A number of factors determine migratory birds’ distribution in habitats and their departure timing. Local land use patterns and global climate change have been implicated as relevant factors, occasionally resulting in losses of biological diversity (Vitousek 1994). Global warming is also responsible for earlier breeding onset in most animals, including birds (Brown et al. 1999). Cotton(2003) discovered evidence to suggest that global climate change has extended growing seasons, changed distribution patterns, and altered the phenology of flowering, breeding, and migration. The habitat characteristics and adjacent land use pattern play an important role in determining the composition of avian species (Lee et al.2010). Numerous scientific reports have recently become available worldwide; however, despite the fact that South Korean avifaunal habitats are recognized as important for migratory birds, only a small amount of systematic research has been achieved thus far.
Long-Term Ecological Research (LTER) is an ecological study approach involving the collection of both biological entities and their surrounding environmental characteristic information quantitatively, on the basis of regular intervals and long research periods. The LTER program can provide useful information for changes not only in biological species composition but also ecosystems’ changes, and allow researchers to detect the forcing variables that caused the changes by comparing the present with past information. Recently, the South Korean government initiated the LTER program to collect ecosystem information quantitatively since 2005 (Park et al. 2006), and the distribution of migratory birds and their ecological characteristics were discovered by the program. The program focused on the Upo Wetland System, which consists of four small compartments, henceforth referred to as the Upo Wetland, in terms of avifaunal distribution.
The Upo Wetland is listed as one of the Ramsar protection sites, and is an important wetland system especially for wintering migratory birds (Kang and Hahm 1997, Hahm and Son 1998, Kim 2000, Nasantogtokh 2002). This wetland also provides a breeding site for resident waterbirds (Lee et al. 2007), and is a wetland habitat group consisting of the Nakdong River, including the estuary, the Junam reservoir (Hahm and Son 1998), small wetlands in the river basin (e.g., Dalsung [Kim 2005], Gumi), and the Andong and Imha lakes. Specific characteristics exist in each of these wetland habitats, including different farming patterns, species distribution, size of wintering groups, protection levels, etc. (Park et al. 2006, 2008), and the Upo Wetland is geographically centered in the boundary of the habitat group.
In this wetland, we selected the Bean Goose (
In this study, we attempted to collect basic information regarding the population dynamics of the Bean Goose in the Upo Wetland. We first compared the current status of the population with that reported in the related literature in order to detect recent changes of population distribution over time. The secondary objective of this study was to determine the manner in which this species utilized the Upo Wetland as a habitat, and we finally compared their departure pattern with temperature changes inter-annually. The results of this study may provide us with basic information regarding migratory bird population changes in the Upo Wetland in relation to the land use patterns of human beings and meteorological variations. The future significance of those environmental characteristics to the Bean Goose population was also discussed.
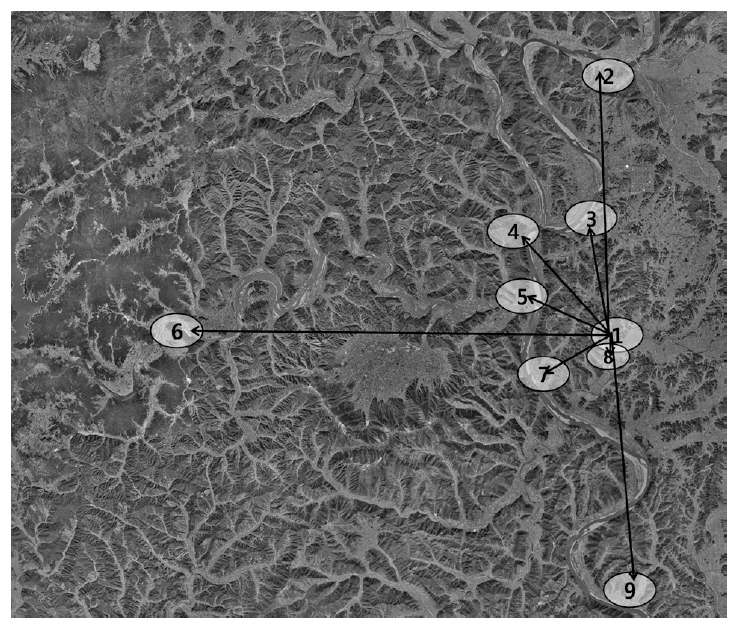
The Upo Wetland is geographically located at 35°33' N, 128°25' E, and 8.7 km2 in area size. This wetland was connected to the Nakdong River through the Topyeong Stream (Ministry of Land, Transport and Maritime Affairs 2008). The wetland system is composed of four small compartments: the Upo, Sajipo, Mokpo, and Jjokjibeol wetlands (Fig. 1).
