Males of the field cricket Teleogryllus emma produce calling songs that are attractive to receptive females. The calling songs of T. emma consist of two components, the long chirp that is composed of up to 12 single pulses, followed by a variable number of short chirps. Based on the analysis of coefficient of variation (CV), temporal characters of the long chirp were less variable than those of the short chirps in male calling songs. To test for phonotactic selectivity of females,we conducted a single-stimulus playback experiment in which five stimuli (standard, long chirp only, long chirp augmented,short chirps only, and short chirps augmented) were used. The standard stimulus included both long and short chirps whose characteristics were derived from the calling songs of field populations. Results of the playback experiment showed that female crickets oriented more frequently toward the stimuli that included the long chirp (standard, long chirp only, and long chirp augmented stimuli) than toward the stimuli lacking the long chirp (short chirps only and short chirps augmented stimuli), indicating that the long chirp in the calling songs was required to elicit positive phonotaxis in the female crickets. The result of CV analysis of the male calling songs and the findings of the female phonotaxis experiment suggested that the long chirp of calling songs may play a role in species recognition in T. emma.
The mating behavior of a cricket species typically consists of production of male calling song and female phonotaxis(Alexander 1962, Huber and Thorson 1985, Zuk and Simmons 1997). The primary function of such longrange mating behavior is that the male and female of a species come together to mate. Upon coming together,the male and female engage in close-range mating behaviors including antennal contact, courtship song, and mounting (Loher and Dambach 1989, Adamo and Hoy 1994, Jang et al. 2007). Acoustic communication studies,especially on species in areas of sympatry, have revealed that the long-range calling songs of many crickets are species-specific (Alexander 1962, Gerhardt and Huber 2002, Greenfield 2002). Furthermore, females prefer the songs of conspecifics over those of heterospecifics(Doherty and Callos 1991, Hennig and Weber 1997, Gray and Cade 2000). However, calling songs can also be used in the context of sexual selection, in that females discriminate among males on the basis of certain properties of calling songs for mating (Ritchie et al. 1995, Price 1998, Ferreira and Ferguson 2002, Klappert and Reinhold 2003).
