We monitored the zooplankton community dynamics of a regulated river system (the Nakdong River) in South Korea,in an effort to characterize the relationship between basin rainfall quantity and changes in zooplankton community structure. The river studied herein has four multipurpose dams upstream and one estuarine barrage at the river mouth,resulting in intensive flow regulation. We hypothesized that flow regulation would effect zooplankton community structure changes not only within the summer period of concentrated rainfall, but also during the subsequent seasons. Field monitoring was conducted on a weekly basis (1999 to 2009) at the study site (27 km upstream from the estuary dam).The studied years were divided into two groups: rainy years with annual rainfall greater than total average annual rainfall,and dry years (years with lower than average annual rainfall). The zooplankton community data was also divided into two groups according to year and community structural characteristics. The summer density of zooplankton was low during the summer concentrated rainfall period. In the autumn, zooplankton density was statistically related to river flow (r2 = 0.30, P < 0.05, N = 11), which was affected by the summer concentrated rainfall (r2 = 0.31, P < 0.05, N = 11).Furthermore, autumn zooplankton density was positively related to that observed in the summer (r2 = 0.53, P < 0.05, N= 11). Therefore, it can be concluded that summer concentrated rainfall can affect the following seasons’ zooplankton densities caused by dam flow control, and the potential growth rate of zooplankton
Flow is a primary factor that determines the dynamics of river ecosystems (Descy 1987, O’Farrell 1993, Ha et al.1998, Ha et al. 2002, Shcherbak and Bondarenko 2005),and this is the case in both mountainous areas (Iroume et al. 2010) and lowland rivers (Jeong et al. 2007). Rivers or streams have different habitat structures resulting in different community structures between up- and down-stream reaches; this is known as the river continuum concept of Vannote et al. (1980). Currently, it is somewhat difficult to find natural lotic systems, because of the increased importance of water resource management in relation to climate change; therefore, flow regulation is a common characteristic of river systems, at least in Far Eastern Asia, which includes Korea, Japan, and China (Tharme 2003).
The influence of flow regulation on community structure in lotic systems has been reported on extensively in the past few decades (Jeong et al. 2007). The accumulation of nutrients in the reach of flow regulation has resulted in the acceleration of eutrophication (Joo et al. 1997), which also generated increases in phytoplankton density and dominance by harmful algal species such as cyanobacteria (Ha et al. 2002, Teichberg et al. 2010). Reservoir-like patterns in the microbial food chain have also been frequently observed in regulated river systems (Kim and Joo 2000).
In eastern Asia, rainfall typically evidences a clear seasonality induced by monsoonal climate or typhoon events. This meteorological characteristic is responsible for the water flow changes in lotic systems, which have a ‘resetting’ function on ecological community structures (Park et al. 2002). Summer concentrated rainfall frequently introduces dynamic changes not only of physico-chemical factors (Joo and Francko 1995) but also of bacterial and plankton communities (Ha et al. 2002). Functional changes in ecological communities have also been reported in relation to summer rainfall (Allan and Castillo 2007). The heterogeneity of seasonal rainfall distribution in this region introduces some difficulty in managing water resources, such that lotic systems in Far Eastern Asia have experienced intensive alterations of stream structures, principally as the result of the installation of locks and dams. The aforementioned ecological characterisics of regulated rivers can be readily observed in this region.
Zooplankton constitutes a route for energy and material flow between producers and other consumers in river systems. They graze on phytoplankton, resulting in a reduction of algal biomass (Lampert et al. 1986, Kim 2001); moreover, they become food sources for young fish (Chang et al. 2004). Sometimes water quality improvements can be observed in regulated river systems, due to extensive grazing activity on phytoplankton biomass by zooplankton (Power 2001). Intensive flow regulation increases water residence time, which is a primary factor in determining zooplankton community structure and functions in river systems (Havel et al. 2009), and strong grazing activity has been previously reported in regulated river systems (Kim and Joo 2000).
The Nakdong River is a good example of a regulated river system in the Eastern Asian region, and is also part of a Long-Term Ecological Research (LTER) program that is currently underway. In the past decade, research has identified the role of zooplankton in this river system, 50and the greatest growth of zooplankton in this river is usually observed in the spring (March to May) or autumn (late September to November) (Kim and Joo 2000, Kim et al. 2000, 2001, 2002, 2003). Those studies have hypothesized that the zooplankton community in a regulated river system would be affected profoundly by water flow regulation; however, this hypothesis referred to overall patterning, and was focused primarily on spring zooplankton growth, during which time the river was typically stable due to limited rainfall and the resultant low flow. Comparatively, the autumn zooplankton community has been insufficiently understood in regard to changes in river flow caused by concentrated summer rainfall. Therefore, in this study, the zooplankton community structure data obtained from the Nakdong River LTER database was employed to evaluate the influences of summer rainfall on autumn zooplankton dynamics.
The Nakdong River, which has an approximate drainage area of 23,817 km2, is one of the largest river systems in South Korea, and travels ca 521.5 km. Water flow in the Nakdong River is regulated by seven multi-purpose dams (Andong, Imha, Hapcheon, Namgang, Yeongcheon, Miryang, and Unmoon) and an estuarine barrage, which results in eutrophication in the lower reaches (Ha et al. 1999, Kim et al. 2001). The study site is located 27 km upstream from the estuary dam (Fig. 1). Weekly monitoring of the zooplankton community was conducted for approximately 11 years, from 1999 to 2009.
