As gradual phenomena progressed in large spatial scale, the spans of research into climate and its changes can include the climatology, geography, and evolutionary biology. Ongoing climate change is regarded as a driving factor for ecosystem changes in the 21st century(Intergovernmental Panel on Climate Change 2001). Effects on the dynamics of biotic communities have been frequently associated with extreme weather events on ecological time scales (Easterling et al. 2000), and with climatic extremes at evolutionary time scales (Gutschick and VassiriRad 2003). Since the middle of the 20th century, average global forest net primary production has apparently increased, largely because of various combinations of increasing temperature, precipitation, cloudless days, atmospheric CO2, and nutrient deposition (Boisvenue and Running 2006). Regional exceptions also exist, because compensatory increases of precipitation have not been homogeneous (Jump et al. 2006). Such observations underscore the necessity for long-term monitoring programs to improve our understanding of environmentally-driven changes in ecosystems.
Forest (or community) development, maturity, and monograph are complex processes which occur in spans of decades to hundreds of years. Even ecologists have yet to clarify the precise processes and patterns relevant to the establishment and development of forests. In temperate regions, the vegetation responses to gradual climate change and extreme weather events have been observed in mature and old growth forests (e.g., van Mantgem and Stephenson 2007, van Mantgem et al. 2009). In young forests, during the maturational process, the majority of changes in structure, composition, and demographics are attributed not so profoundly to exogenous processes such as climate change and pathogen, but rather largely to endogenous processes such as competition and disturbance exclusion (e.g., fire).
In Korea, forest land, as emerging landscape, is in a developmental stage after severe disturbances, the Korean War, heavy utilization, and fire; thus, most Korean forest land is less than 60 years old. Therefore, long-term ecological research involving the installation of permanent
[Table 1.] Geographical locations and physical descriptions of KNLTER sites
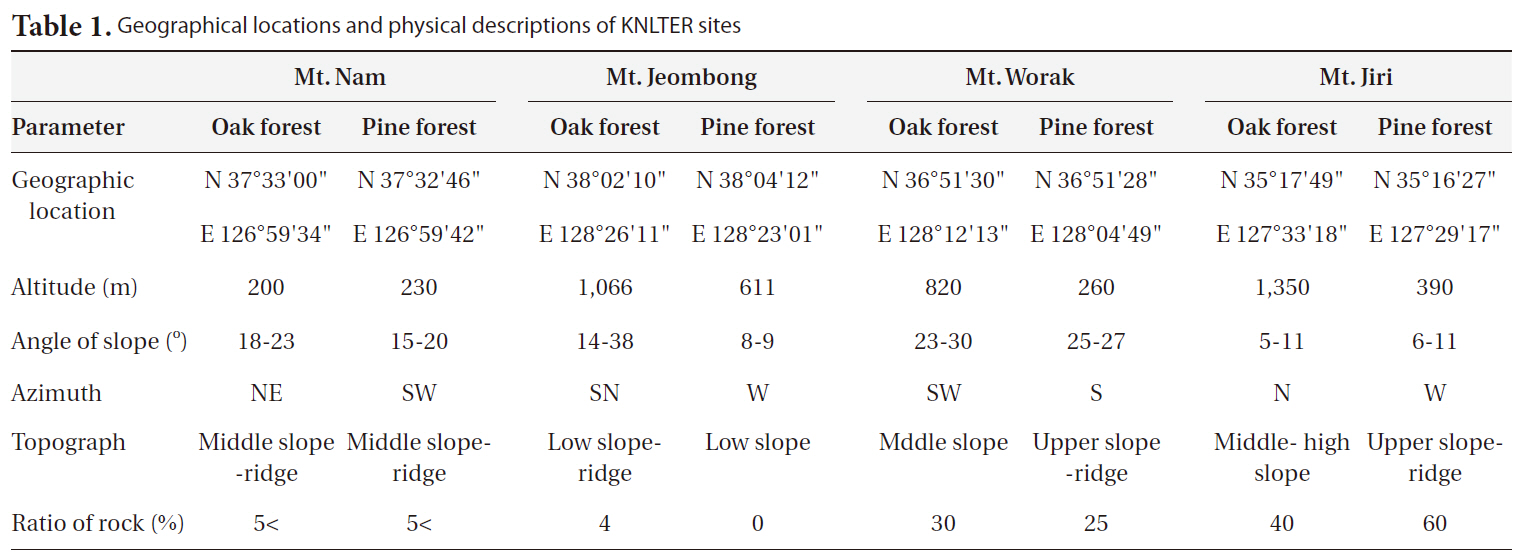
Geographical locations and physical descriptions of KNLTER sites
plots and meteorological stations can be employed not only to evaluate climate-related responses, but also to clarify the processes and patterns inherent to community development. Based on such long term observations and acquired information, sustainable and adaptive natural resource management can be achieved.
