A central pattern in ecology is the interaction of species in many different ways with each other; they can compete,be mutual exclusive or can occur together and often depend on each other (Begon et al. 1996). Since decades co-occurrence has been used to identify indicator species which are applied in terms of conservation biology(Pearson and Cassola 1992, “umbrella species”), habitat descriptions (Aydin and Kazak 2010) and in biodiversity surrogacy. Several studies showed that unrelated taxa can act as surrogates for one another to predict biodiversity.Using different taxa of plants, insects and vertebrates,Leal et al. (2010) showed that this is true for different biomes.In a similar approach Schulze et al. (2004) revealed certain taxa as good predictors for species richness of other taxa, for example trees for fruit- and nectar-feeding birds. In concordance with this, Ricketts et al. (2002)found that phylogenetic relatedness is no reliable criterion for selection of suitable indicator taxa since butterflies were found to be poor indicators of moth diversity at a local scale. Instead, using unrelated taxa in terms of biosurrogacy seems to be more convenient. For instance, Blair (1999) demonstrated that birds and butterflies can be used as surrogates for one another in assessing biodiversity at the community level within a single habitat of a former oak wood.
Birds and plants provide an interesting example for studying biosurrogacy and indicator groups for one another since both taxa often rely on each other. Many bird species feed on plants and build their nests in trees, shrubs or grassy vegetation. In turn, plants can profit since their seeds are dispersed by migrating birds (Howe and Smallwood 1982). The co-occurrence between these two unrelated taxa can be very tight as demonstrated by Howe (1977) who showed that the extinction of a certain tree species led to the disappearance of the bird species feeding on it. Bird species assemblages were also used as ecological indicators of forest condition showing that certain bird species were typical for disturbed habitat or undisturbed habitat in a pine forest in northern America (Canterbury et al. 2000). Further examples from Martin and Possingham (2005), Barry