>
Rainfall, discharge, and water quality measurement
Two hydrological parameters (rainfall and discharge) were evaluated, in this study, in an effort to characterize hydrological influences on zooplankton community changes at the study site. Daily rainfall data from meteorological stations in the Nakdong River basin (Andong, Daegu, Hapchon, Jinju, and Miryang) were used in this study, and were provided by the Korea Meteorological Administration. Daily river flow data was provided by the Water Management Information System (WAMIS; http://www.wamis.go.kr), and discharge data was from the Samryangjin station, the discharge station closest to the study site. Three months’ daily rainfall data was summed (i.e., June to August), and the daily discharge data was averaged for comparisons of seasonal or inter-annual variability.
Water samples were obtained at a depth of 0.5 m, placed into 20 L sterile polyethylene bottles, and maintained in the shade at ambient temperatures until returned to the laboratory. Water temperature, % saturation of dissolved oxygen, and conductivity were measured with a YSI Model 58 meter (YSI Incorporated, Yellow Springs, OH, USA) at the study site immediately after collection. Phytoplankton biomass (chlorophyll a), total nitrogen (TN) and total phosphorus (TP) were determined spectrophotometrically, based on the method of Wetzel and Likens (2000).
>
Zooplankton collection and identification
For the determination of zooplankton density, an 8 L water sample was collected at a depth of 0.5 m. The collected water was filtered through a 35 ㎛ mesh net, and preserved with 5% formaldehyde. Zooplankton were orgcounted at individual levels per 1 L using an inverted microscope and identified to the genus or species level (Koste 1978, Smirnov and Timms 1983).