>
Survey times and observation methods
We monitored the Bean Goose population at least once per month from November 2005 to November 2010. We conducted surveys on the weekdays in order to avoid human interference factors (e.g., ecotourism etc.). If the Bean Goose population movement was investigated due to local activities such as fishing, we surveyed the population again after 2-3 days since the occurrence of interference.
We established 13 observation points for bird counts using a Spotting scope (SWAROVSKI STM 80 HD; Swalovs-
ki Optik, Absam, Austria) and binoculars (12X, Nikon Vision Co., Ltd., Tokyo, Japan) and recorded the geographic location of every observation point using GPS (Garmin ColoradoTM 300; Garmin Corp., Taipei, Taiwan). We counted individual numbers of the Bean Goose, and recorded its behavior using a fieldscope (30X-50X; Swarovski) and binoculars (NIKON 10X 32HG L DGF; Nikon Vision Co., Ltd.). In order to measure the flight distance from the Upo region to the feeding areas, we followed the Bean Goose to the landing place. We recorded the geographical locations of the landing places with GPS and calculated the flight distances.
The Korean Meteorological Administration provided the monthly lowest, mean, and highest temperature values at the meteorological station (Changnyeong) during the study period, closest to the study site, and we analyzed the thermal effects on the birds’ migratory patterns. Because the majority of the Bean Geese departed the Upo Wetland in March, we also compared the population size of the Bean Goose monitored in March and the temperature status over this month. We compared two populations of Bean Goose between the Upo Wetland and the Junam reservoir located at 35°16' N, 128°19' E with 5.75 km2 in size at distances from approximately 33 km to the southeast from the Upo region (Fig. 1). The Junam reservoir’s Bean Goose population survey methods were the same as the Upo’s. The Junam survey was processed during the winters, from 2006-2008.
>
Dominant index and correlation
The Bean Goose was the most dominant species in the bird community of the Upo region since the 1997 survey to the present. We calculated the dominant index of the Bean Goose and other birds from the Upo Wetland via Simpson (1949)’s dominant index (λ) and Whittaker (1972)’s dominant index (D), in order to compare species distribution patterns among wintering birds at the Upo Wetland (Eq. 1).