et al. ( 2006) or Lehmkuhl et al. (2007) showed that studies focused so far only on forest types with certain dominating tree species inhabiting characteristic bird species in few cases. However, to our knowledge no study has ever before revealed certain bird and plant species predicting the presence and absence of a given bird species.
In this study we recorded the companion bird and plant species of the endemic breeding bird Cyprus Wheatear (
Field work was carried out on the Mediterranean island Cyprus from 22 March 2008 to 21 April 2008 during spring bird migration and totalled > 300 h. Despite its relatively small size of approximately 9,250 km² the island has a high diversity of natural vegetation and nearly 20% of Cyprus is covered with woodland (Stagg and Hearl 1998, Jones 2006). When performing field observations, we walked around randomly and searched for individuals of the target species (see Salewski et al. 2003). We assessed the habitats within a radius of 100 m for the target species Cyprus Wheatear using the main location of a singing male. Control habitats were also measured with 100 m radius. They were assigned when there was no Wheatear present. Here, playback experiments were used to ascertain the absence of a Cyprus Wheatear because it responds strongly to conspecific playbacks. In migrant wheatear habitats, we chose a radius of 25 m because migrants often used only a very small area, and extending the 25 m radius would have included vegetation structure that was not used by the migrants. Furthermore, preliminary observations in 2005 (by CR) indicated that 100 m for breeding habitat and 25 m for migrant
We sampled a total of 192 habitats, 129
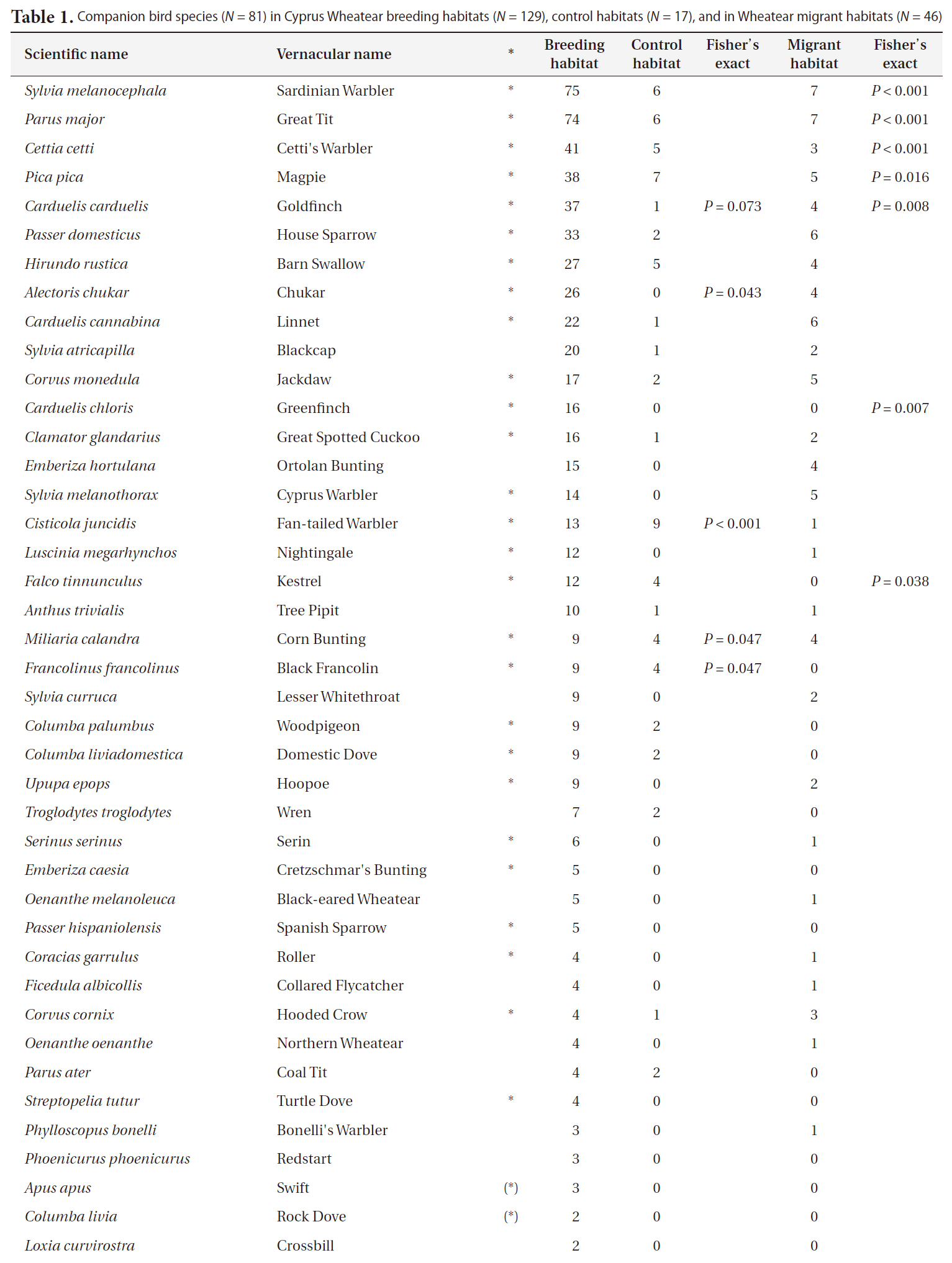
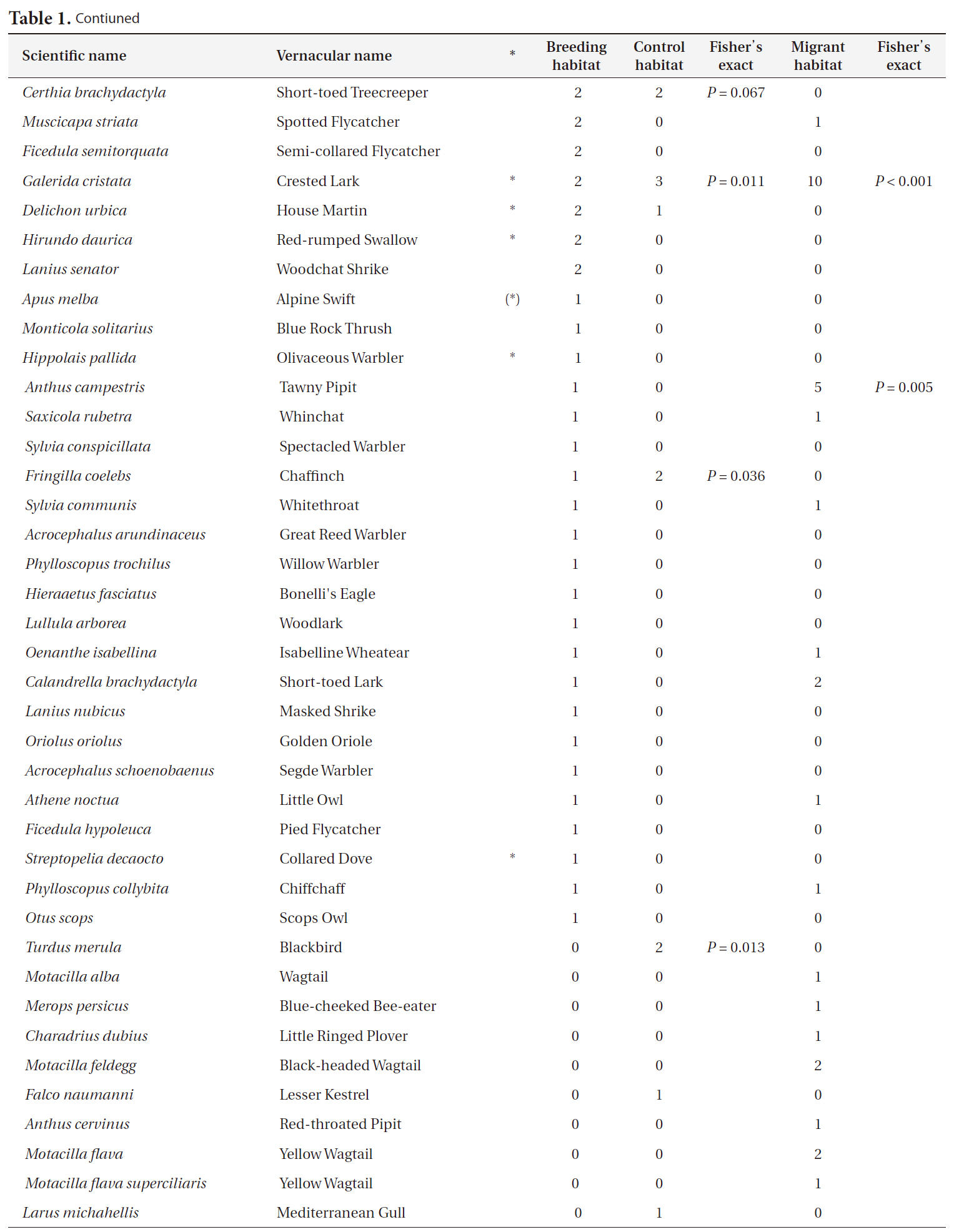
Companion bird species (N = 81) in Cyprus Wheatear breeding habitats (N = 129) control habitats (N = 17) and in Wheatear migrant habitats (N = 46)
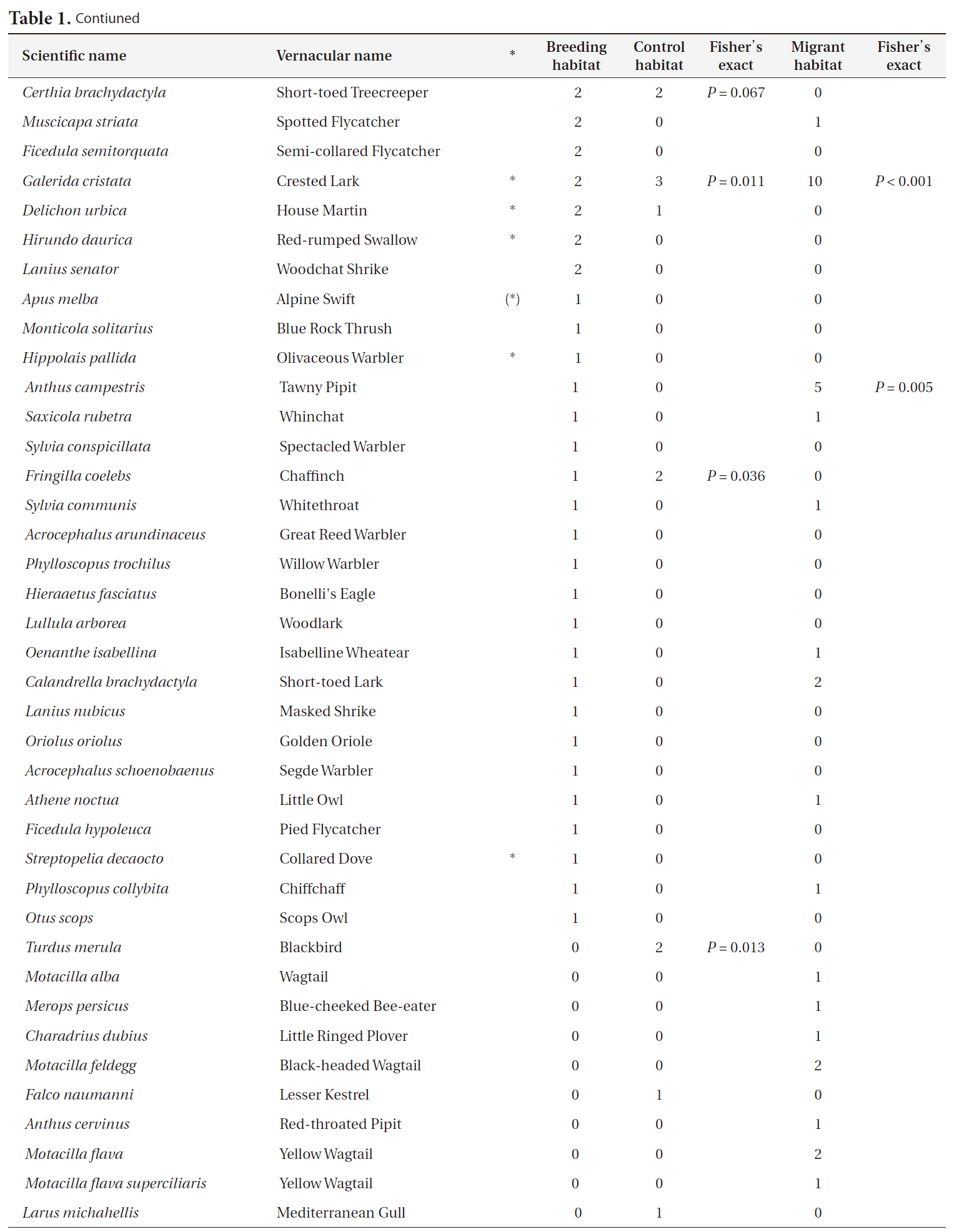
Contiuned
and Goldfinch
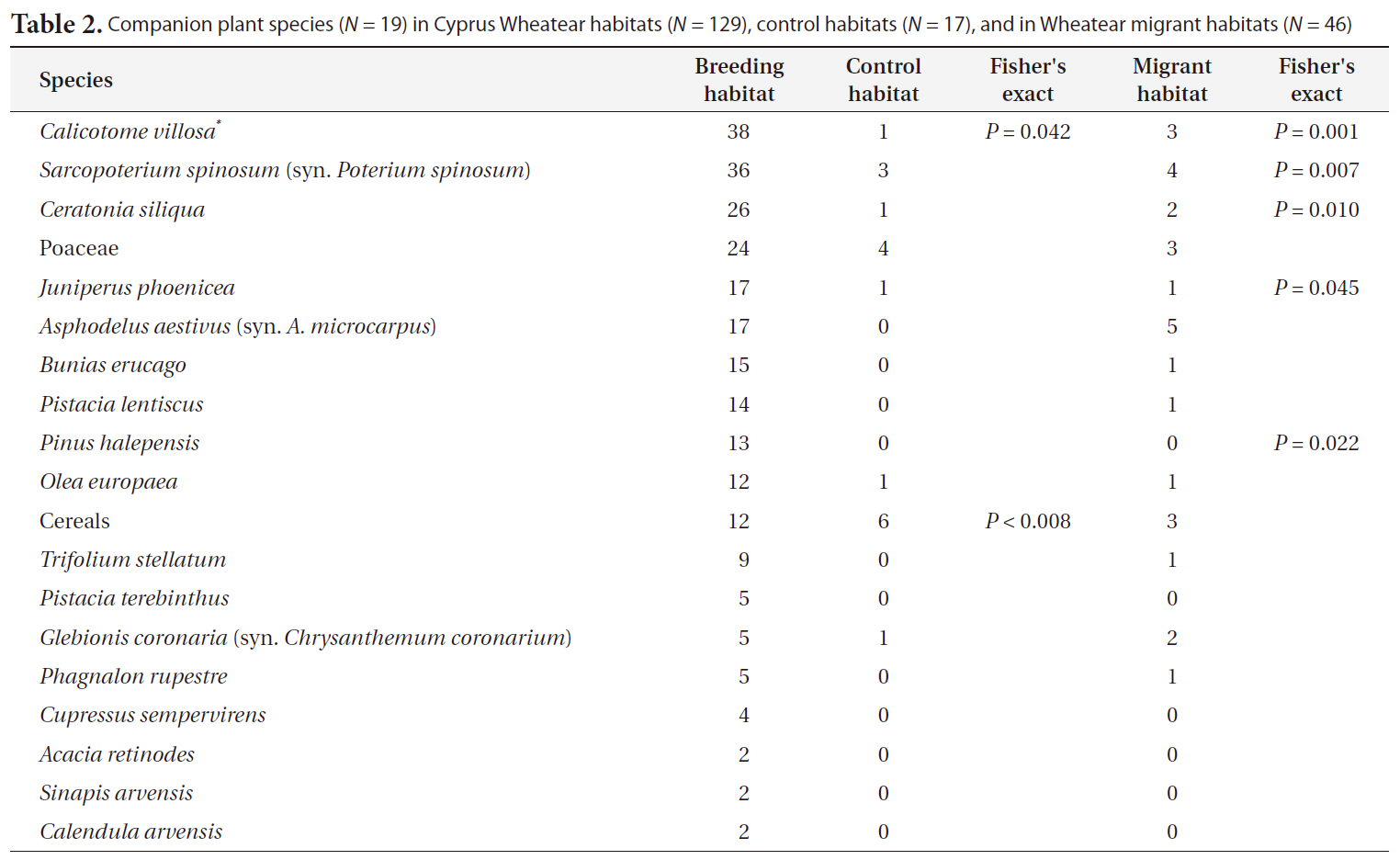
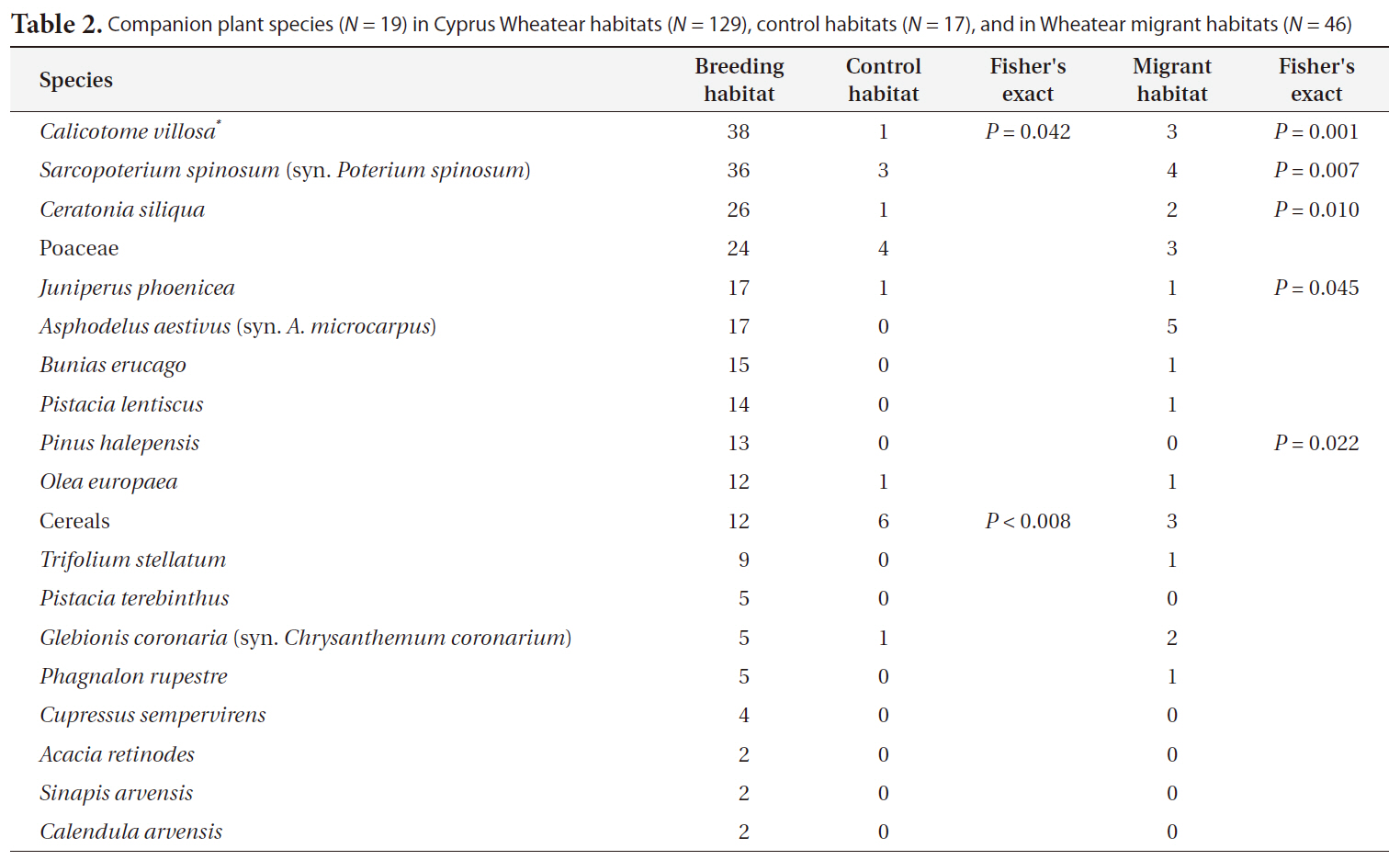
Companion plant species (N = 19) in Cyprus Wheatear habitats (N = 129) control habitats (N = 17) and in Wheatear migrant habitats (N = 46)