We applied four statistical methods to the LTER data (i.e., hydrological, water quality, and zooplankton data sets): factor analysis (FA), one-way analysis of variance (ANOVA), cross correlation (CC), and regression. FA was used to reduce the dimensionality of the monitored environmental parameters and zooplankton community datasets, and the relationship among those input parameters was examined. Factor extraction was conducted based on the principle component analysis (PCA) method, and Varimax rotation was applied to sort extracted factors more meaningfully. Factors with eigenvalues over 1 were primarily selected among the extracted factors, and a scree plot was used to select the most ‘meaningful’ factors (Costello and Osborne 2005). We used all environmental and zooplankton data collected to identify general patterns in the environment and zooplankton community. ANOVA was used for comparisons of the zooplankton community based on years with different rainfall distribution, to clarify summer rainfall impact. We used average total summer rainfall to divide the 11 years of the study into two groups: ‘rainy years’ with annual rainfall higher than total average annual rainfall, and ‘dry years’ with annual rainfall less than the total average annual rainfall. Zooplankton community data was also divided into two groups in accordance with this rainfall/year criterion, and the community structural characteristics were assessed. Inter-annual variations in the zooplankton community were statistically analyzed via ANOVA.
Because the principal focus of this study was to evaluate the delayed influence of summer concentrated rainfall on the following season’s zooplankton community, we also used a time-series analysis method in CC. Monthly summed rainfall quantity and averaged zooplankton community data were compared, and correlation factors between rainfall and zooplankton were obtained. Finally, we applied regression analysis to determine the quantitative influence of summer rainfall on changes in the autumn zooplankton community. All statistical significance assessed was evaluated at α = 0.05, under an environment of statistical shell SPSS for Window ver. 14.0 (SPSS Inc., Chicago, IL, USA).
>
Hydrological and limnological characteristics
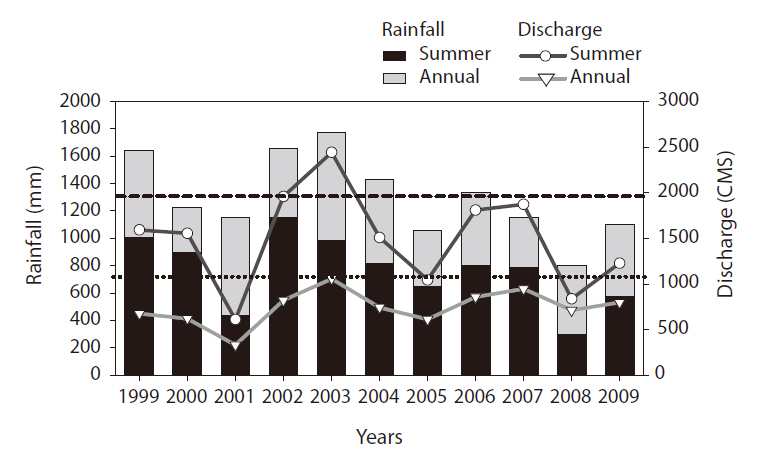
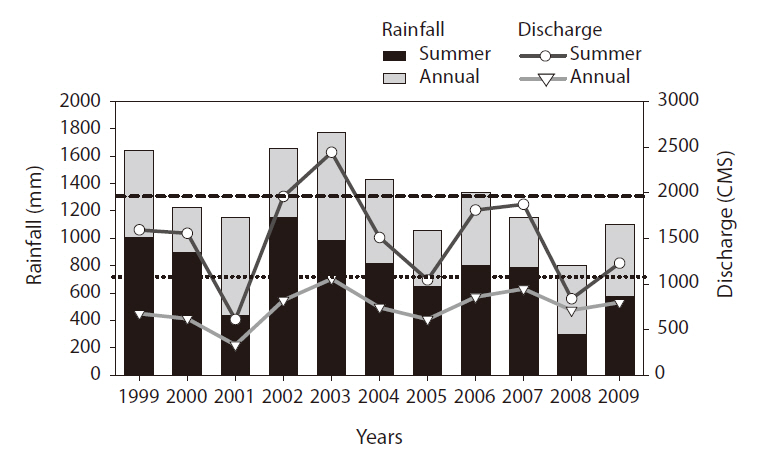
Strong inter-annual variability was noted in rainfall during the study period (Fig. 2). Rainfall in the summer (June to August) and total annual rainfall evidenced similar changing patterns (
Summer discharge had a pattern similar to changes in summer rainfall (

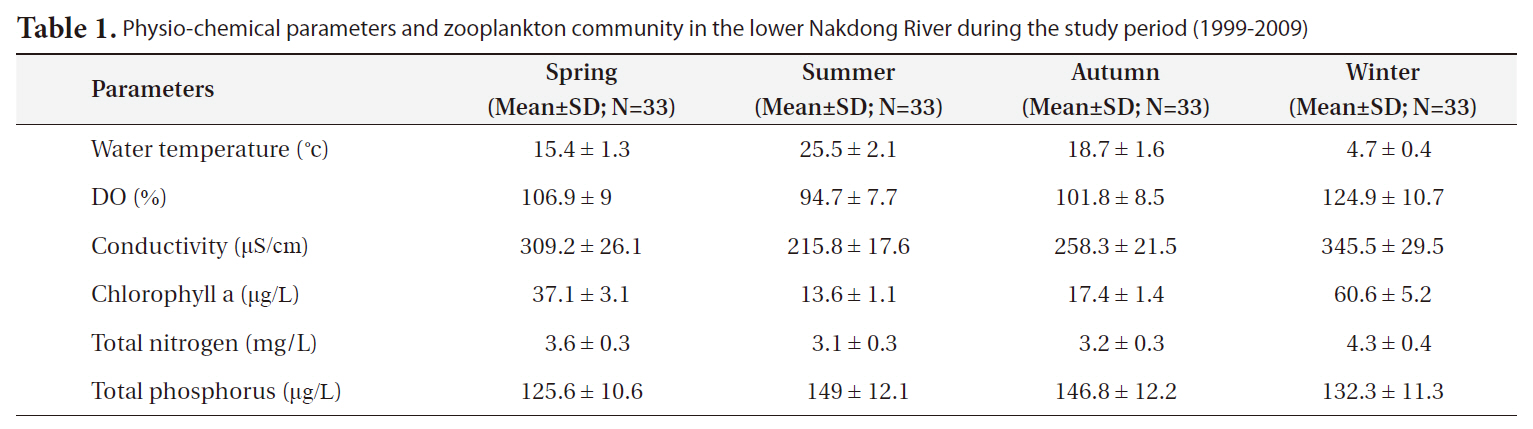
Physio-chemical parameters and zooplankton community in the lower Nakdong River during the study period (1999-2009)
2). The lowest summer discharge was recorded in 2001 (611 cubic meters per second [CMS]), whereas the highest discharge was noted in 2002 (1,959 CMS). Annual and summer discharge data evidenced a significant positive relationship (