The calling song characters primarily used for species recognition are less likely to vary within and among individuals.The female preference functions associated with species recognition may be stabilizing or “unimodal,”such that females prefer males with intermediate values of calling song characters (Butlin et al. 1985, Ewing and Miyan 1986, Ferreira and Ferguson 2002, Klappert and Reinhold 2003, Saarikettu et al. 2005). When other calling song characters are used for mate selection, however, the intermale variability of these characters is typically higher than that of calling song characters used for species recognition. Female preference functions associated with the mate selection context are frequently directional or “open-ended,” such that females prefer males with extreme values of calling song characters (Ritchie 1996, Shaw and Herlihy 2000).
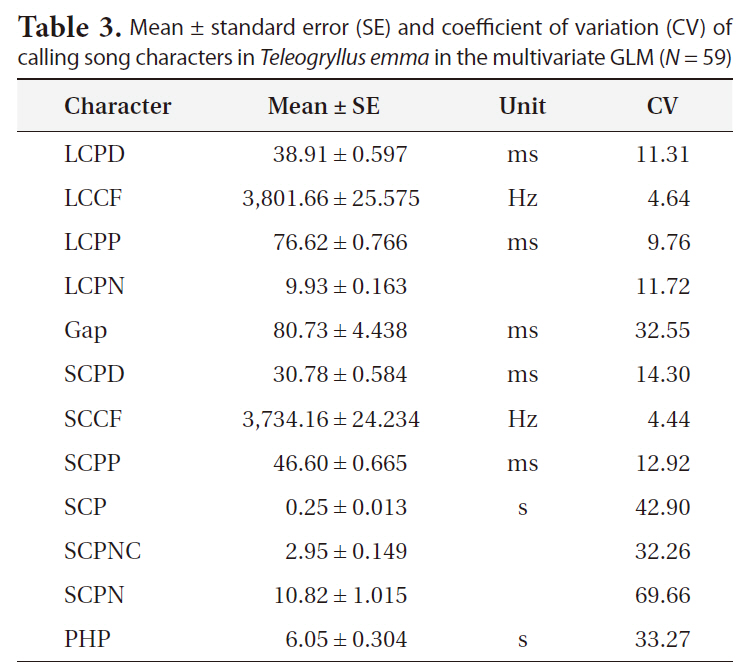
Based on within-male variability, which was expressed as the coefficient of variation (CV), Gerhardt (1991) identified static and dynamic characters in male advertisement signals. Static characters change relatively little within and between males in natural populations, and the CV values for static characters are typically lower than 5% (Gerhardt 1991). On the other hand, dynamic characters whose CV values are higher than 12% are more variable within males and between males. The expectation that selection from female choice is stabilizing or weakly directional for static characters and is directional for dynamic characters has been largely confirmed in the North American tree frog species
The Emma field cricket,
The habitats of
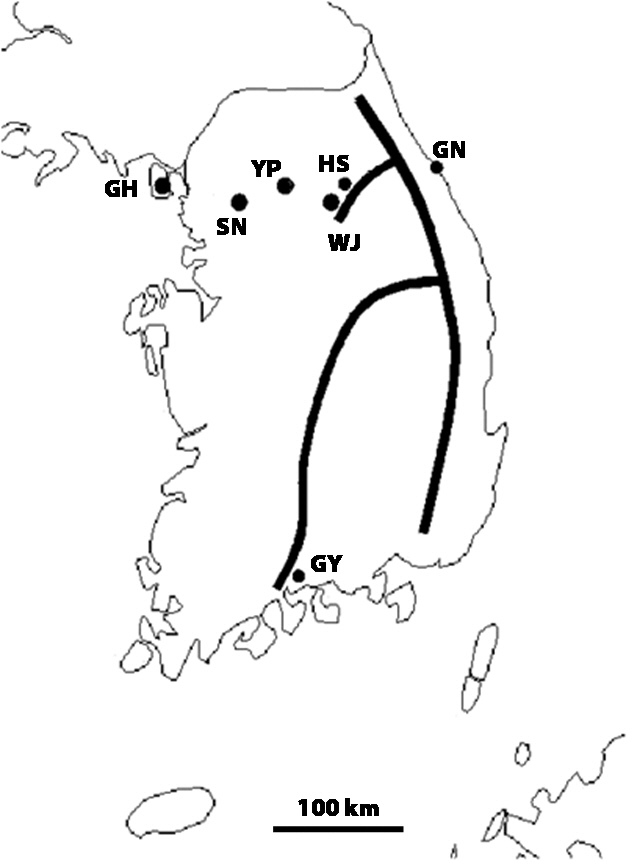
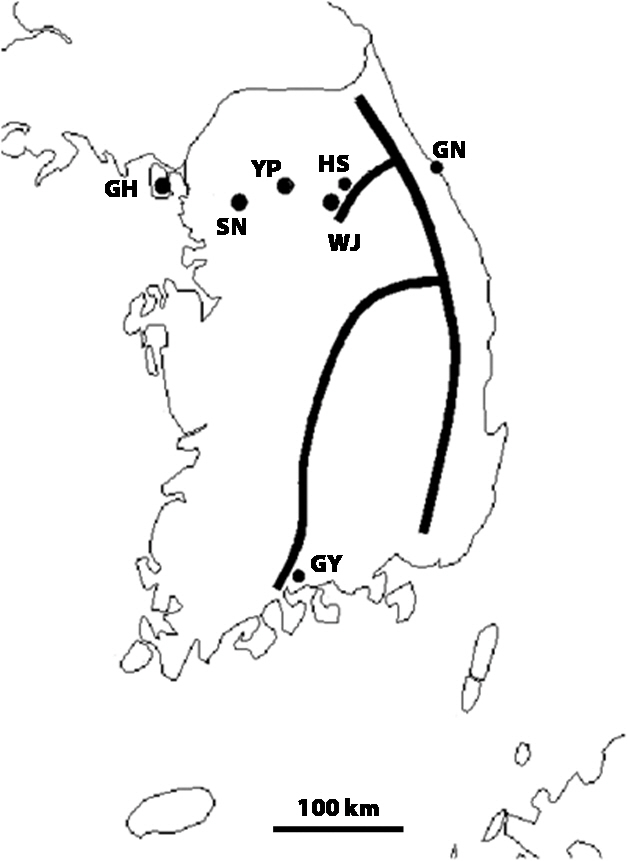
Incheon (GH, 9), Seongnam, Gyeonggi-do (SN, 8), Yangpyeong, Gyeonggi-do (YP, 10), Hoengseong, Gangwon-do (HS, 4), Gangneung, Gangwon-do (GN, 17), Wonju, Gangwon-do (WJ, 12), and Gwangyang, Jeollanam-do (GY, 5) (Fig. 1). The parentheses above are abbreviations of the locality and the total number of recordings for that locality. One male cricket contributed to only one recording. Calling songs were recorded in the field using a tie-clip microphone (TCM 110; In Young Electron Co., Ltd., Taipei, Taiwan; flat frequency response from 50 to 18,000 Hz). We placed the microphone as close as possible to a singing male and recorded his calling song for more than 50 s. Output from the microphone was fed into a DAT recorder (PCM-M1; Sony Corp., Tokyo, Japan) at a sampling rate of 44.1 kHz. The temperature near the calling male was noted using a HOBO data logger (H08-004-02; Onset Computer Corp., Pocasset, MA, USA).
