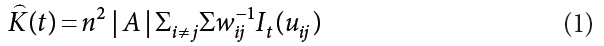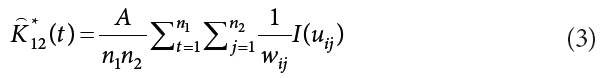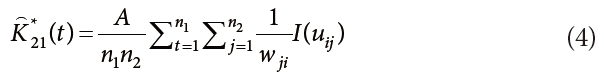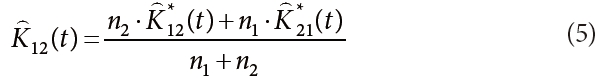Knowledge about the spatial relationships amongspecies in a stand can help researchers to understandinter- and intraspecific relationships and to developmanagement plans that mimic natural processes (Salaset al. 2006). Accordingly, the spatial distribution patternsof species are a topic of interest among ecologists andforest managers (Anderson 1992, Yang and Kim 2002,Monzeglio and Stoll 2005). With the increasing use ofstatistics in biology, many methods for interpreting thespatial distributions of species have been introducedand developed, such as the Clark and Evans (1954) test,Pielou’s (1960) index of non-randomness, the contiguousquadrat technique (Greig-Smith 1983), and Morisita’s(1962) index (Haase 1995).
Species in a stand compete with each other for resources such as light, soil moisture and nutrients. Species that require similar resources and similar environmental conditions have a tendency to be distributed in similar locations. However, the competitive exclusion principle (Hardin 1960) suggests that two species competing for the exactly same resources cannot coexist indefinitely, implying that species coexistence may indicate physical or temporal differentiation of the food resources used that reduces interspecific competition below the value of intraspecific competition, or that morphological differences among coexisting species enable the species to utilize different resources or slightly different types of environments.
Quercus mongolica Fisch. ex Ledeb. is a representative deciduous species in temperate forests in Northeast Asia. Q. mongolica is widely distributed across Korea and dominates Korean deciduous forests, especially on ridges and upper slopes. The topographical distribution of Q. mongolica on upper slopes and ridge areas may indicate that this species is adapted to dry soils and cold weather conditions. Q. mongolica individuals live for 100-300 years, and dominate the upper canopy layer (Abrams et al. 1999). They regenerate both by seeds and sprouts. In South Korea, more than 70% of Q. mongolica seedlings in natural forests were found to have originated from sprouts (Suh and Lee 1998). Their ability to sprout enables them to quickly reoccupy areas after catastrophic disturbances, especially fires, while their acorns are favored animal foods and make a relatively low contribution to Q. mongolica emergence. Although sprouting allows the species to reoccupy an area faster than other species due to its rapid early growth after a disturbance, it limits the spatial dispersion of the species to within the boundaries of the root distribution and competition with neighboring species has a substantial influence on Q. mongolica survival. Therefore the spatial relationships between Q. mongolica and neighboring species are important factors in Q. mongolica stands, especially stands with trees from sprout origin. However, little information is available about the spatial relationships of Q. mogolica with neighbor species despite the fact that Q. mongolica in Korean forests has been a major research subject. Our study aims to examine stand structural characteristics and the spatial relationship of Q. mongolica with neighboring woody species and to estimate the probability of persistence of this species in the study areas.