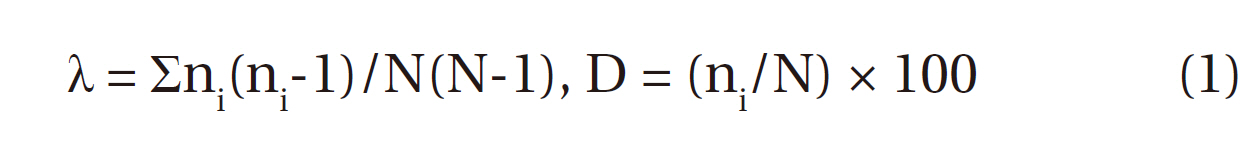
N = total numbers in community, ni = a species numbers in community
>
Changes of the species dominant index at the Upo Wetland
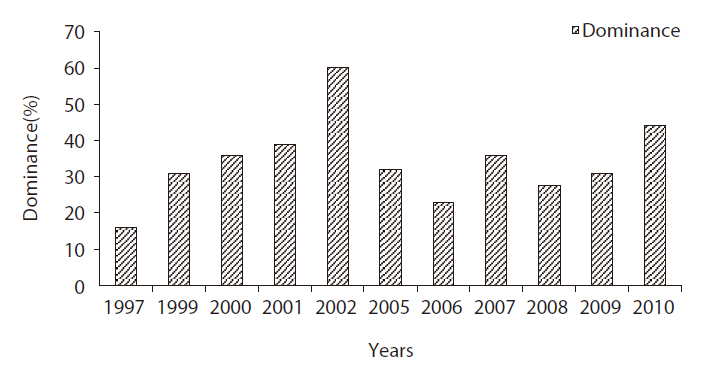
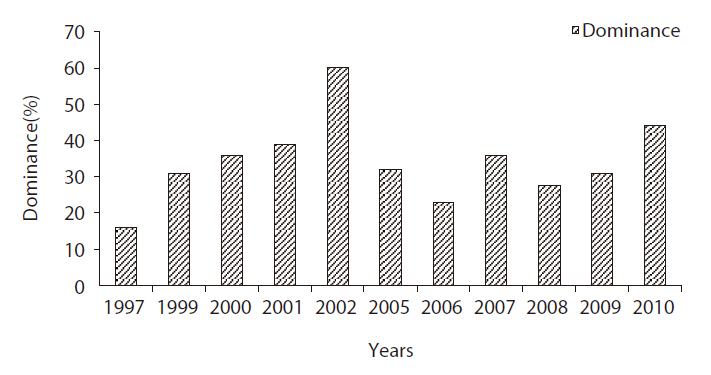
The species dominant indices of the Bean Goose at the Upo region increased continually from 15.9% (Kang and Hahm 1997) to 60.2% (Nasantogtokh 2002), and the monitoring results in this study showed a similar pattern of change in the dominance index. The dominance of the Bean Goose wintering at the Upo Wetland in 2010 was 44.0% (Fig. 2), which was as high as that observed at the Cheonsu Bay site located on the western coast (44.5%) (Yoo et al. 2008). From the dominance index, we determined that this species is the dominant species in the Upo Wetlands, and this may be attributable to the land use pattern in the regions surrounding the Upo Wetland area. The majority of rice paddy land was converted to onion or garlic cultivation fields during the winter season, which resulted in the reduction of food sources for bird populations. Other bird species, including ducks, dispersed to barley farm areas far removed from the Upo region, and the Bean Goose remained in the wetland.
>
Flight distances of the Bean Goose from the Upo to the feeding areas
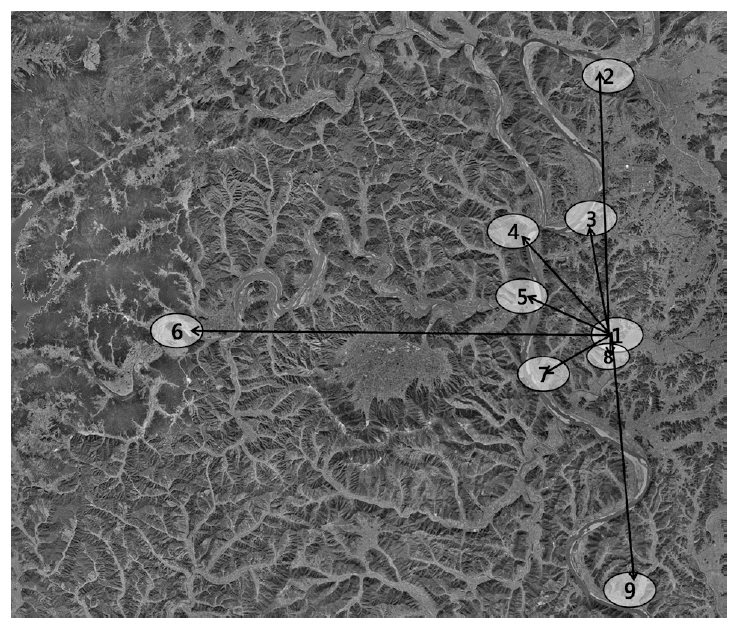
Even though the Bean Goose dominated the Upo Wetland and employed this system as their main habitat, the species moved everyday to find their food source outside the Upo Wetland boundary (Fig. 3). In large part, their feeding areas were barley and wheat fields at an average distance of ca. 6 km from the Upo. The Bean Goose visited Namji (13.5 km), Hyunpung (14.9 km), and the Hwang River (25 km) far from the Upo, in the Nakdong River basin. The species’ flight direction was diverse; however, the Bean Goose rarely used the eastern areas of the Upo Wetland. The eastern area was populated and a relatively small area of barley or wheat fields was maintained; this might be responsible for the selection of flight direction. When they foraged for food outside the Upo Wetland, they returned to the Upo to sleep.