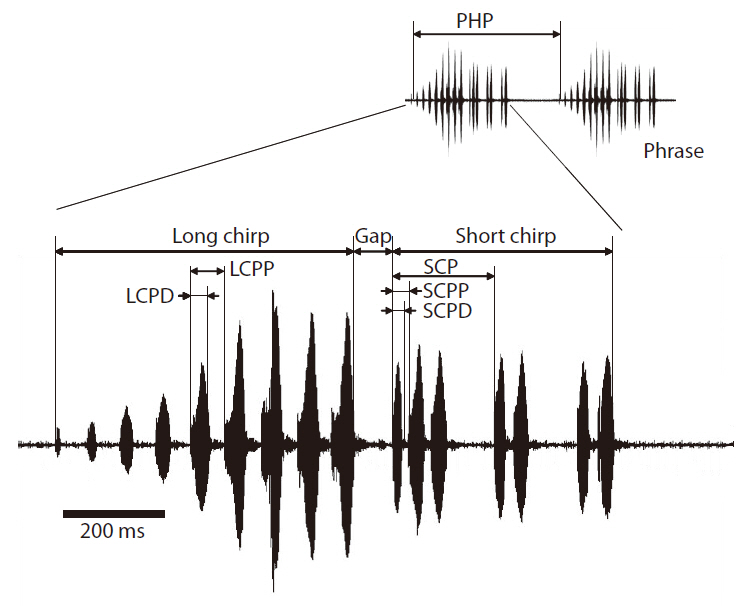
Cricket calling songs were analyzed using Raven ver. 1.2 software (Cornell Laboratory of Ornithology, Ithaca, NY, USA). Frequencies below 1 kHz were filtered out to reduce background noise. We analyzed 12 consecutive phrases from each recording for statistical analyses. The calling song of
[Table 1.] Definitions of calling song characters of Teleogryllus emma
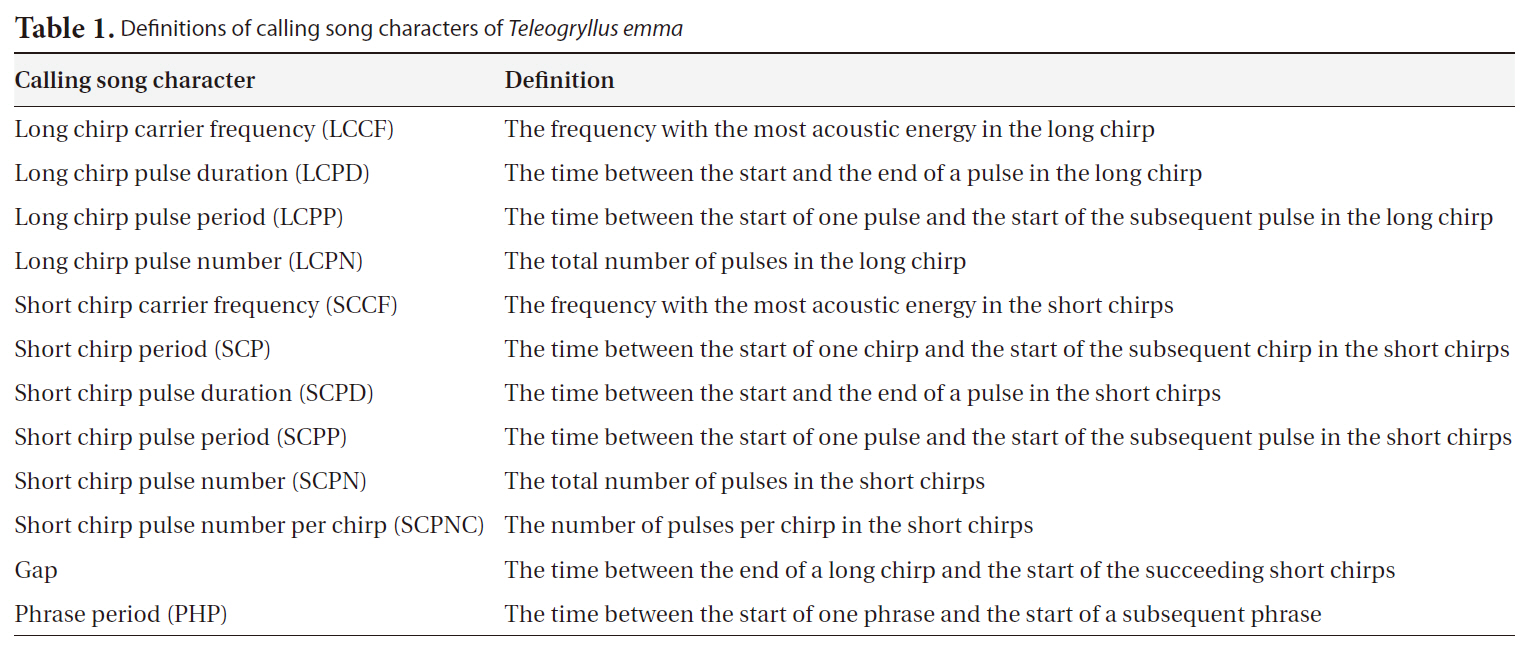
Definitions of calling song characters of Teleogryllus emma
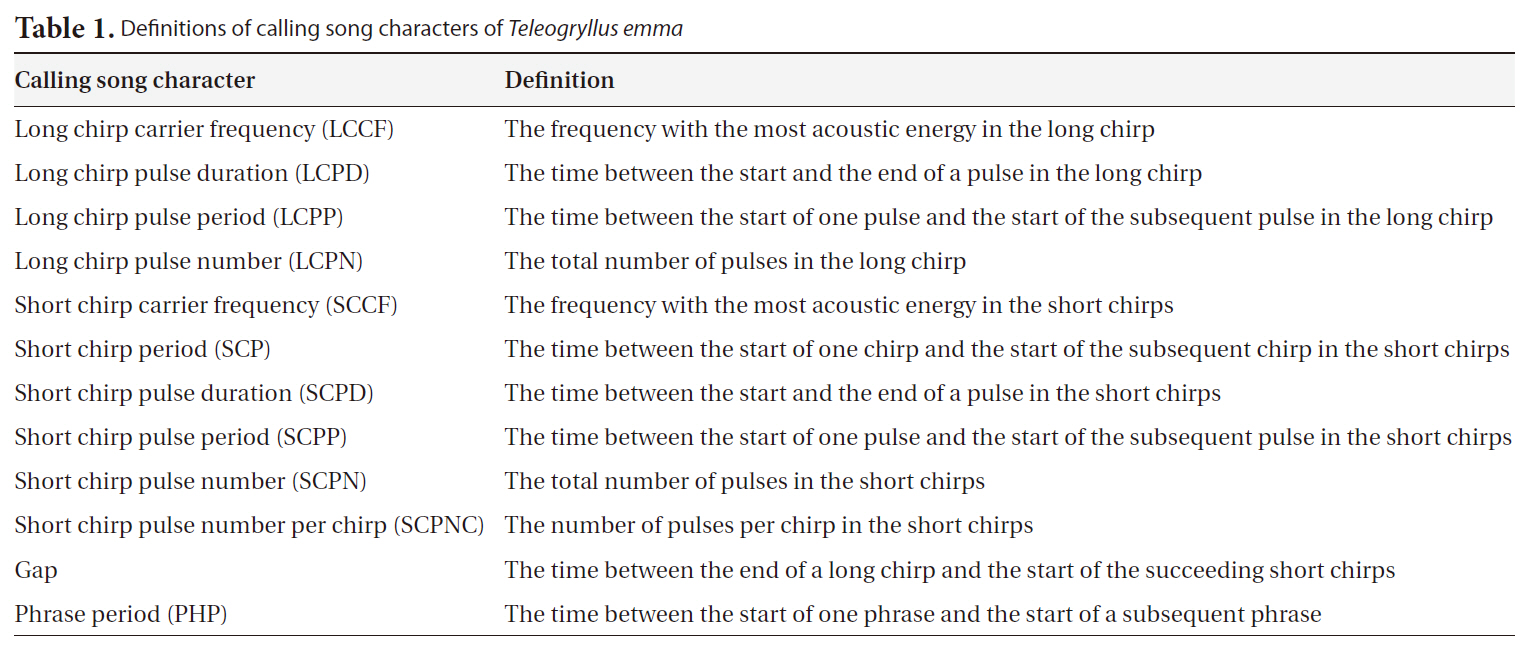
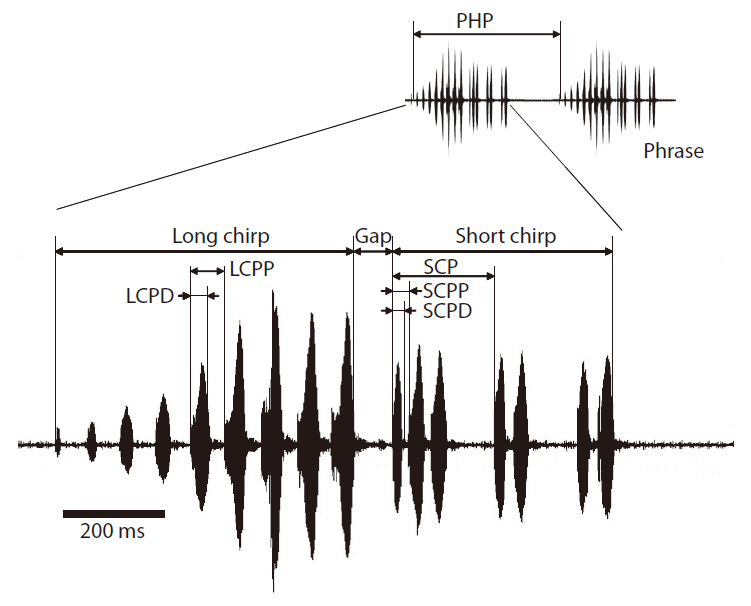
short chirp carrier frequency (SCCF), short chirp pulse duration (SCPD), short chirp period (SCP), short chirp pulse period (SCPP), short chirp pulse number per chirp (SCPNC), short chirp pulse number (SCPN), Gap, and phrase period (PHP). See Fig. 2 and Table 1 for definitions of these calling song characters. LCCF and SCCF were spectral characters, and all others were temporal characters. The definitions of calling song characters follow Bennet-Clark (1989).
For the phonotaxis experiment, final-stage juvenile crickets were provided by the Department of Agricultural Biology of the National Academy of Agricultural Science in Korea, which had maintained the stock population of T. emma for more than 2 y. Crickets were reared in an environmental room (2 × 2.5 × 2.5 m) that was maintained at 23.5 ± 1.5°C and 75% relative humidity, with a 12 h light:12 h dark photoperiod. The juveniles were housed in plastic containers with cat chow and water. Adult crickets were removed from the colony within 48 h of the final molt to ensure that all crickets used for this study were virgin. Newly emerged adults were individually housed in plastic containers (12 × 12 × 9 cm) with cat chow and vegetables. Both juveniles and adults were provided with egg cartons for shelter. Age was calculated as the number of days after the final molt.
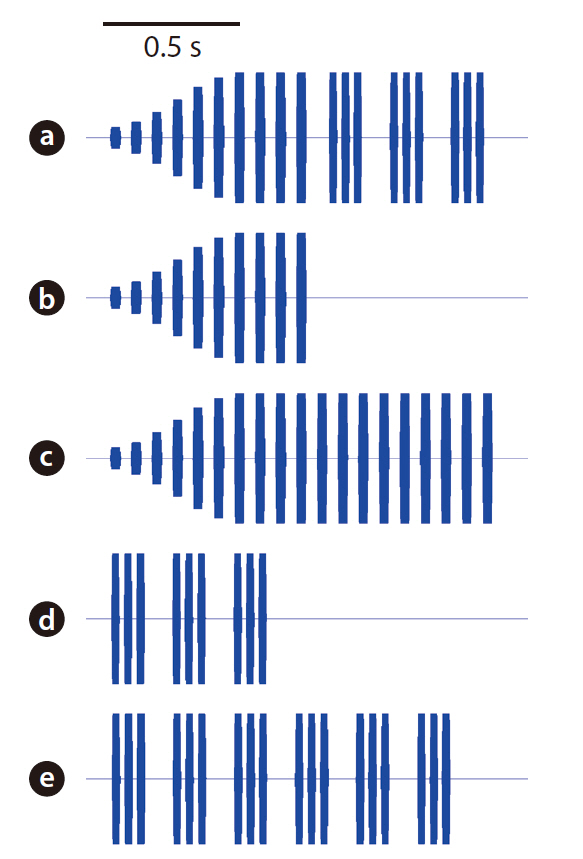
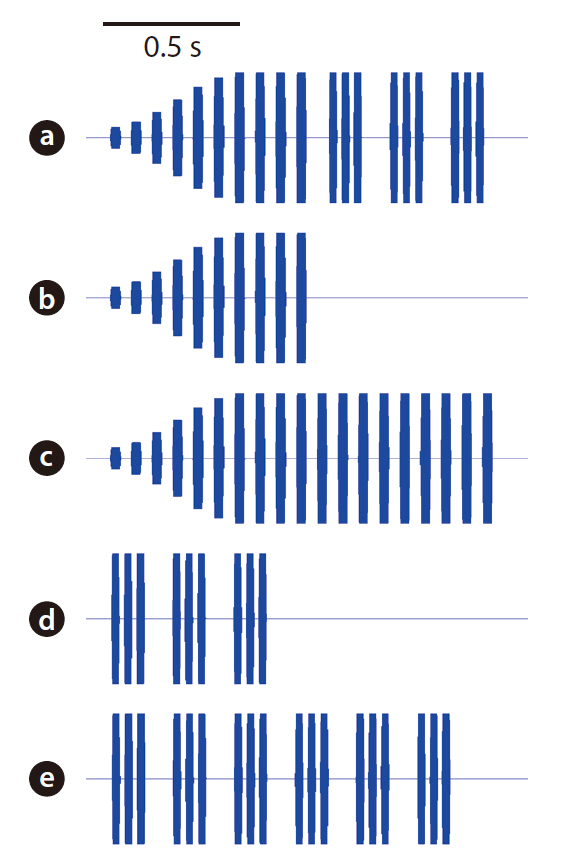