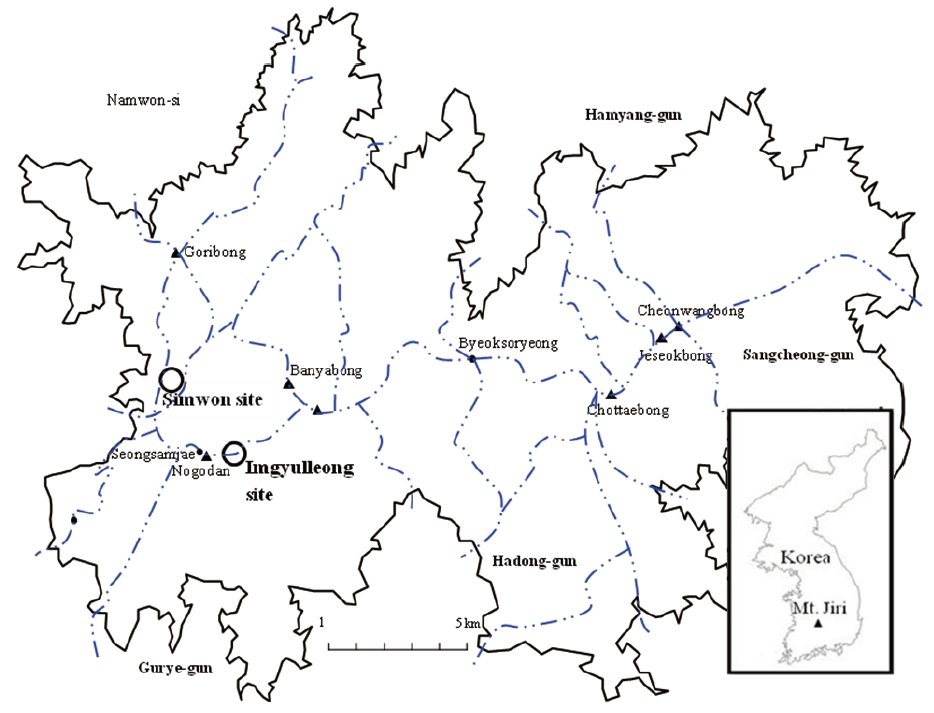
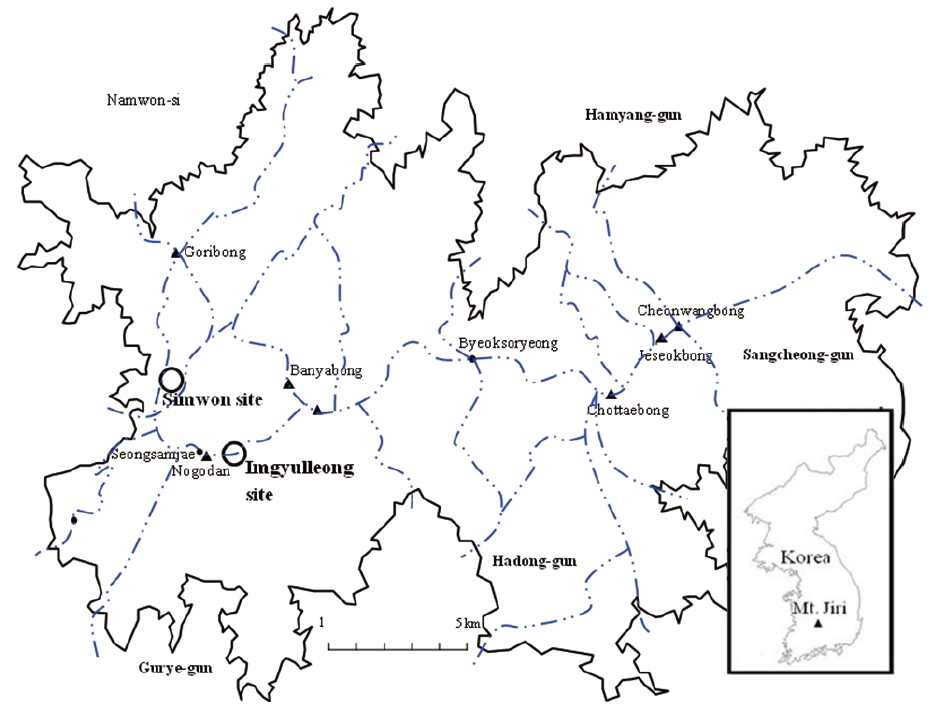
This study was conducted in two Q. mongolica stands on the western part of Mt. Jiri, which is located in the southern part of Korea. Mt. Jiri is the second highest mountain in South Korea with most ridges and peaks reaching altitudes over 1000 m asl. Q. mongolica is found at altitudes above 850 m asl on Mt. Jiri (Yim and Kim 1992, Korea National Park Service 2008). One study site (the Simwon study site) was located at the lower altitudinal limit of the Q. mongolica-dominant area and the other site (Imgyulleong study site) was located within the altitudinal range of Q. mongolica-dominant areas on Mt. Jiri (Fig. 1). The Simwon site was located from ridgetops to lower slopes near a valley on a steep slope (Table 1). The Imgyulleong site was about 0.7 km west of Imgyulleong at 1357.4 m asl. The Imgyulleong plot was located on an upper slope and included a small peak on a gentle slope. While the Simwon plot was in a quite rocky area because it was near a valley, rock exposure in Imgyulleong plot was less than 20%. Both stands had trees with fire scars, indicating that they had experienced forest fire in the past.