For the last 11 years, the lower Nakdong River evidenced seasonality in limnological characteristics (Table 1). Water temperature exhibited the strongest seasonality, and was highest in the summer and lowest in the winter. The percent of dissolved oxygen (DO%) varied depending on temperature, and the chlorophyll concentration tended to increase from winter to spring. TN and TP changed slightly with the season; however, these differences were not large.
>
Zooplankton community changes
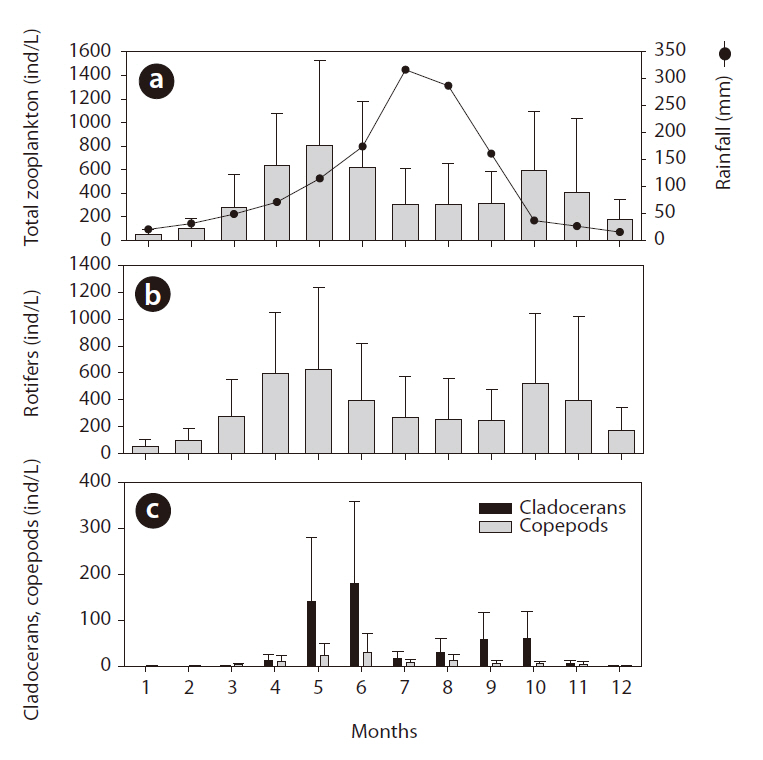
During the study period, 147 species were observed, including 114 rotifer species, 22 cladoceran species, and 12 copepod species. Rotifers dominated the zooplankton community, with Polyarthra sp. and Keratella cochlearis being the dominant species. As compared with planktonic rotifers, benthic species (such as Lecane sp. and
>
Relationship between environmental parameters and zooplankton
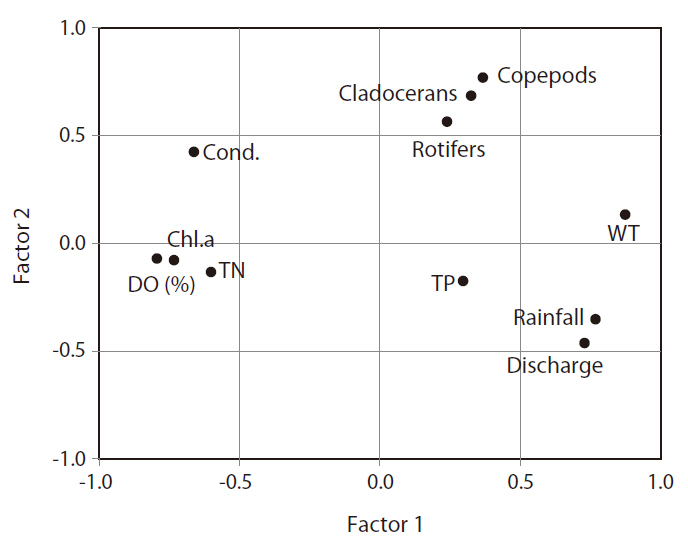
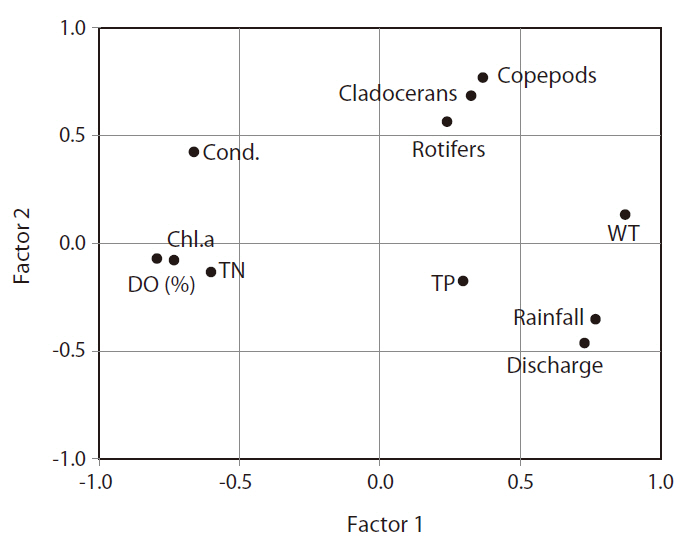
The application of FA extracted 11 factors from the input database (Fig. 4). Among the 11 factors, three evidenced eigenvalues exceeding 1, and the cumulative explained variance of these three exceeded 65.9%. Fig. 4 shows the first two factors (variance explained, 54.8%) selected based on a scree plot, as well as eigenvalues > 1. Factor 1 consisted of rainfall, discharge, water tempera-
ture, DO (%), chlorophyll a, and TN, and we designated this factor as ‘phytoplankton growth factor.’ Increased temperature, rainfall, and discharge induced a decrease in the concentration of chlorophyll a. The second factor mostly consists of zooplankton community data and hydrology (i.e., rainfall and discharge), which can be interpreted as ‘rainfall impact on zooplankton.’ Conductivity was relatively close to factor 1, and TP concentration was not related strongly with either factor 1 or 2.