We analyzed changes in community structure (breast height area and stem density), composition, diversity (richness), and demographics (mortality and recruitment rates) in four Korea National Long Term Ecological Research (KNLTER) sites: Mt. Nam (Mt. N), Mt. Jeombong (Mt. Je), Mt. Worak (Mt. W), and Mt. Jiri (Mt. Ji). Herein, we have focused on the changes in vegetation attributes, because meteorological sensors are not installed at all LTER sites. We concluded by discussing emerging research challenges (feedbacks) for KNLTER.
Four KNLTER sites-Mt. N, Mt. Je, Mt. W, and Mt. Ji-were analyzed in this study. The geographical location and physical environments of each site are shown in Table 1. Four mountains and three forest types (
We analyzed the changes in community structure (basal area and stem density), species composition (by detrended correspondence analysis [DCA]), richness, and mortality and recruitment rates. For ordination analysis, the matrix of importance values for each species was established by the sum of relative basal area (BA) and density, and fed to the DCA ordination using PC-ORD ver. 4.0 (McCune and Mefford 1999). Mortality rate (for 5 years) was calculated by dividing the number of dead trees for 5 years by the number of living trees in the previous measurement. The recruitment rates (for 5 years) were obtained by determining the ratio of the number of trees added at the current measurement to the number of living trees in the previous measurement. Notably, our results were not expressed on an annual basis demograph. Finally, we conducted regression analyses between mortality and recruitment rate and between the rate of change in basal area and stem density. SPSS ver. 15.0 (SPSS Inc., Chicago, IL, USA) was applied for analyses.
>
Changes of basal area and stem density
Most of the sites and forests studied exhibited incre-
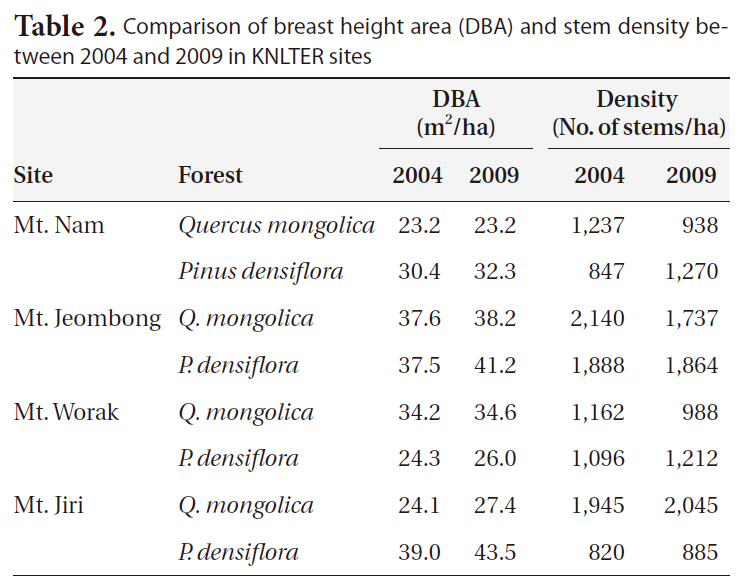
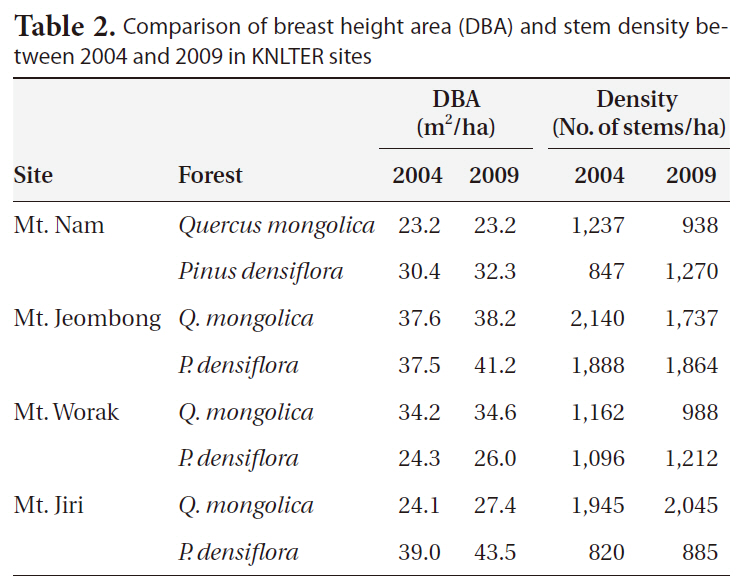
Comparison of breast height area (DBA) and stem density between 2004 and 2009 in KNLTER sites
ments in total BA, but Q.