[Table 1.] Correlation between populations of the Bean Goose and temperatures at the Upo Wetland

Correlation between populations of the Bean Goose and temperatures at the Upo Wetland
>
Changes in the wintering population of the Bean Goose
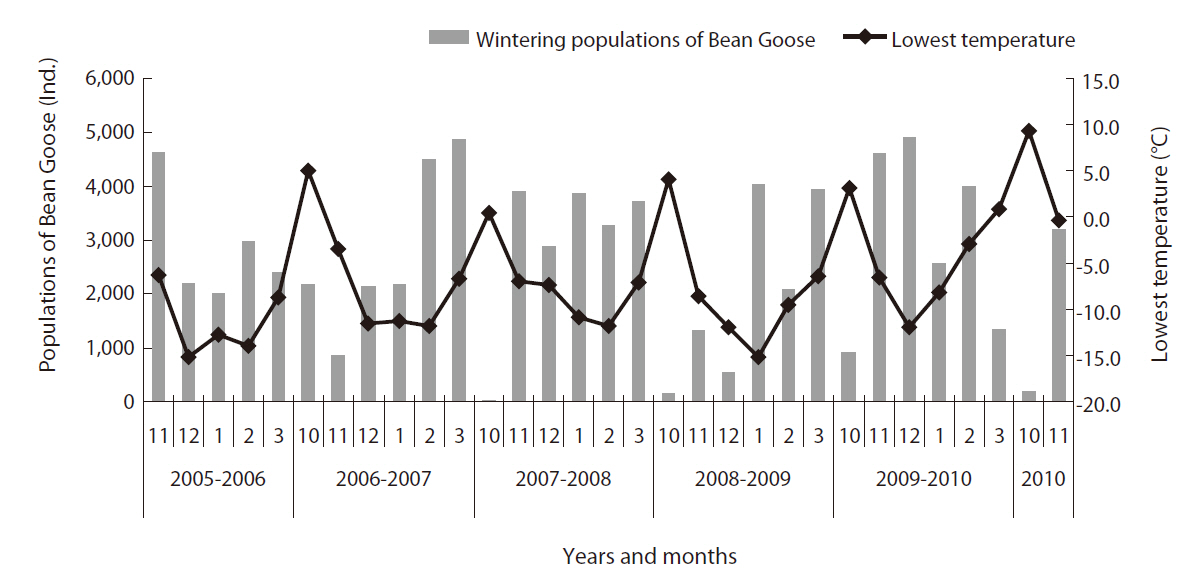
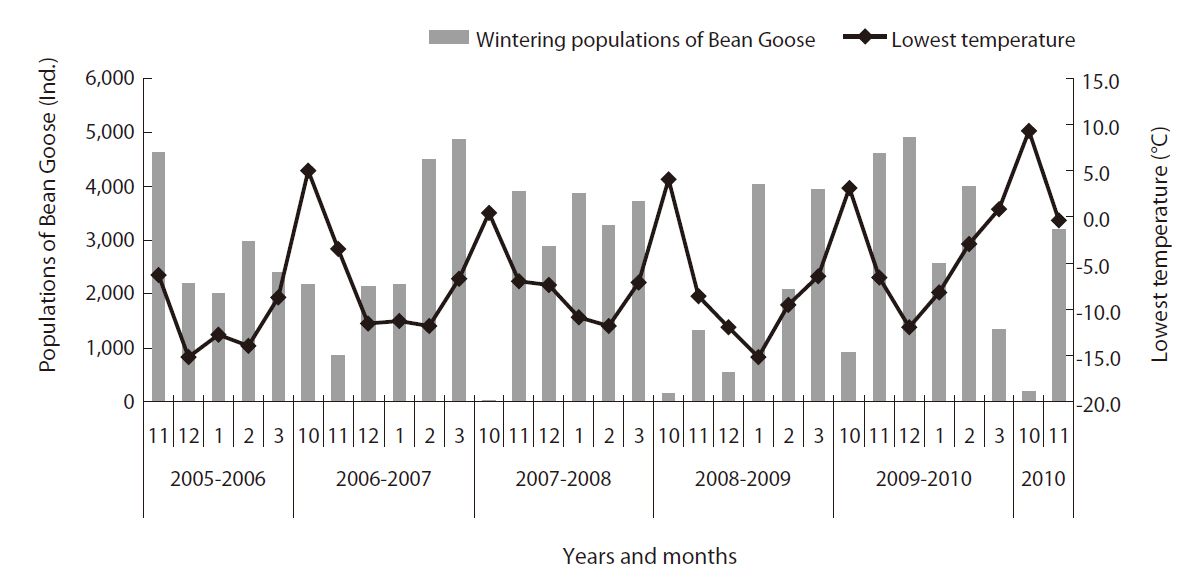
During the study period, we analyzed the relationship between the wintering population of the Bean Goose and temperature in winter (October to next March) at the Upo Wetland site (Fig. 4). When the lowest temperatures at the Upo in October were between 0°C and 5°C, the Bean Goose population size was ca 1,000 individuals. When the lowest temperature in the middle of winter (December to January) was between -5 to -10°C, the population size of the Bean Goose decreased to ca 550. Correlation coefficient between monthly observed individual numbers and lowest temperature was -0.461 (P = 0.009) (Table 1). And mean, highest temperature did not show statistical significance. That is, the greater the reduction in temperature, the greater was the increase in Bean Goose individuals wintering in the Upo Wetland. van Noordwijk (2003) proposed