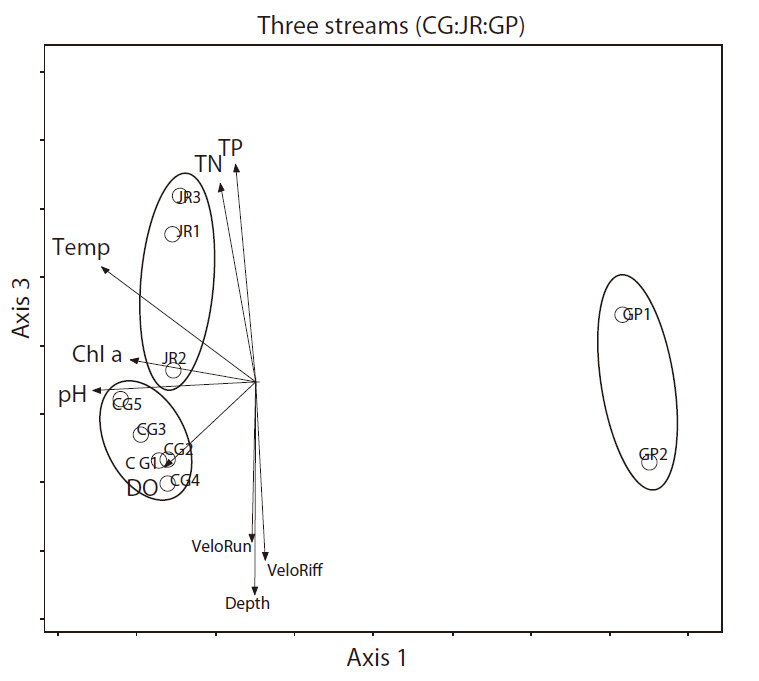
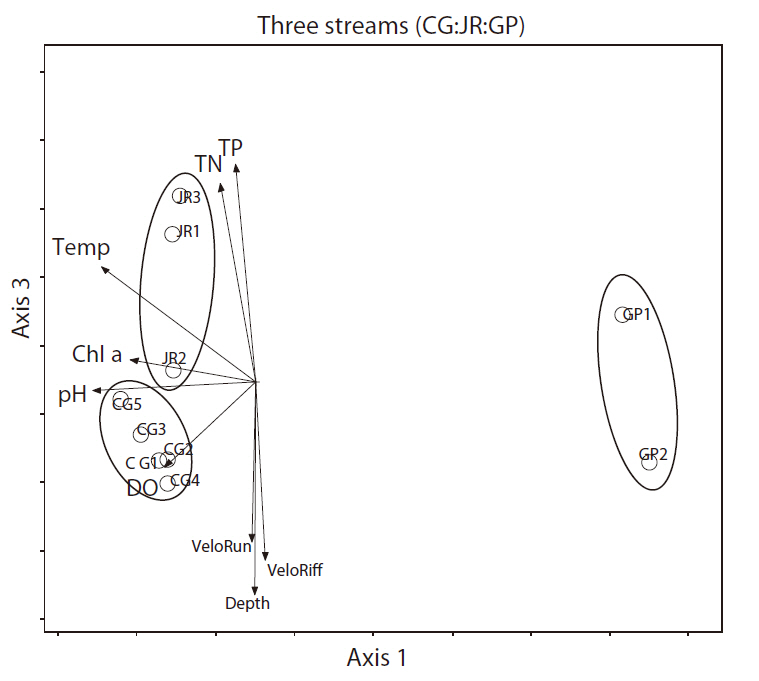
We analyzed all three streams and all seasons with DCA to confirm the results of the cluster analysis. The first two axes of the DCA ordination accounted for the majority of the variation in the species data, with eigenvalues of 0.72 and 0.18 for axes 1 and 3, respectively, and a total inertia (total variation in the species data ) of 3.197. The macroinvertebrate communities of the Cheonggye and Jungnang streams were separated from those of the Gapyeong Stream, but not clearly separated by those from the Cheonggye and Jungnang streams (Fig. 9). We examined 11 quantitative environmental variables in the DCA, and our results showed that axis 1 was negatively correlated with water temperature (
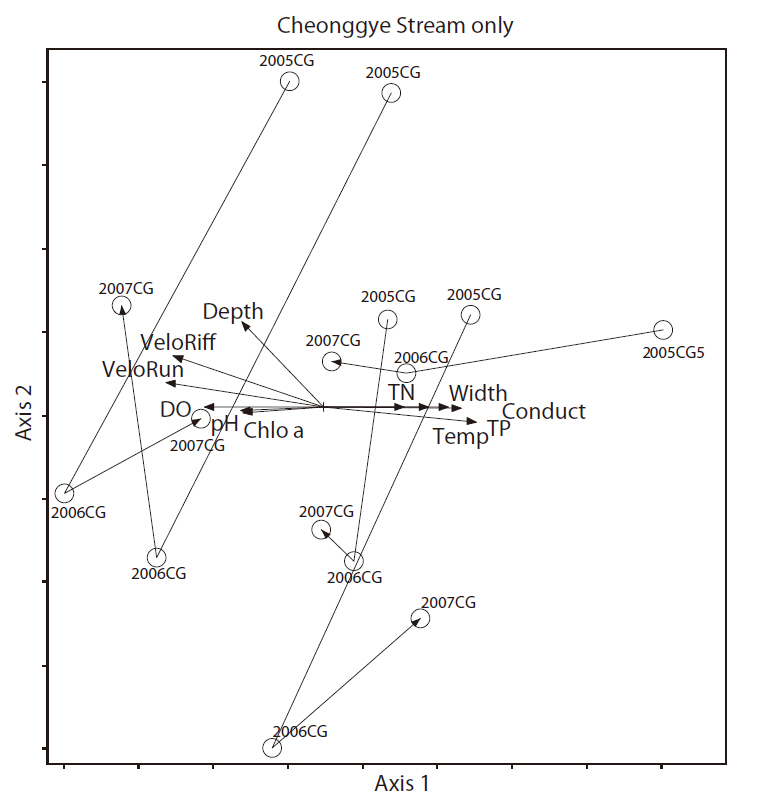
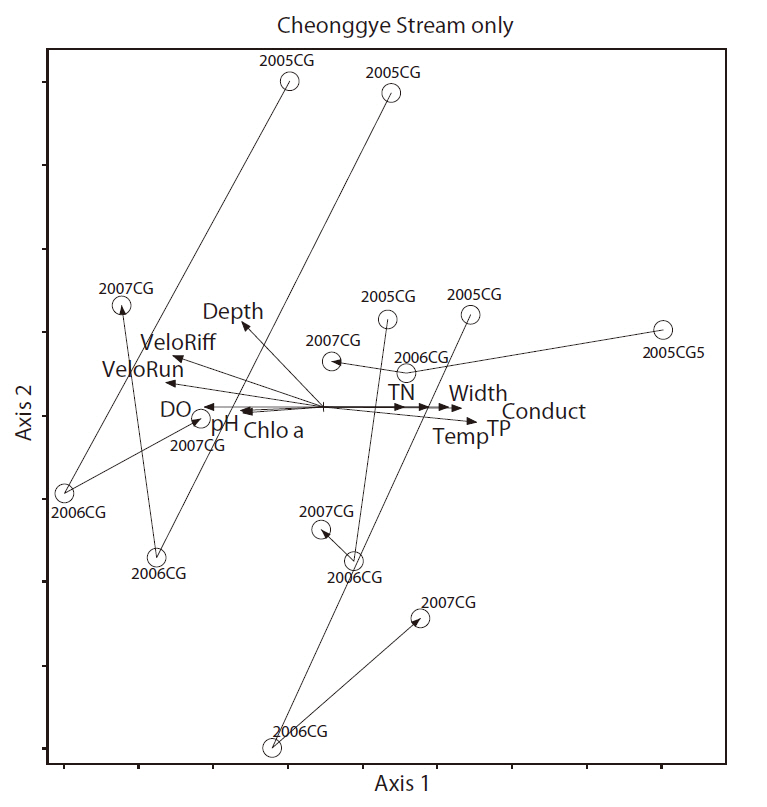
We only used the Cheonggye Stream November samples for 3 years (2005, 2006, and 2007) to determine the annual changes in macroinvertebrates. Then, we examined the 11 quantitative environmental variables with the Cheonggye Stream macroinvertbrate community using GapyNMS(Fig. 10). The results of the NMS ordination showed separation among the Cheonggye Stream communities; axis 1 and axis 2 explained 14% and 57% of the variance between sampling points, respectively. Axis 1 was positively correlated with water flow width (
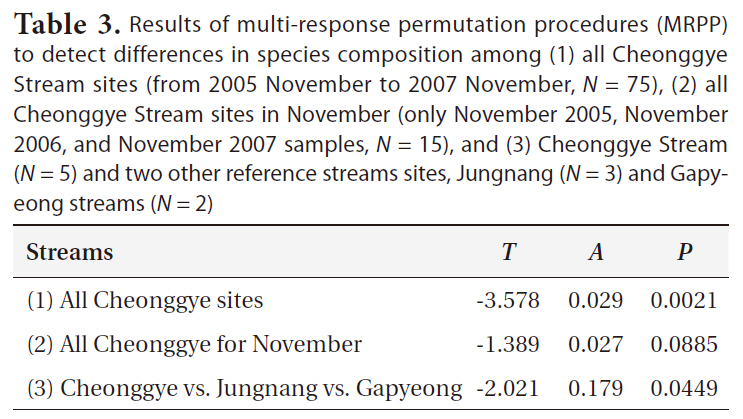
Results of multi-response permutation procedures (MRPP) to detect differences in species composition among (1) all Cheonggye Stream sites (from 2005 November to 2007 November N = 75) (2) all Cheonggye Stream sites in November (only November 2005 November 2006 and November 2007 samples N = 15) and (3) Cheonggye Stream (N = 5) and two other reference streams sites Jungnang (N = 3) and Gapyeong streams (N = 2)