Changes in stem density showed severe variation (Table 2). Abrupt changes in density occurred in the pine forest of Mt. N, evidencing the largest increase (33.3% as from 848 stems/ha to 1,270 stems/ha), and the oak forest exhibited the largest decrease (-31.8% as from 1,237 stems/ha to 938 stems/ha). Increases of density were in the pine (9.6%) forest of Mt. W and in the oak (4.9%) and pine (7.3%) forests of Mt. Ji. Stem density was reduced in the oak (-17.6%) stand of Mt. W and in the oak (-23.2%) and pine (-1.3%) forests of Mt. Je.
>
Changes in species composition
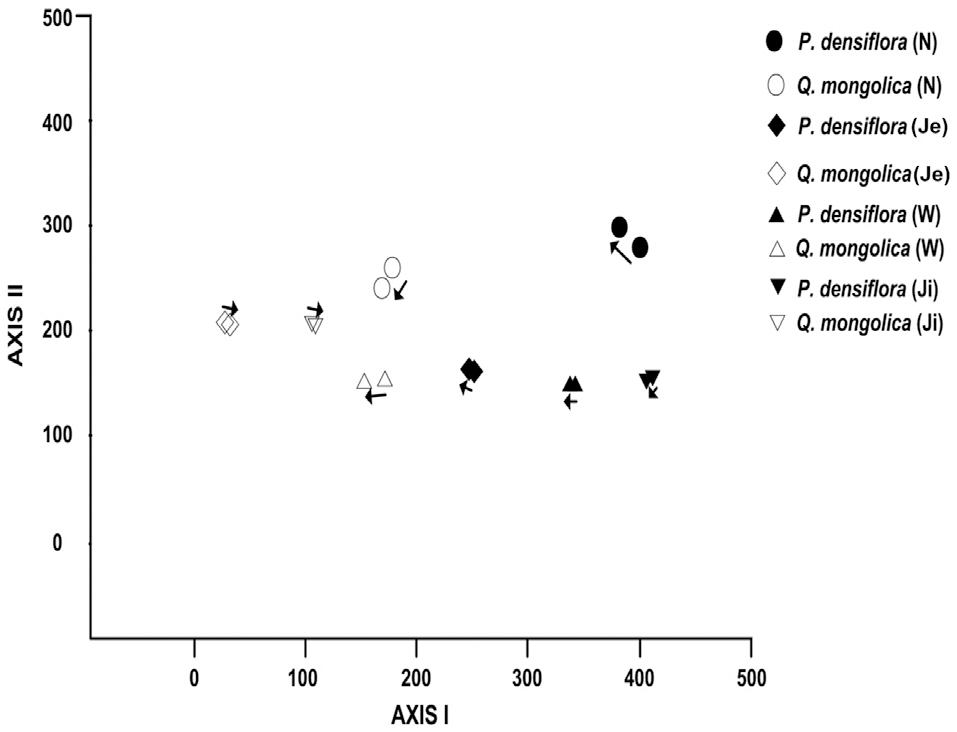
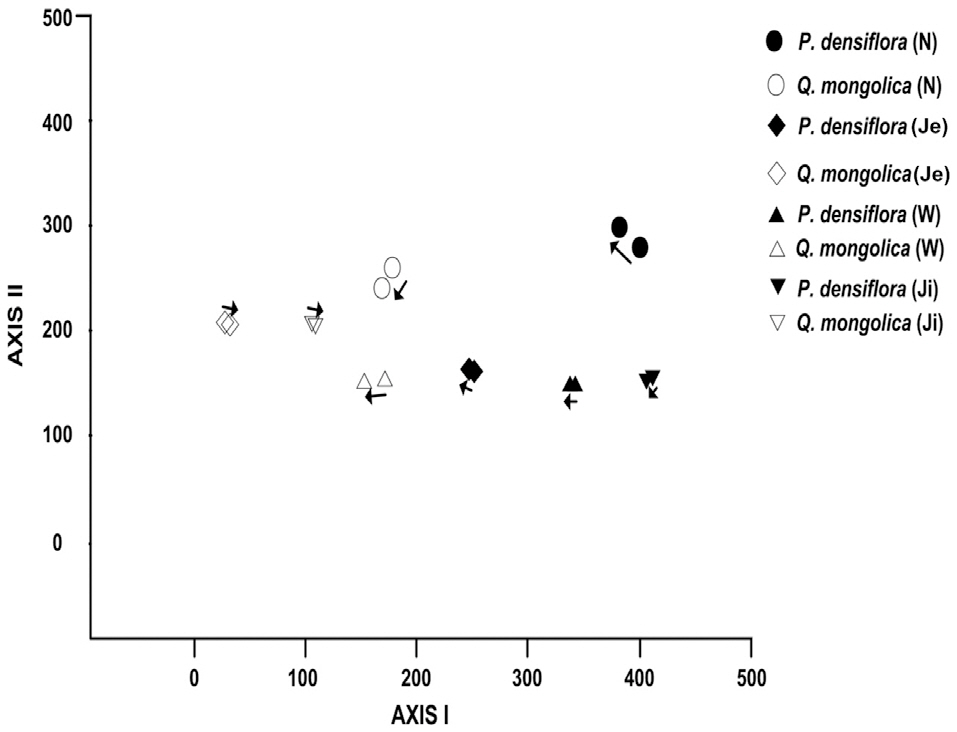
Compositional change in woody vegetation in the KNLTER sites was analyzed by applying DCA ordination method (Fig. 1). Altitude gradient was the representative factor for the differences in species composition, as sites in higher areas occupy the left side and those in lower areas were distributed on the right side, forest types (deciduous broadleaved and evergreen coniferous forests) also affected the results; the oak stands were clearly divided from the pines in the left and right sides, respectively.