three hypotheses that might have explained the earlier passage of long-distance migrants. First, they may leave Africa on the same date but are able to complete their migration more rapidly because foraging conditions in early spring in Europe are better in years when the North Atlantic Oscillation Index (the NAOI, an index of air pressure over the North Atlantic Ocean) is high, giving rise to warmer, wetter, and windier conditions. Second, if the weather in Africa is correlated with the NAOI, then seasons in Africa may also be, which in turn indicates that the birds may leave earlier. Third, the weather in Africa may not have changed, but natural selection may have altered the trigger event initiating migration (Cotton 2003).
>
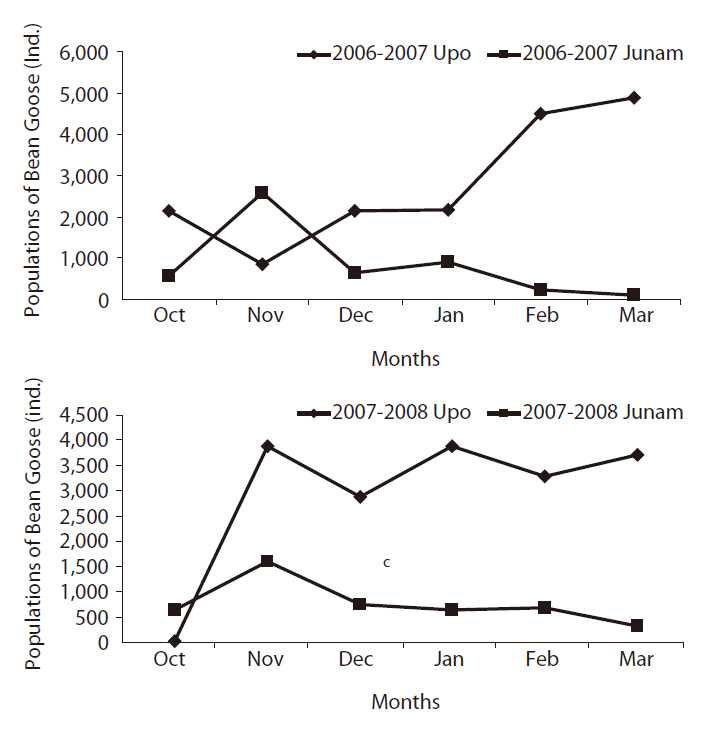
Comparison of the Bean Goose populations between two neighboring wetlands
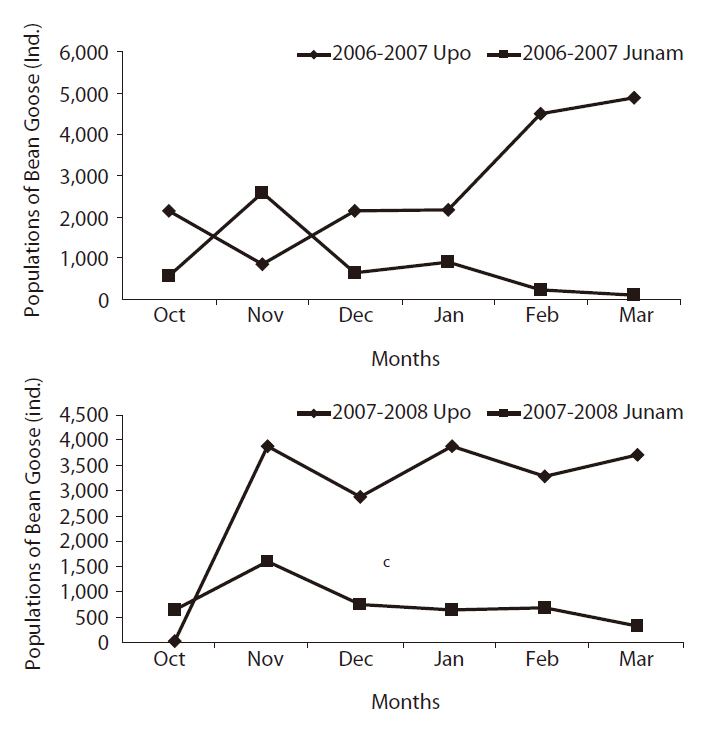
The co-migration pattern of the Bean Goose between the two habitats has been evaluated in a previous study (Lee 2007). The results were as follows; 1) The population of the Bean Goose in the Upo region have wintered more than that of the Junam, 2) especially, with fluctuations