Climate data from the Seongsamjae weather station, which was located between two study sites at 1100 m asl, for 3 years from April 2002 to April 2005 showed an annual mean air temperature in the study area of 7.8℃ and annual mean precipitation of 2083.5 mm (Korea Meteorological Administration, 2008). Based on meteorological data from Hobo weather stations (HOBO H08-032-08, Onset Computer Corporation, USA) placed in both study sites, the difference in annual mean temperature between the two plots was 2.1℃
Data were collected in a 1-ha plot (100 m × 100 m) in each study site. The study plots were divided into twenty-five 20 m × 20 m subplots. All trees with ≥ 2.5 cm diameter at breast height (DBH) were measured, and the species name, DBH, and height of individual trees were recorded. All standing trees ≥ 2.5 cm in DBH were mapped and their geographic coordinates were recorded. Height was measured using a hypsometer (Vertex laser, Haglof, Sweden), and DBH was measured at a height of 1.3 m above the ground. We determined whether the stems had originated from seeds or sprouts by examining the stem numbers of a tree. Single-stemmed trees were recorded as being of seed origin and multi-stemmed trees were considered to be of spout origin (Dolezal et al. 2009).
Where n is the number of trees, A is the plot size (m2), and uij is the distance between trees i and j. The weight function, wij provides the edge correction. It(u) is the counter, which has a value of 1 if u ≤ t (the tree is within the circular plot of radius t) or 0 otherwise (Haase 1995, Salas et al. 2006). If a tree is distributed at random, the the expected value of K(t) is equal to πt2. We used the L(t) function, modified from K(t) as follows, to check the difference between the observed value and the expected value (zero) under a random distribution (Besag 1977, Salas et al. 2006).

We used Monte Carlo simulations to test the hypothesis of random distribution of trees. We conducted each simulation 99 times to determine 95% confidence envelopes (Haase 1995). If L(t) was greater than, equal to or less than the confidence envelopes, the we described the spatial pattern of trees as clumped, random or regular at any distance, respectively (Ripley 1977, Leemans 1991, Fajardo and Alaback 2005, Salas et al. 2006). The value of t ranges from 0 to 50 m with 1-m intervals.
We conducted bivariate spatial pattern analysis to detect the degree of repulsion, attraction or indifference between two species (Anderson 1992, Haase 2001, Salas et al. 2006, Song 2007). The bivariate estimator, K12(t) is calculated from K*12(t) and K*21(t), which represents the interaction between species 1 and 2, and species 2 and 1, respectively. The formulas for K*12(t), K*21(t) and K12(t) were as follows:
where, n1 and n2 are the numbers of trees in species 1 and 2, respectively. K12(t) can be modified in the manner of the univariate analysis, as follows:
We conducted each simulation 99 times to determine 95 % confidence envelopes. If L12(t) was inside of the confidence envelope, we considered the association between two species to be independent, whereas if L12(t) was greater than the upper limit of the confidence envelope we concluded that there was attraction between the two species at distance t, and if L12(t) was below the lower limit of the confidence envelope, we concluded that the two species displayed repulsion. The value of t was allowed to range from 0 to 10 m at 1-m intervals. We used the spatial statistics program SPPA 2.0.3 (Haase 2004) for the spatial point pattern analysis.