Two patterns of compositional change--vegetation convergence and divergence--were noted (Fig. 1). Most of
the samples of oak and pine forests moved to the central part of the AXIS Ⅰ. The change in
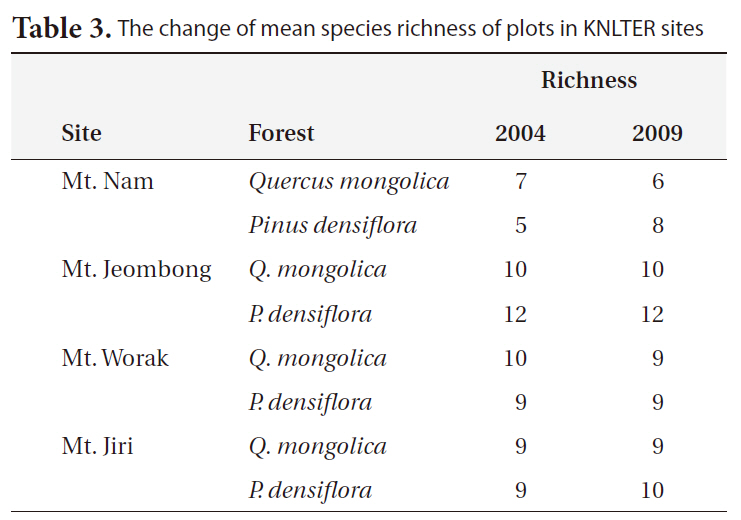
The changes in woody species richness for 5 years between 2004 and 2009 are shown in Table 3. In the P.
>
Mortality and recruitment rate
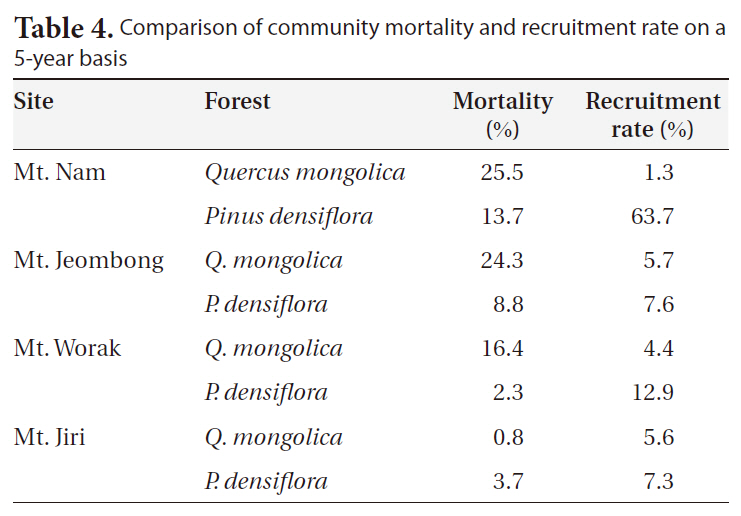
We calculated 5-year mortality and recruitment rates at the KNLTER sites (Table 4). With the exception of Mt.
[Table 3.] The change of mean species richness of plots in KNLTER sites
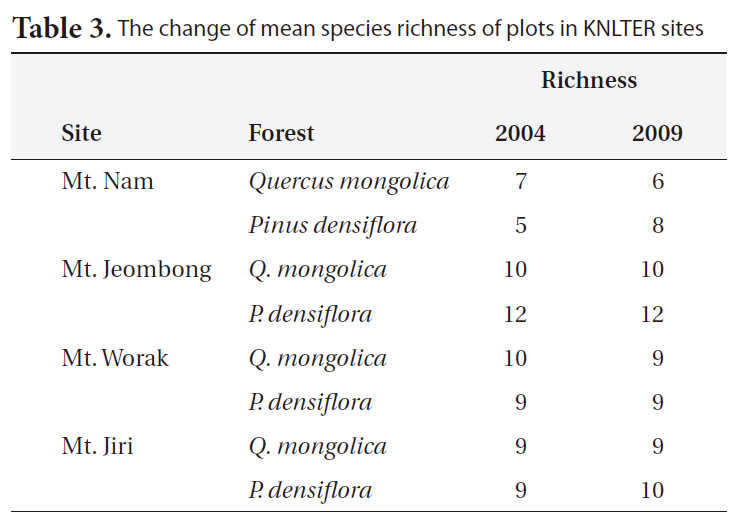
The change of mean species richness of plots in KNLTER sites
[Table 4.] Comparison of community mortality and recruitment rate on a 5-year basis
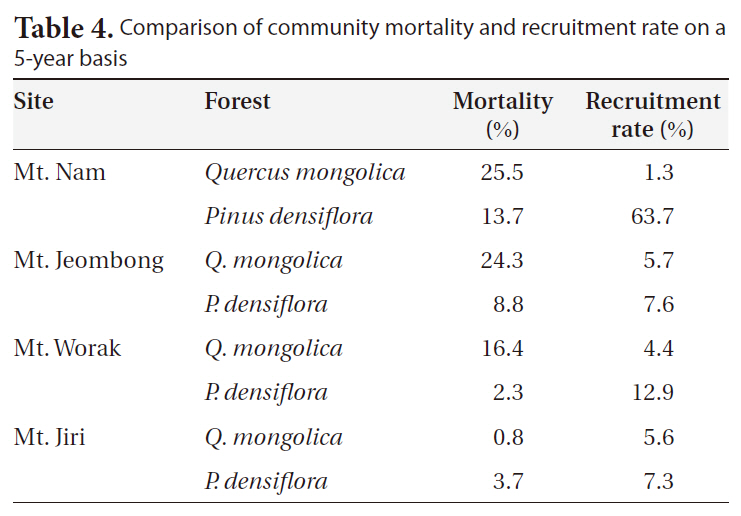
Comparison of community mortality and recruitment rate on a 5-year basis
Ji, the Q.