The acoustic stimuli used for the playback experiment were as follows: 1) standard, 2) long chirp only, 3) long chirp augmented, 4) short chirps only, and 5) short chirps augmented (Fig. 3). They were synthesized using custom-designed software with 16-bit resolution and 44.1 kHz sampling rate (Schul and Bush 2002). The standard stimulus included both long and short chirps, the characteristics of which were derived from the calling songs of field-caught males (LCPD: 37.95 ms; LCCF: 3,750 Hz; LCPP: 74.86 ms; LCPN: 10; Gap: 74.66 ms; SCPD: 29.48 ms; SCCF: 3,750 Hz; SCPP: 43.64 ms; SCPN: 3; SCP: 0.22 s; PHP: 6.53 s). The last four pulses of the long chirp and all pulses in the short chirps were at a constant amplitude. However, the amplitude of the first six pulses of the long chirp was modulated, with the magnitude of the amplitude increasing sequentially relative to the rest of the song as follows: 0.14, 0.29, 0.43, 0.57, 0.71, and 0.86. The “long chirp only” and “short chirps only” stimuli were the standard stimulus without the short chirps and long chirp, respectively. Female phonotactic responses may be biased in the long chirp only and short chirps only stimuli, because these stimuli had fewer numbers of pulses than had the standard stimulus. Thus, we eliminated this possibility by introducing two augmented stimuli whose pulse numbers were equivalent to that of the standard stimulus. The “long chirp augmented” stimulus was an elongated long chirp only stimulus, with 19 pulses instead of 10. Likewise, the “short chirps augmented” stimulus was an elongated short chirps only stimulus with 18 pulses instead of 8.
The playback experiment was conducted in a semi-anechoic chamber (2.6 × 1.7 × 2.4 m). Playback trials were run in an octagonal arena with a 50 cm diameter. The arena was constructed on a plywood base covered with a thin layer of carpeting. The boundary of the arena was a 13-cm high acoustically transparent screen. The stimulus was played back using a CD player (Denon DN-C630; Nippon Columbia Co., Ltd., Tokyo, Japan), and the output from the CD player was sent to an amplifier (No. 1700; QSC Audio, Costa Mesa, CA, USA). The signal from the amplifier was then sent to one of two tweeters (Dayton No. 275-100; Euro series textile tweeter, Springboro, OH, USA; flat frequency 2,000-20,000 Hz). The two tweeters were placed at floor level outside the arena wall (55 cm from the center of the arena), angled at 120°. The choice of a tweeter for playback trials was switched daily to prevent any phonotactic bias that might be present in the arena. The amplitude of a stimulus played back from a tweeter was calibrated to 80 ± 0.5 dB SPL (0 dB = 20 μPa) using a Bruel and Kjær sound level meter (No. 2250; Bruel and Kjær, Nærum, Denmark). During the trials, the anechoic chamber was maintained at 23.5 ± 1.5°C in complete darkness.