axis 1 was negatively correlated with depth (
Macroinvertebrate colonization in new stream reaches occurs via drift, aerial sources, hyporrheic sources, and upstream migration (Mackay 1992). The benthic macroinvertebrate fauna in the newly-built section of the Cheonggye Stream may have originated from a drift from upstream reaches (e.g., the Bukaksan Stream) during floods or from the Han River, the major origin of the water source. Possible migrants from the headwater streams include mountain stream-dwelling mayflies [e.g.,
However, the species occurring very abundantly throughout the restored stream section (e.g., Chironominae spp., B.
Larvae of dragonflies (
Among the aquatic insects, the Baetidae (mostly
We detected an early dominance of collector-gatherers, including Baetidae and Chironomidae. This predominance of collector-gatherers (e.g., B.
Although benthic macroinvertebrates in the Cheonggye Stream colonized rapidly, species richness was maintained at relatively low levels compared with other natural streams (Appendix 1). Other colonization studies conducted in new stream channels have shown that macroinvertebrates require 70 to 125 days to reach a similar level of density and community similarity to an upstream reach (Gore 1982, Mackay 1992). Cumulative species richness reached a maximum approximately 700 days after the opening of the new channel (Minshall et al. 1983, Mackay 1992). During a 2-year study period in the restored Cheonggye Stream, the density, species richness, and community structure changed, depending on season, location, and other unpredicted environmental factors, including floods (Table 3). Other anthropogenic effects, such as increases in tourism, stream bed cleaning for filamentous algae, etc., may have enhanced such unstable conditions.
In a survey conducted in the Cheonggye Stream prior to the restoration project in 1986 (as in Fig. 1c: before), only a few species of benthic macroinvertebrates, e.g., tubificid oligochaetes, psychodid fly larvae, and chironomid larvae, were detected inside the stream tunnel (Yoon et al. 1986). These organisms were considered very tolerant indicator species and are found only in very polluted water bodies, such as sewage channels (Rosenberg and Resh 1993). Considering this observation, changes in benthic macroinvertebrate communities after restoration projects can be substantial. However, compared with other urban streams (Jungnang Stream) and natural streams (Gapyeong Stream), the Cheonggye Stream currently shares more similarities with urban streams in terms of benthic macroinvertebrate community structure (Figs. 8 and 9). Diverse biotic and abiotic environmental factors could affect the structuring of the benthic macroinvertebrate communities in Cheonggye Stream and other reference streams. Physical environmental factors such as depth, current velocity, DO, and pH, as well as nutrient (total nitrogen and total phosphorous), water temperature, and conductivity factors could affect the distribution of benthic macroinvertebrates in the study streams, as shown in Figs. 9 and 10. These findings indicate that the restored Cheonggye Stream is a relatively unstable dynamic system compared to reference urban or natural streams.