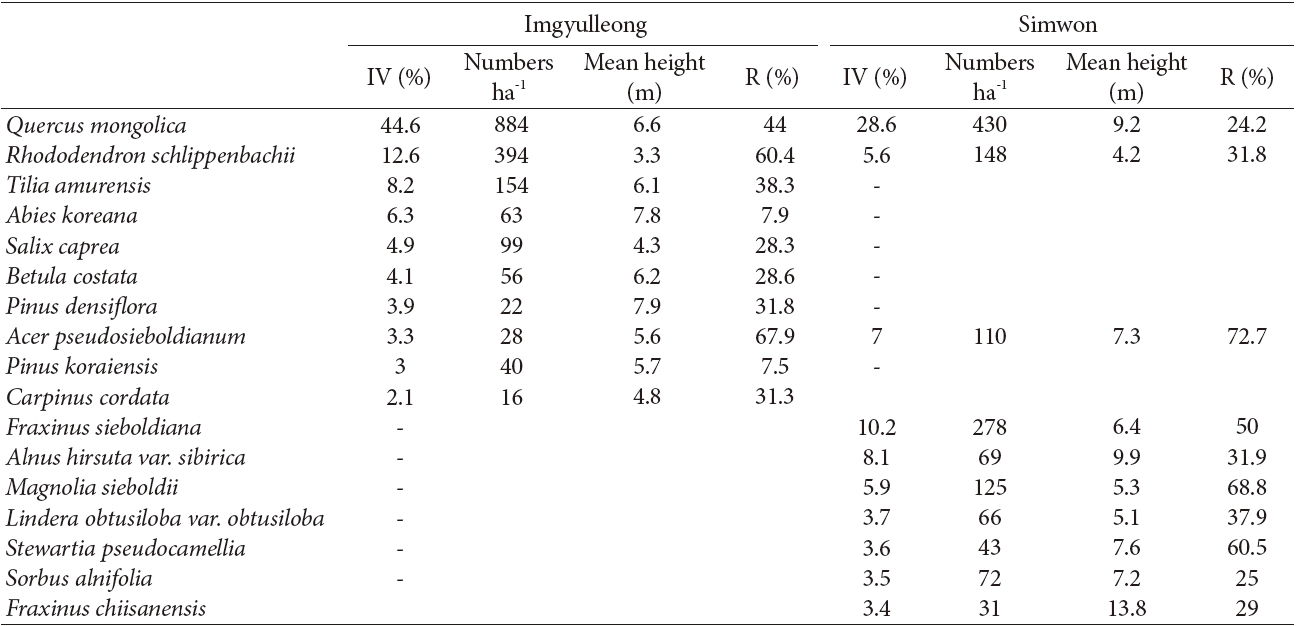
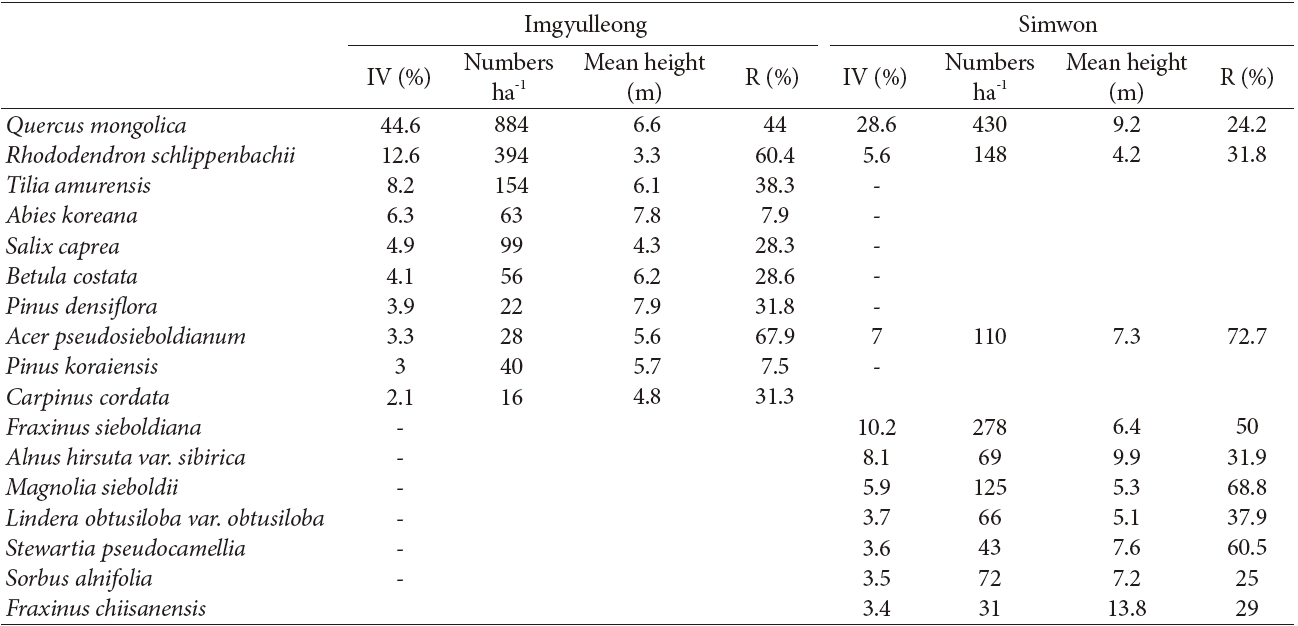
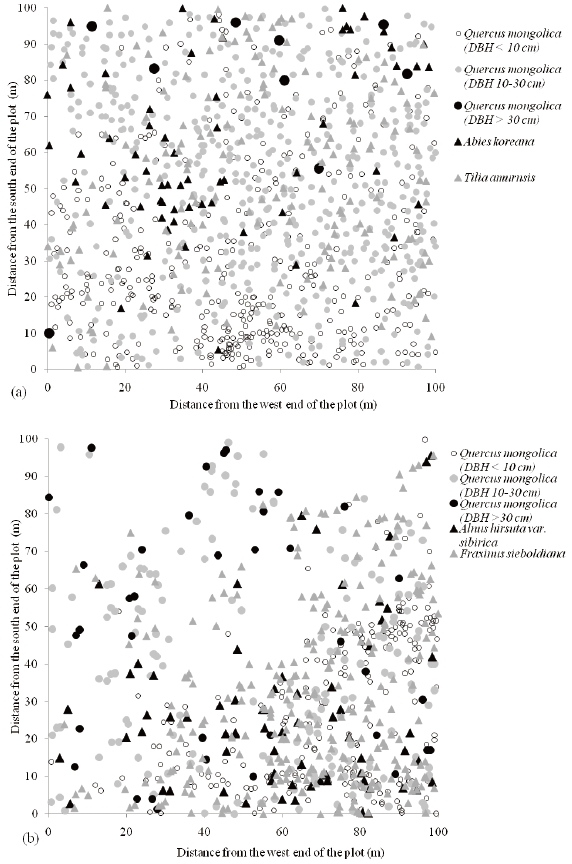
The species composition of the Imgyulleong and Simwon study sites differed (Table 2). At the Imgyulleong site, Rhododendron schlippenbachii Maxim., Tilia amurensis Rupr., and Abies koreana Wilson were the species with the greatest importance values, whereas in the Simwon site, the species with the greatest importance values (in orer of decreasing importance) were Q. mongolica, Fraxinus sieboldiana Blume, Alnus hirsuta var. sibirica C. K. Shneide, Acer pseudosieboldianum (Pax) Kom., and Magnolia sieboldii K. Koch.