Female crickets to be tested were drawn haphazardly from the stock population after eclosion. Only intact female crickets between 7 and 33 days of age were used.
Each trial was begun by placing a female cricket under an opaque plastic cover located in the center of the arena. The plastic cover (250 mL) had holes for sound permeation. The cricket was placed in the arena for 1 min without any sound to acclimate and was then exposed to the test stimulus broadcast from one of the tweeters for 1 min. The cricket was released by removing the cup. Female response to the stimulus was monitored using an IR camera (DSP No. HN8071R UC3; Seoul, Korea; minimum illumination: 0 Lux when IR LED on). Trials were terminated after 5 min or when the test females evidenced positive phonotaxis. A positive phonotactic response was recorded if a female oriented itself toward the tweeter and touched the wall of the arena directly in front of the loudspeaker broadcasting the test stimulus, or if a female made a ‘double reversal’ movement in front of the tweeter (Jang and Gerhardt 2006). That is, crickets sometimes ran toward the arena screen after release without orientation and then walked along the screen. A female cricket occasionally reversed its walking direction in front of the tweeter broadcasting the test stimulus twice in a row within close proximity of the tweeter; each reversal of direction had to cross the line between the tweeter and the female release point. The double reversal movement was a local search, and occurred when an individual was very close to the object for which it was searching (Bell 1991). Each cricket was tested only once per day, and was tested a total of five times with five test stimuli. The order in which the stimuli were presented was determined using a random number table. A total of 30 female crickets completed five stimuli in the playback experiment.
After completing the playback experiment, female crickets were measured once for head width, thorax length, and hind femur length to the nearest 0.05 mm using a digital Vernier caliper. Head width was measured as the distance between the outer edges of the compound eyes. Thorax length was the distance between the anterior and posterior ends in the midline of the thorax. A right hind leg was removed from the thorax and placed on its side. Hind femur length was measured from the base of the femur to the joint with the tibia.