>
Importance of summer rainfall on autumn zooplankton
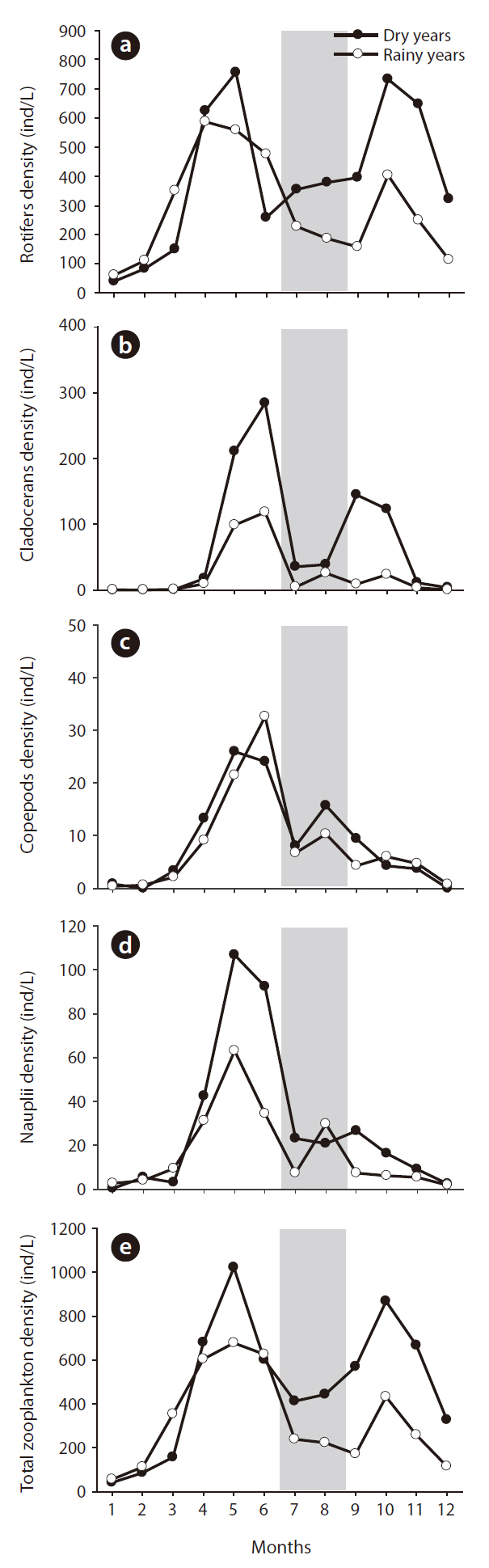
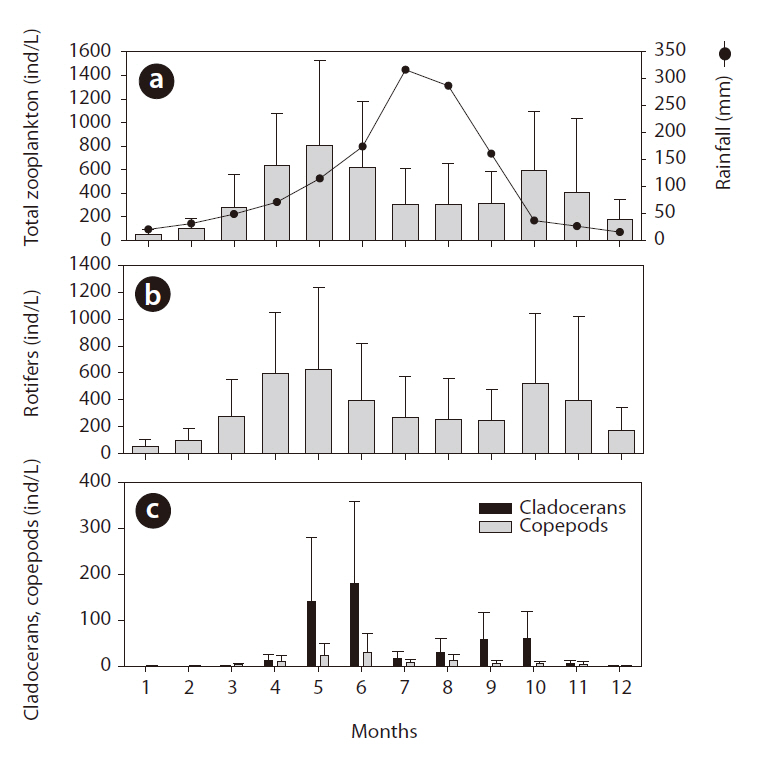
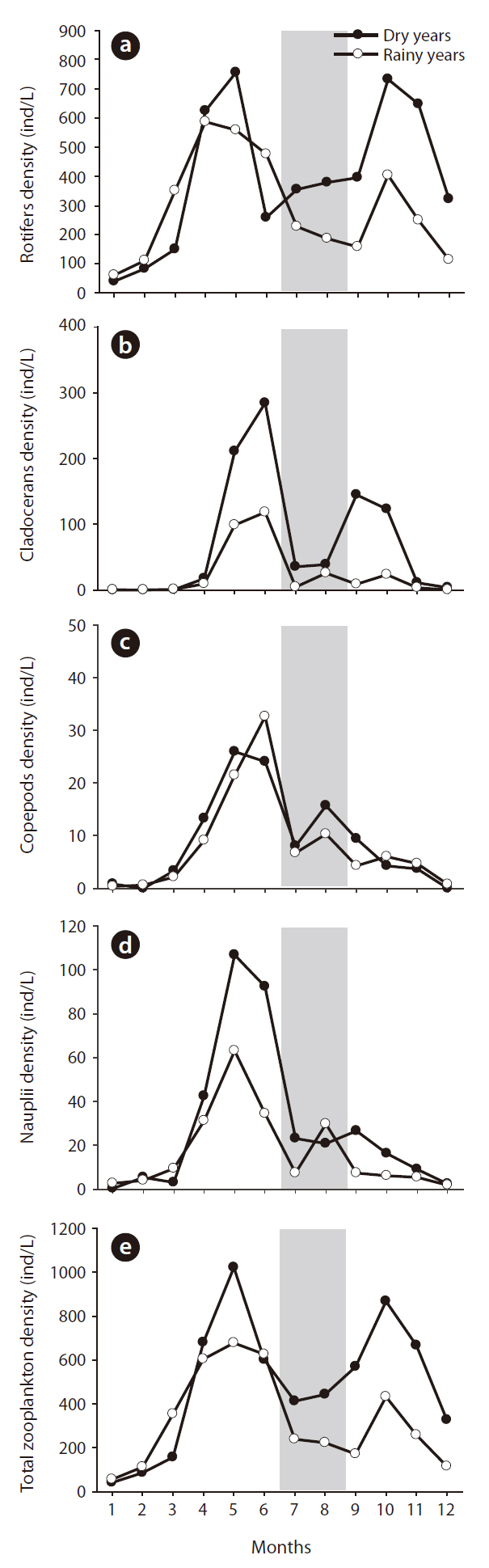
Summer rainfall quantity was statistically related to zooplankton community structure changes (Fig. 5). Zooplankton evidenced relatively lower density when summer rainfall was concentrated in the summer, and in the following autumn the zooplankton density differed according to the increase and decrease in summer rainfall. In dry years, autumn zooplankton density increased to the spring level (spring, 619 ind/L; autumn, 701 ind/L); however, in rainy years the autumn zooplankton density decreased to half of the spring level (spring, 545 ind/L; autumn, 287 ind/L) (one-way ANOVA, F = 16.714, P < 0.01). Rotifer density in both year groups was similar to the pattern detected with total zooplankton and evidenced distinct differences according to the quantity of summer rainfall (
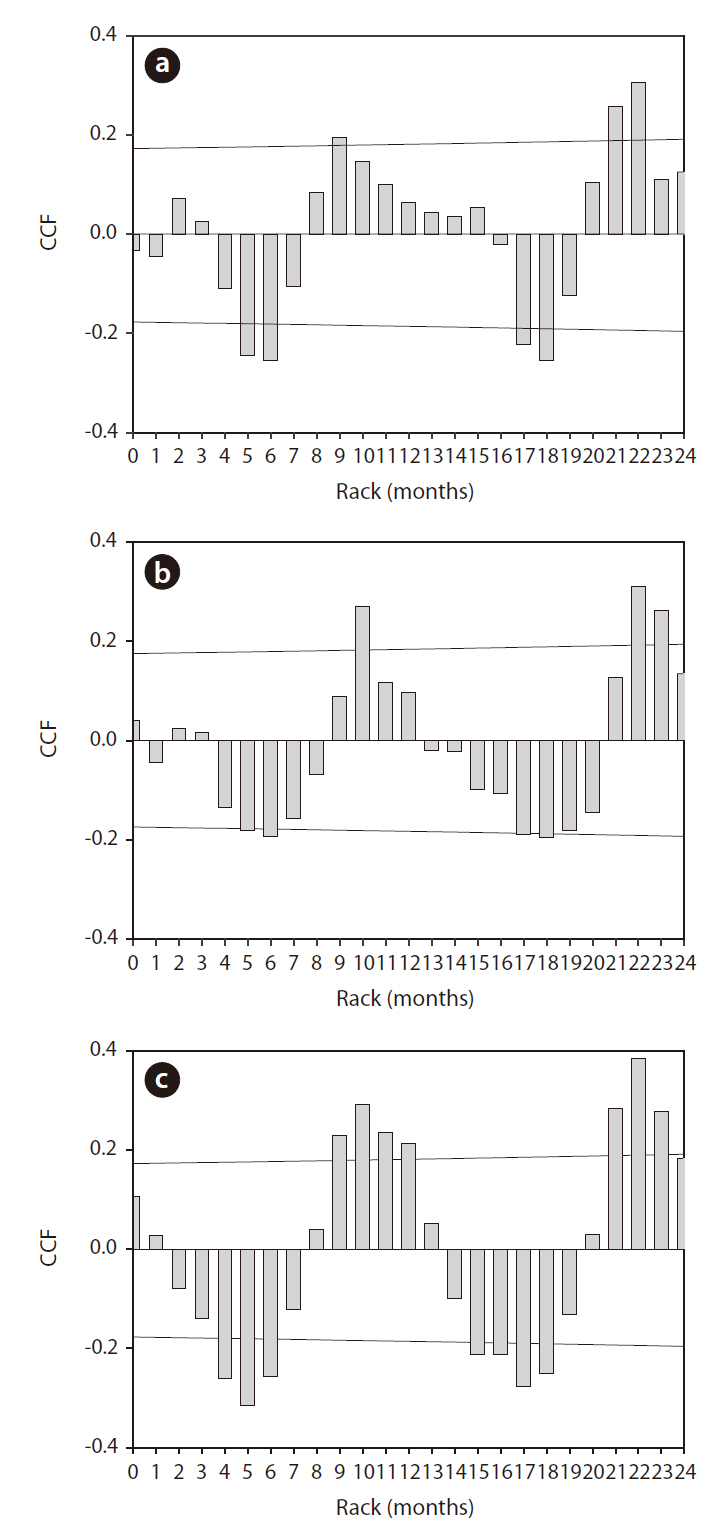
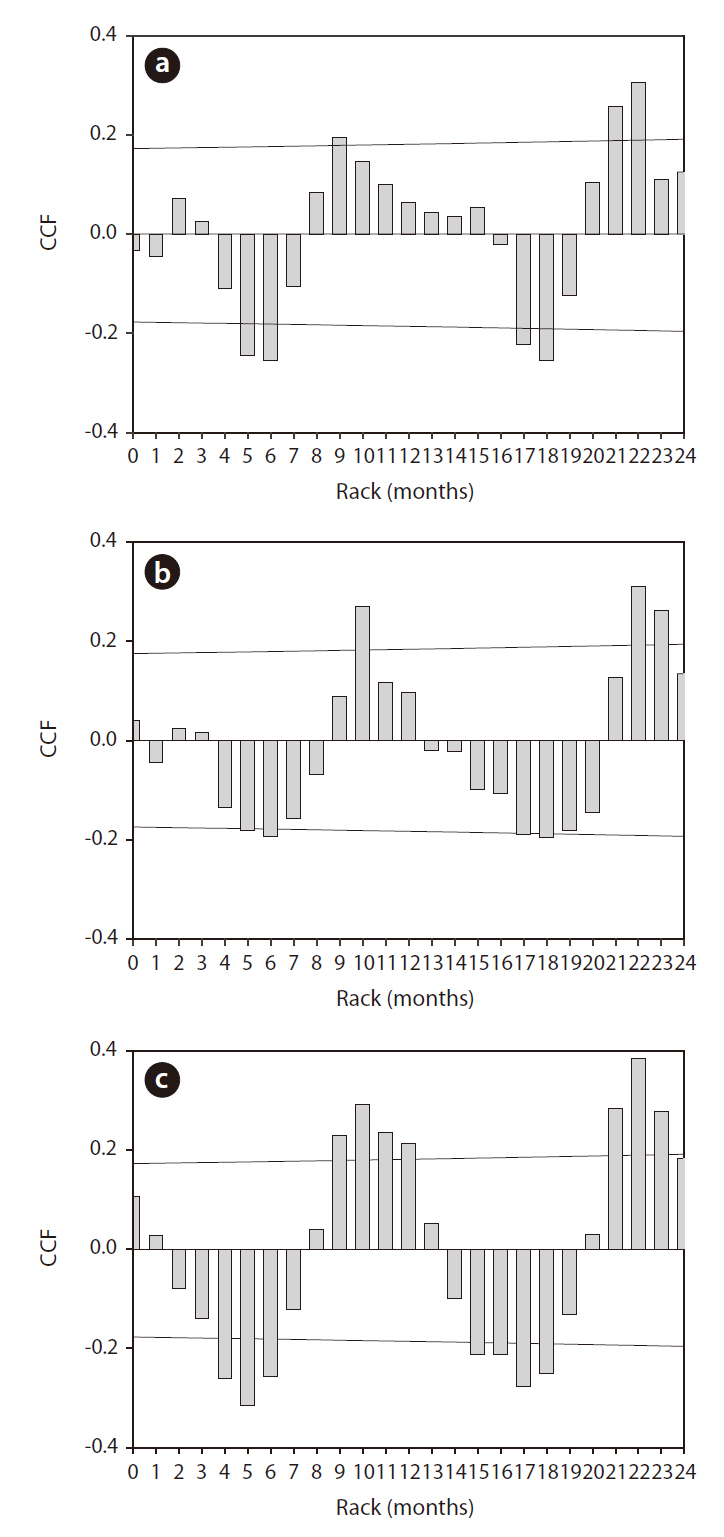
The application of CC to rainfall over the zooplankton community data revealed a significant time-series relationship (Fig. 6). Rainfall generally had a significant negative impact on the changes in zooplankton community density, and the period of delay was approximately 4-6 months. Among the three groups of zooplankton, rotifers and copepods responded strongly to rainfall, which occurred several months earlier. The cladocerans evidenced patterns similar to those two groups, but the
degree (i.e., cross correlation factor) was not as strong.