in winter temperature, the Bean Goose population between the Upo and the Junam rose and fell, it appeared that the Bean Goose evidenced a departing and returning migration behavior between two wetlands (Park et al. 2006, 2008).
>
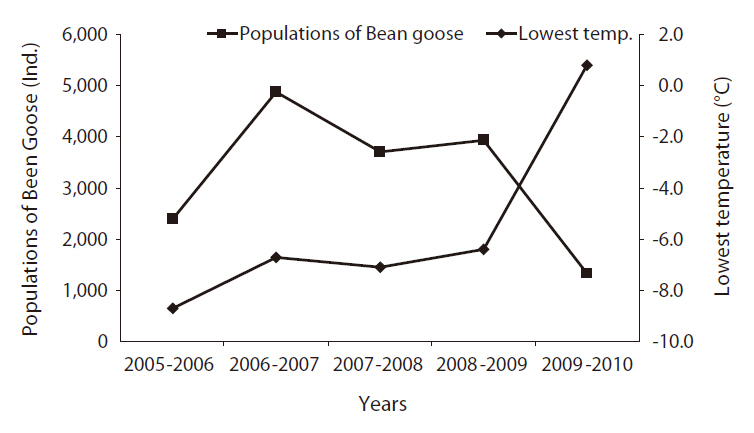
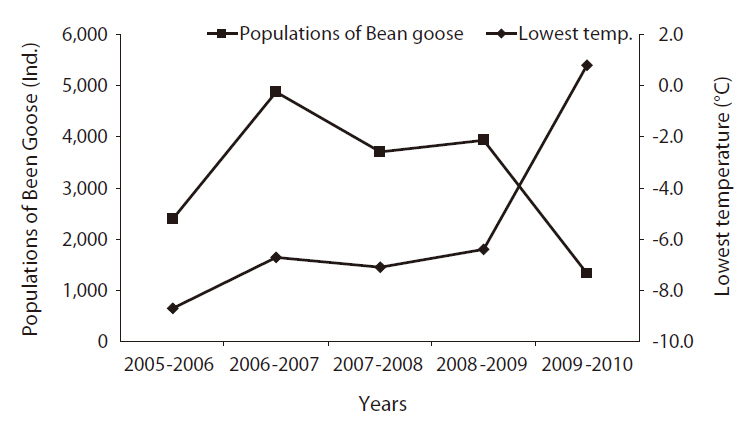
Lowest temperature and population change of Bean Goose at the Upo Wetland in March
Wintering birds return to their original habitat in March. During the study period, the lowest tempera-
tures in Mar. in the Upo area gradually increased, and the relatively high temperature in Mar. resulted in an increase in the Bean Goose population at the Upo Wetland area (Fig. 5). The Bean Goose population from southern areas moved to the Upo and made groups prior to their migration to the north (Lee 2007). However, a sudden high warm temperature in March 2010 (Fig. 6), after the severely cold weather in December 2009, caused a Bean Goose population to migrate very early from the Upo to the north. The drop in the Bean Goose population in Mar. 2010 may be attributable to the departure of birds to the northern region prior to the regular monitoring date.