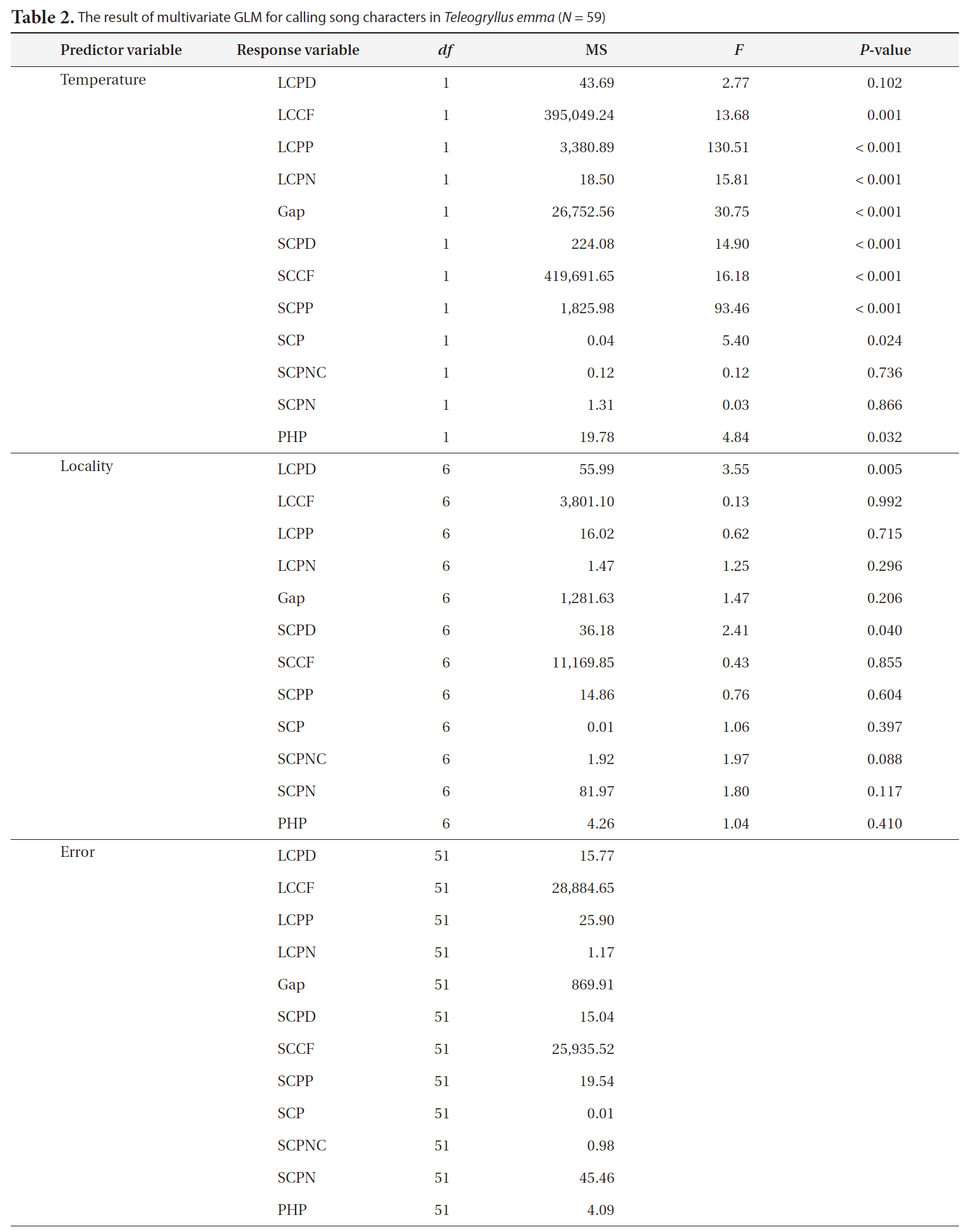
The multivariate general linear model (GLM) was used to evaluate the effect of locality on the calling song characters of
>
Variation in calling song characters
The results of multivariate GLM showed that locality was not a significant factor for any of the calling song characters, with the exceptions of LCPD and SCPD (Table 2). Temperature significantly affected most calling song characters, except for LCPD, SCPNC, and SCPN (Table 2). Carrier frequencies of both long chirp and short chirps whose CV values were less than 5% showed the lowest levels of variation (Table 3). The CV values for the temporal characters of the long chirp ranged between 9.76 and 11.72%, whereas the CV values for the temporal characters of the short chirps were greater than or equal to 12.92%. Calling song characters related to short chirp pulse number, SCP, SCPNC, and SCPN, had the most variation. Thus, the carrier frequency of calling songs may be a static character, whereas the temporal characters of the short chirps were dynamic characters in