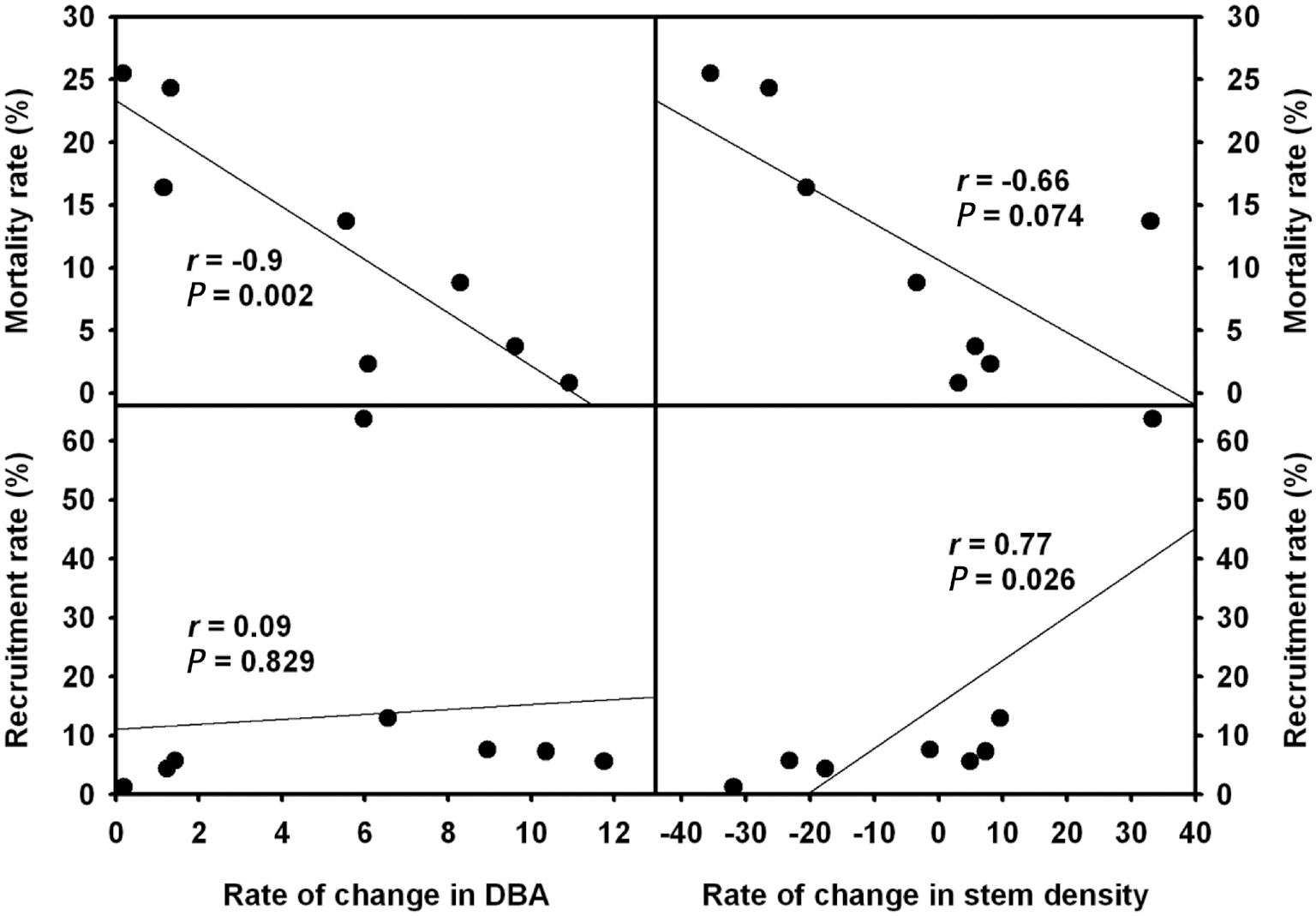
Overall correlation analyses between community demographs and rates of change in DBA and stem density were shown in Fig. 2. Mortality rate and rate of DBA change were strongly negatively correlated (
Climate change, which includes gradual pattern and extreme events, can affect the attributes (dynamics, composition, structure, productivity, diversity, etc.) of the biological community in various and complex ways. Ecosystem responses to climate change, which are the target of the KNLTER program, can be interpreted only by long-term observations with comprehensively and precisely designed systems. We presented only the results of the first re-measurement (5-year interval) on woody vegetation. Thus, our focus in discussion is on short-term findings and research feedbacks rather than the vegetation responses to climate change and regional weather.
>
Vegetation changes in KNLTER sites
Variations in structural changes were detected in this study, and such variations can be interpreted as the results of forest developmental processes and environmental differences among sites and vegetation types. Q.
forartificial disturbances (but see Lim et al. 2004). This also means a lack of background (or baseline) levels of variation in the structural attributes required to explain our results. Additionally, annual variations in BA and stem density depending on site condition may be involved in our results. In the Q.
Compositional convergence and divergence were illustrated in our ordination analysis. According to the results of DCA ordination, the direction of compositional changes in all the P.
Q.