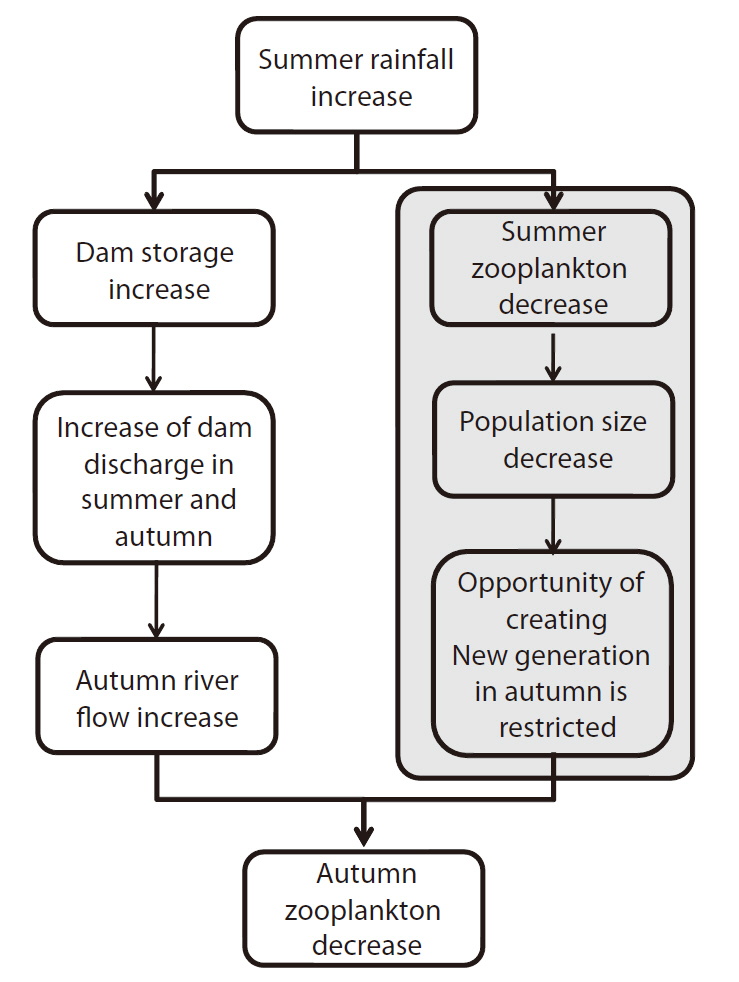
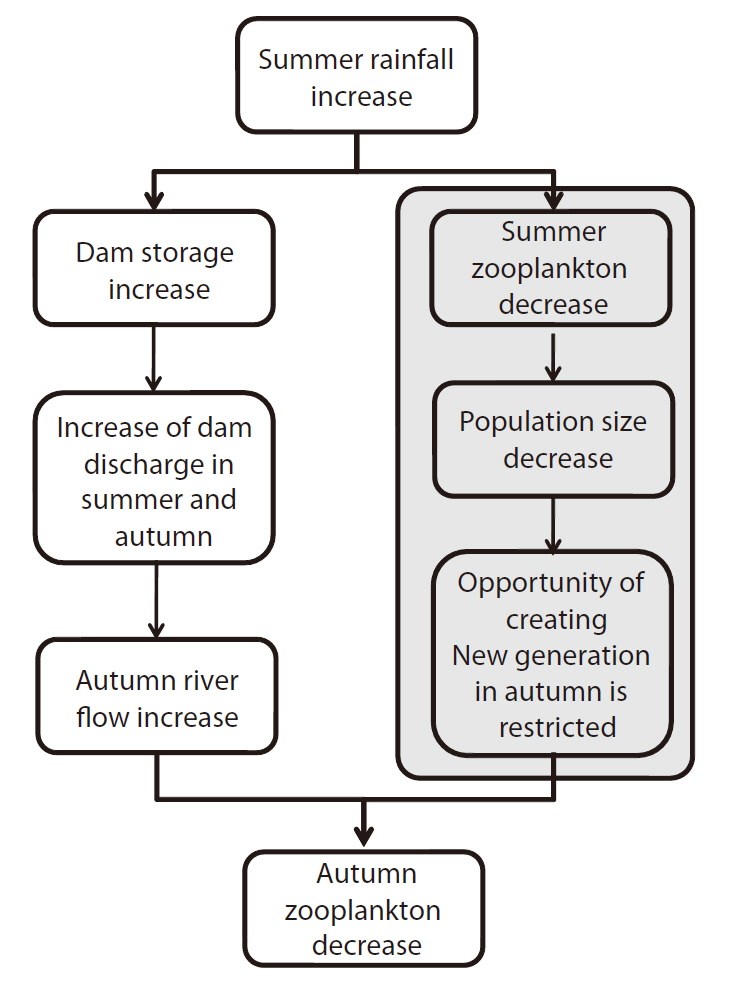
Regression analysis showed the existence of a possible relationship between rainfall, discharge, and zooplankton (Fig. 7). With this result, we can draw a plausible sequential direction that explains the impact of summer rainfall on the autumn zooplankton community. First, rainfall in summer increased the summer and autumn river flows significantly (Fig. 7a and 7b). The increased autumn river flow then negatively affected the autumn
zooplankton community (Fig. 7c). In addition to the hydrological impact, the zooplankton density in summer significantly affected the following season’s zooplankton density in a positive manner (Fig. 7d). Even though summer zooplankton density was not related statistically to summer rainfall quantity (
South Korea is subject to a monsoonal climate and experiences several typhoons during the summer season (typically from June to early September); this meteorological characteristic is believed to be responsible for the concentrated summer rainfall (Kim et al. 2005). Rainy summers and dry winters are a typical pattern in East Asia, which differs from Europe and Australia, where spring discharge originating from snow melt dominates river flow in Europe (Allan and Castillo 2007), and Australia has a relatively significant rainfall in the winter (Murphy and Timbal 2008). The rainfall pattern in the southwestern United States is slightly similar to that of the Eastern Asian region, owing to the presence of a partial monsoon climate (Hu and Feng 2002); however the dry season there is from May to June, unlike in the Asian region. Therefore, summer concentrated rainfall in South Korean peninsula is thought to be a primary water source, since the other seasons are characterized by a relative lack of rainfall.
In this study, autumn zooplankton density was apparently affected by concentrated summer rainfall; comparatively, the ‘dry years’ in this study showed almost similar magnitudes for zooplankton density relative to the spring seasons. Two possible factors can be considered to be responsible for this phenomenon, (1) stable water flow in autumn, which was caused by a lack of summer rainfall (Kim et al. 2003), and (2) a preferable water temperature for zooplankton growth (Richardson 2008). Even though water turbulence information was not directly utilized (e.g., water velocity or retention time), the autumn seasons in ‘dry years’ were believed to sustain relatively stable water flow with temperatures similar to the summer temperatures.
When community structure was compared, the roti-
fer group in autumn was impacted profoundly by concentrated summer rainfall. Zooplankton species, in this group, are typically small and frequently fail to struggle against increased water flow (Larsen et al. 2008). Cladoceran density in autumn also evidenced distinct differences according to summer rainfall. In dry years, the number of cladocerans increased by more than 100 individuals; however, rainfall suppressed autumn zooplankton numbers in the rainy years. The dominant cladoceran species in the lower Nakdong River,