The overall density of trees at the Imgyulleong site was 1807 individuals/ha which was slightly higher than the density of 1575 individuals/ha at the Simwon site. The Imgyulleong site had a Q. mongolica density of 884 individuals/ha, which is double the Q. mongolica density at Simwon of 430 individuals/ha. The overall basal area of trees at the Imgyulleong site was 17.4 m2/ha, which was also greater than the basal area of 11.2 m2/ha at the Simwon site. The proportion of Q. mongolica trees of sprout origin was estimated as 44% at Imgyulleong and 24.2% at Simwon. A. pseudosieboldianum, Q. mongolica and S. pseudocamellia had a high proportion of individuals of spout origin. Most shrub species in both study sites seemed to originate from sprouts.
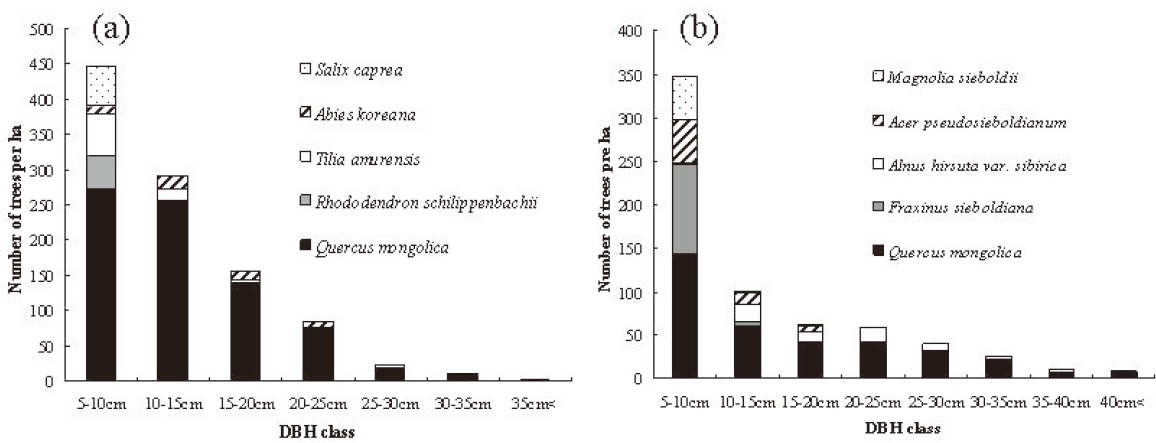
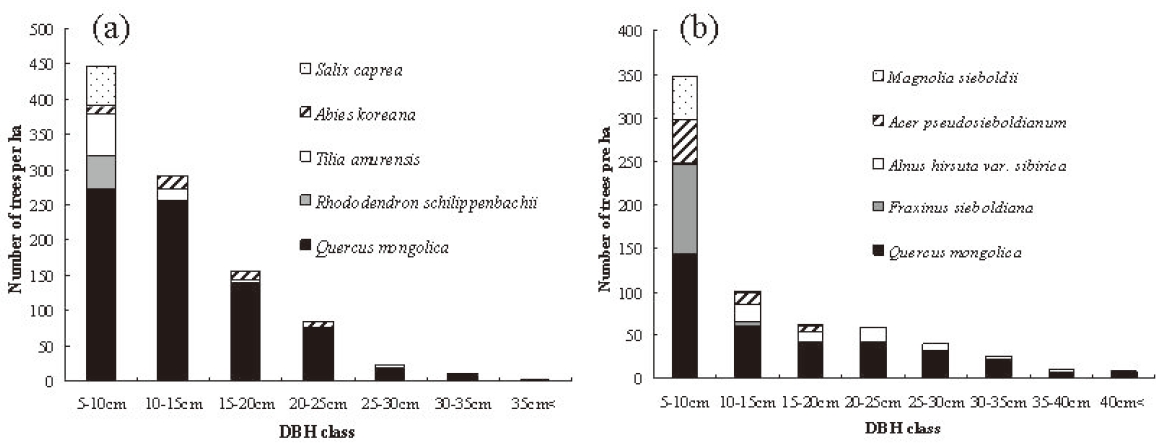
Q. mongolica showed a reverse-J shaped DBH distribution at both study sites (Fig. 2). In Imgyulleong site, A. koreana was evenly distributed in most DBH classes, and T. amurensis and Salix caprea L. were evenly distributed in lower DBH clases. At Simwon, A. hirsute var. sibirica was evenly distributed in most DBH classes, and F. sieboldiana was evenly distributed in DBH classes < 15 cm DBH. The largest tree DBH was 39.9 cm (A. koreana) and the largest Q. mongolica DBH was 36.9 cm at Imgyulleong, indicating that few trees >; 40 cm DBH existed at the Imgyulleong study site. The Simwon site had more large trees than Imgyulleong. At Simwon, the largest tree DBH was 58.0 cm (Q. mongolica), and several trees had DBH >; 40.0 cm, including an individual Fraxinus chiisanensis Nakai with 45.0 cm DBH.