Mortality and recruitment rate at the community level are not available in the inland regions of Korea. In our results, negative correlations were detected between mortality and rates of BA and stem density changes (Fig. 2). The absence of reference demographic data at the community level makes us difficult to understand and interpret. The one year-based mortality rate of a P. parviflora forest located on Ulleung Island was 2.1%/y (Lim et al. 2004) and our 5-year results were higher in the oak forests (except for Mt. Ji) and lower in the P.
>
Feedback for comprehensive and effective vegetation monitoring
In our research design, four dead classes (standing, broken, leaning and fallen dead) are established for individual trees. These classes are just mortality patterns and thereby cannot be used to determine causes of death, such as competition (endogenous process) or natural disturbances (exogenous process). Causes of death of tree species vary among species with different life histories or canopy architectures (Acker et al. 1996, 2003, Canham et al. 2001), and, can change in importance, especially during succession (Bible 2001, Canham et al. 2001).Thus, according to the established protocols of this study, parameters for cause of death such as suppression, pathogens, and insect-induced death and physical processes (strong wind, landslide and snow loading) should be added and quantified.
KNLTER is currently constructing background variations in the forest ecosystem. Such research activities will constitute a fundamental tool for the interpretation and modeling of ecosystem responses to climate change; in this regard, dividing of responses to succession and climate change is particularly relevant. A static monitoring period of at least 10 years is recommended for the accumulation of ground data, because even in old growth sites, approximately 5-10 years are required for the accumulation of base data (e.g., Forrester and Runkle 2000). Without data regarding background dynamics, the effects of gradual increases in weather events (e.g., effect of Typhoon Kompasu on metropolitan area), such as heavy rainfall and drought on ecosystems, cannot be explained.