The results of our FA showed a complex relationship between zooplankton and environmental parameters. Factor 1, identified as the ‘phytoplankton growth factor,’ was the factor that primarily explained the limnological characteristics of the lower Nakdong River. Similar results have been reported in related studies using either statistical or machine learning Ecological Informatics methods (Kim at al. 2006). Factor 2, the zooplankton and hydrological variables, revealed that zooplankton groups were more profoundly affected by hydrological parameters than water temperatures. CC analysis revealed similar patterns, and the delayed influence of summer concentrated rainfall was evaluated. Nonetheless, the CC application results relate both summer rainfall to autumn zooplankton and winter scarce rainfall (low zooplankton density due to low temperature) to spring high zooplankton density, and the following regression analysis results reinforce the first relationship. Therefore, it can be concluded that summer rainfall distribution patterns perform important functions in determining autumn zooplankton community dynamics.
Based on the regression analysis, we can hypothesize that a possible relationship exists between summer concentrated rainfall and autumn zooplankton dynamics, as illustrated in Fig. 8. The multi-purpose dams in the upstream area would collect water from increased summer rainfall. The raised water level of the dams lead to an increase in dam discharge, which results in increased river flow in the lower part of the river during the following dry seasons (Jeong et al. 2007). The autumn zooplankton response to summer concentrated rainfall shown in this study can be explained by this relationship.
The other regression results of summer/autumn zooplankton density can be explained by the chance of population or community growth. If summer zooplankton groups are flushed due to summer concentrated rainfall, the total size of the zooplankton community would be reduced. The zooplankton community will grow when disturbance factors are removed; however, the population may require a longer recovery time (Altermatt and Ebert 2008). According to Levesque et al. (2010), the zooplankton density of a certain season is related to the density of the previous season. The high density of zooplankton in the autumn of a dry year may be attributable to the increased offspring production opportunity of the summer zooplankton group. In the Nakdong River, from the winter to the next spring (typically from December to next May), rainfall is scarce (Park et al. 2002) and temperature begins to increase, which enables zooplankton growth. If in a certain year a slight drought is experienced in the summer, there results an increase in the output of the food chain, which can persist from spring to autumn. However, recent summer rainfall (especially in August) has statistically increased over the last 3 decades (Chang and Kwon 2007), thus reducing the opportunity for this autumn food chain activation. Long-term climate variation and zooplankton community dynamics have to be related in order to evaluate and predict this phenomenon. In conclusion, then, we find that autumn zooplankton dynamics were affected by the summer concentrated rainfall, directly by density dilution, and indirectly by a reduction in population size.