Body size was estimated on the basis of the PCA for head width, thorax length, and hind femur length. We identified only one principal component (PC) with an eigenvalue greater than 1, and this PC explained 79.8% of the total variation in female body size. Thus, PC1 was
[Table 2.] The result of multivariate GLM for calling song characters in Teleogryllus emma (N = 59)
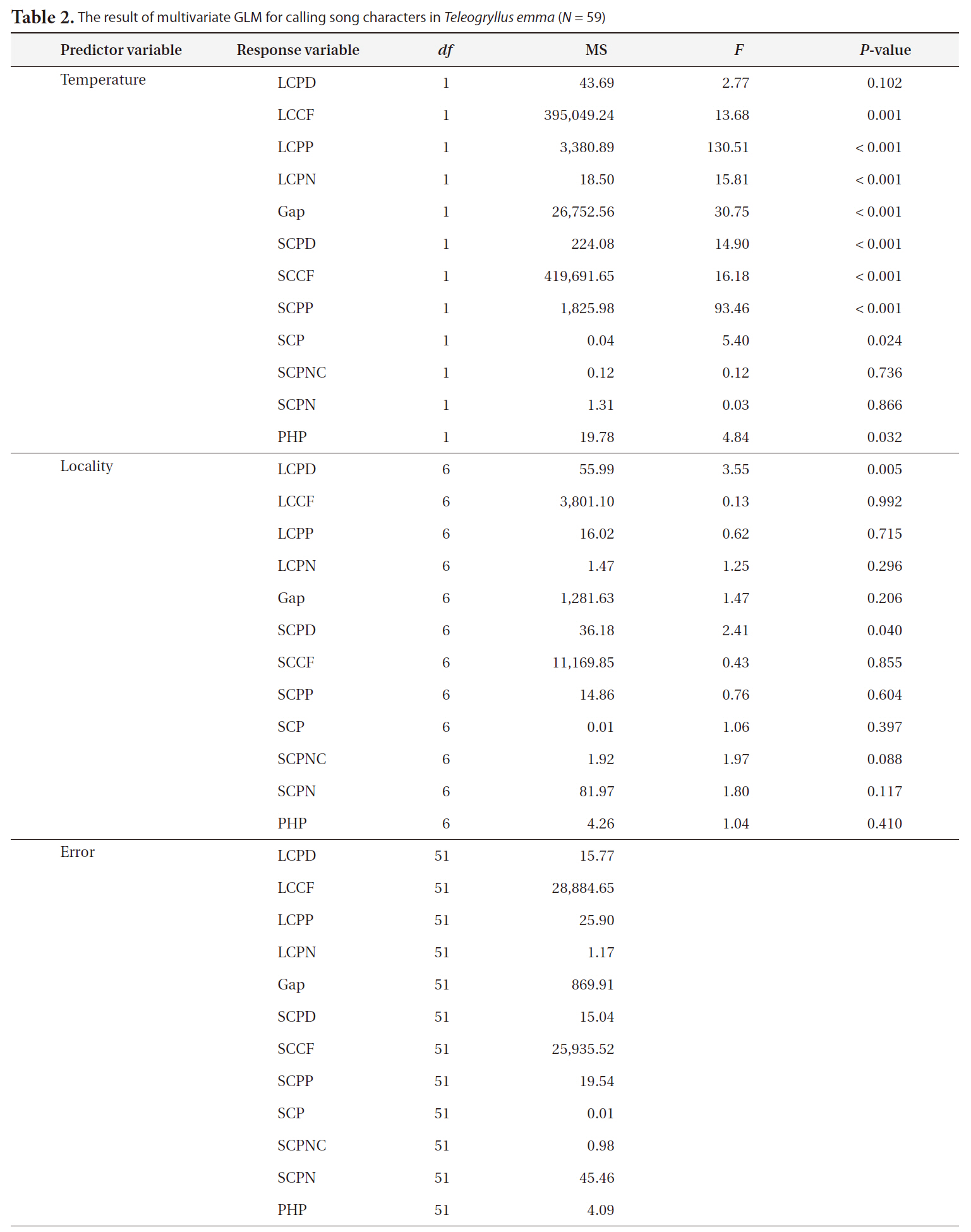
The result of multivariate GLM for calling song characters in Teleogryllus emma (N = 59)
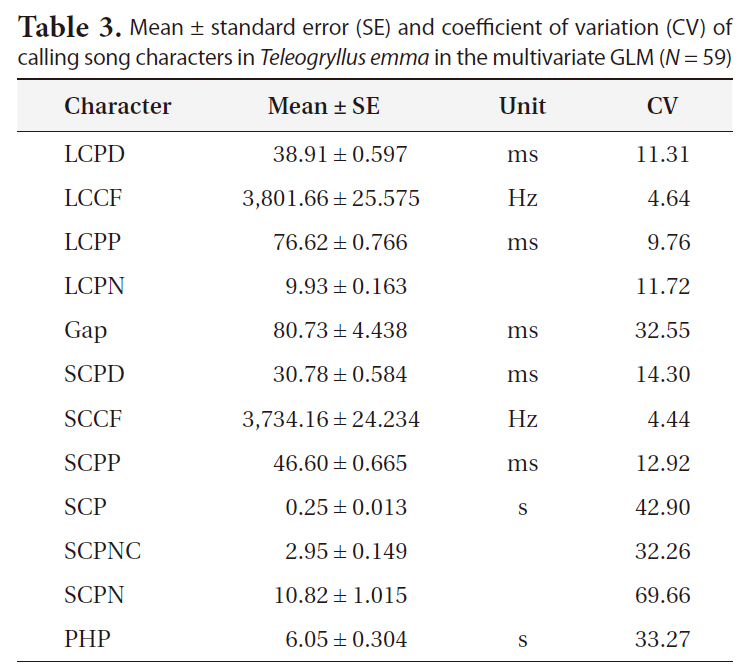
Mean ± standard error (SE) and coefficient of variation (CV) of calling song characters in Teleogryllus emma in the multivariate GLM (N = 59)
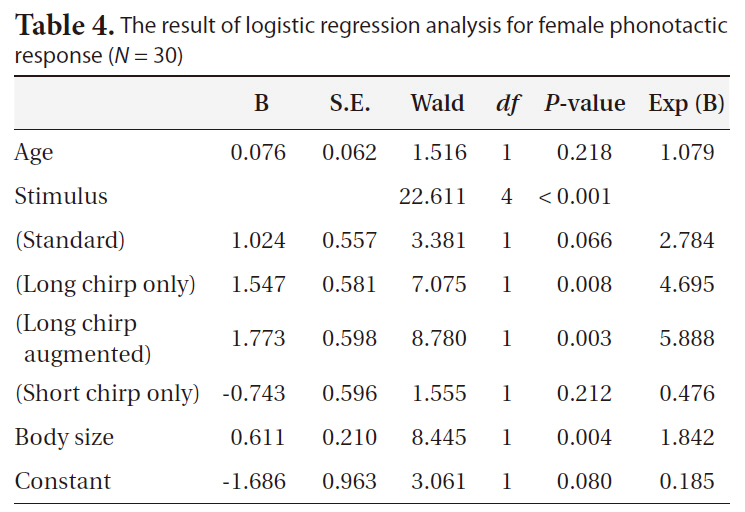
[Table 4.] The result of logistic regression analysis for female phonotactic response (N = 30)
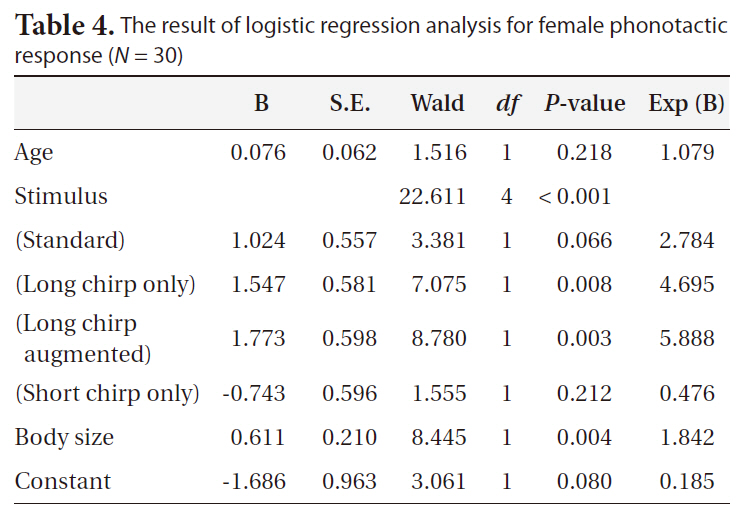
The result of logistic regression analysis for female phonotactic response (N = 30)
used as the body size of females tested for phonotactic trials.