Co-existence of species is a basic pattern of ecosystems. The degree of their phylogenetic relationships may vary. Analysing co-occurrence of unrelated taxa in a certain area has been applied to questions in biosurrogacy and conservation biology mostly with a focus on insects, vertebrates and plants. For example it was shown that insect species can be used as bioindicators for habitats like afforested
In this study we used birds and plants as the experimental system. In the field, both taxa have the advantage that they are relatively easy to detect and to identify and their taxonomy is well resolved (Diekmann 2003, Gregory et al. 2005). We found that the habitat preferences of the companion bird species of the Cyprus Wheatear indicate a habitat dominated by bush land vegetation and low distances to settlements. Likewise the dominating plant species indicate a preference for bush and tree vegetation. Control habitats were clearly separated from the habitats of Cyprus Wheatear by their companion bird species. There was also a difference in the dominating plant species between these two habitats. Cereals were typical for control habitats as indicator species in contrast to Cyprus Wheatear habitats. This indicates that the Cyprus Wheatear avoids woodlands on the one side and intensively managed and even farmland on the other. The habitats of migrating Wheatear species could also be distinguished from habitats of Cyprus Wheatear by their companion species. For example Tawny Pipit
Thus, we provide evidence that the presence or absence of a bird species in a given habitat can be determined by its bird and plant companion species. We showed that companion species ? birds as well as plants ? can be used as predictors for the presence or absence of the Cyprus Wheatear in a certain area. Furthermore, it was possible to identify control habitats and habitats of migrating Wheatears based on their bird and plant companion species.
In a more general context, it might be useful to determine the presence or absence of a species based on its companion species especially when the target species is difficult to localize due to camouflage or rare performance of bioacoustics. In addition, this approach shown here obviates the need for a time-consuming habitat analysis.