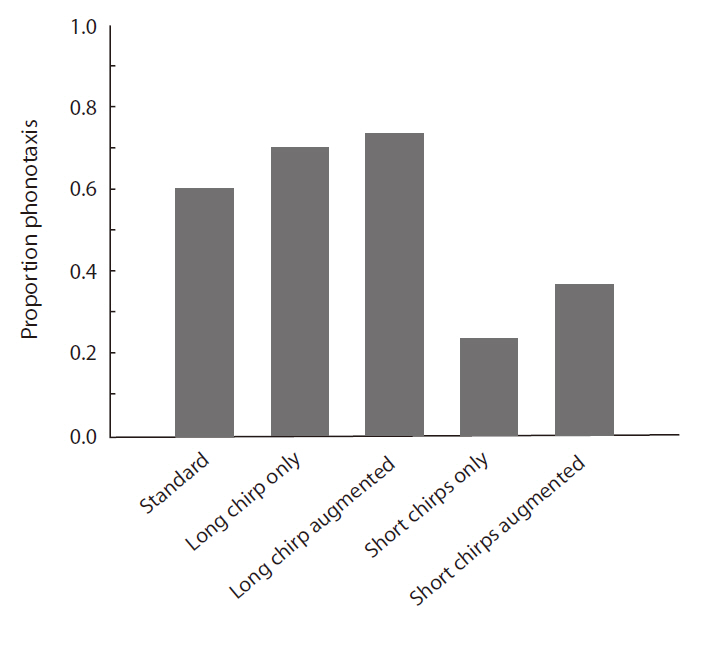
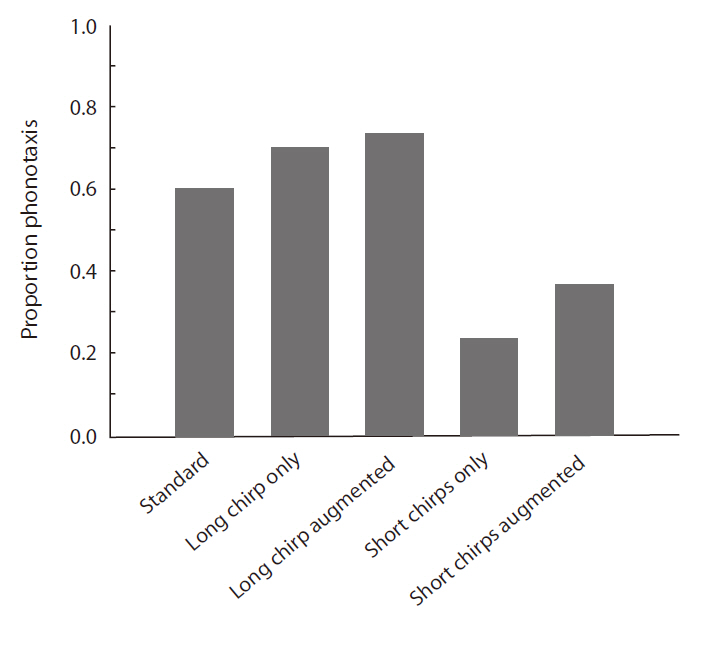
When the standard stimulus was played back, 60% of the females oriented toward it (Fig. 4). The proportions of positive female phonotaxis toward the long chirp only and the long chirp augmented stimuli were greater than that of the standard stimulus. However, the proportions of positive female phonotaxis toward the short chirps only and the short chirps augmented stimuli were lower than that of the standard stimulus. The result of logistic regression analysis revealed that positive female phono-
taxis was affected significantly by stimulus and body size (Table 4). The proportion of female phonotactic orientation increased with increases in body size.
Although the findings of this study revealed that there was almost no geographic variation in the calling song characters, such findings may have been the results of insufficient sampling. The majority of the sampling localities were limited to a narrow range of latitudes. To conclusively address geographic variation in the calling songs of
Based on the results of our CV analysis, the carrier frequencies of both the long chirp and short chirps were identified as a static character in the male calling songs of
The temporal characters of the short chirps were dynamic, suggesting that these characters were unlikely to be relevant to species recognition in
The CV values for temporal characters of the long chirp were lower than 12%, suggesting a potential role in species recognition in
Species recognition sensitively tuned to the properties of male calling songs is a matter of critical importance for