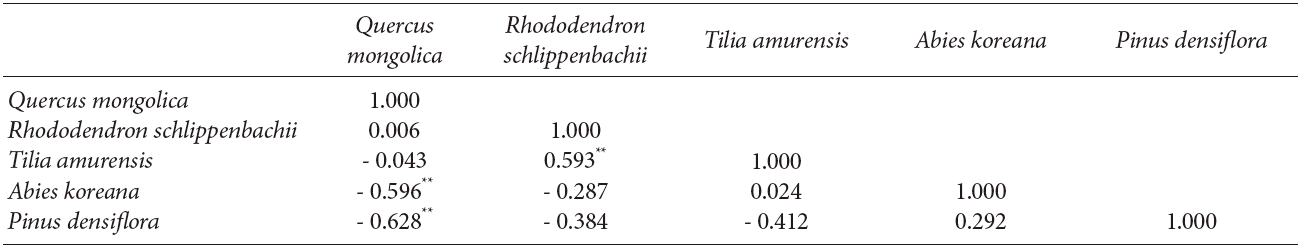
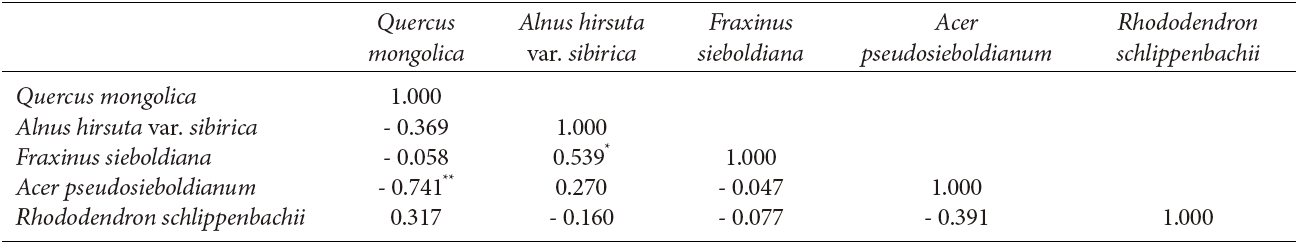
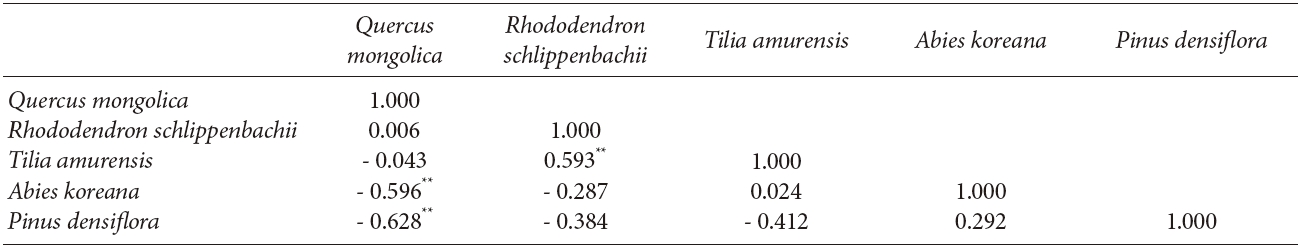
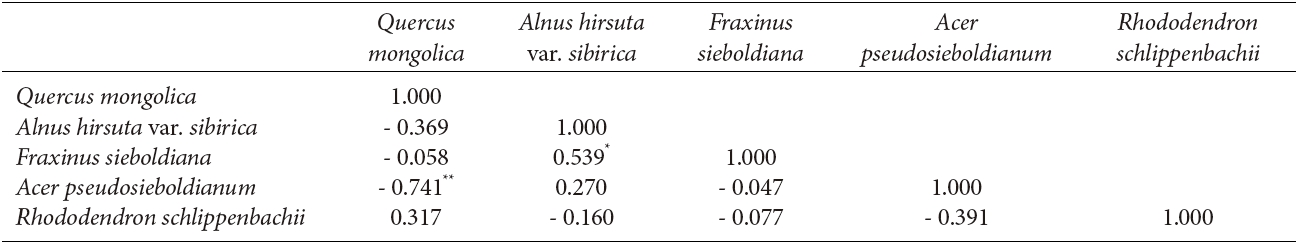
The interspecific spatial association for the five major species at Imgyulleing showed that Q. mongolica had a negative relationship with A. koreana and Pinus densiflora Siebold & Zucc. (P < 0.01), while R. schlippenbachii had a positive relationship with T. amurensis (P < 0.01, Table 3). At Simwon, the spatial associations were not significant for most species except for the negative relationship between Q. mongolica and A. pseudosieboldianum (P < 0.01) and the positive relationship between F. sieboldiana and A. hirsute var. sibirica (P < 0.05, Table 4).
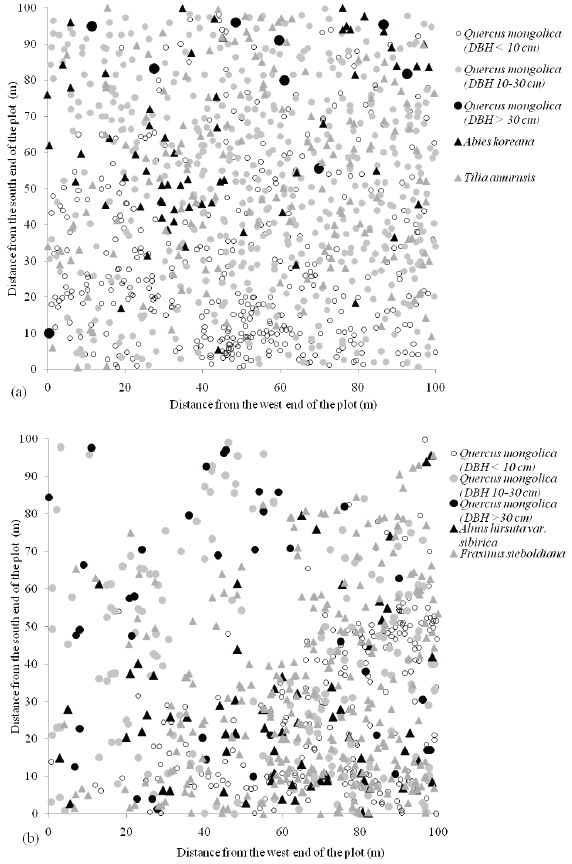
Q. mongolica is distributed throughout the plot at the Imgyulleong site (Fig. 3a), while Q. mongolica is concentrated in the southeast part of the plot in the Simwon site (Fig. 3b). Q. mongolica individuals >; 30 cm DBH were found near the north end of the plot whereas Q. mongolica individuals < 10 cm DBH were more frequent in the south part of the plot in Imgyulleong site. The density of Q. mongolica < 10 cm DBH had a distinctive division in a diagonal direction (SW-NE), which coincided with the ridge direction in the Simwon site. The northwest part of the plot in Simwon was rocky in the area leading down into the valley. Q. mongolica < 10 cm DBH were mostly distributed in the southeast part of the plot, which consisted of the ridge and the upper slope.
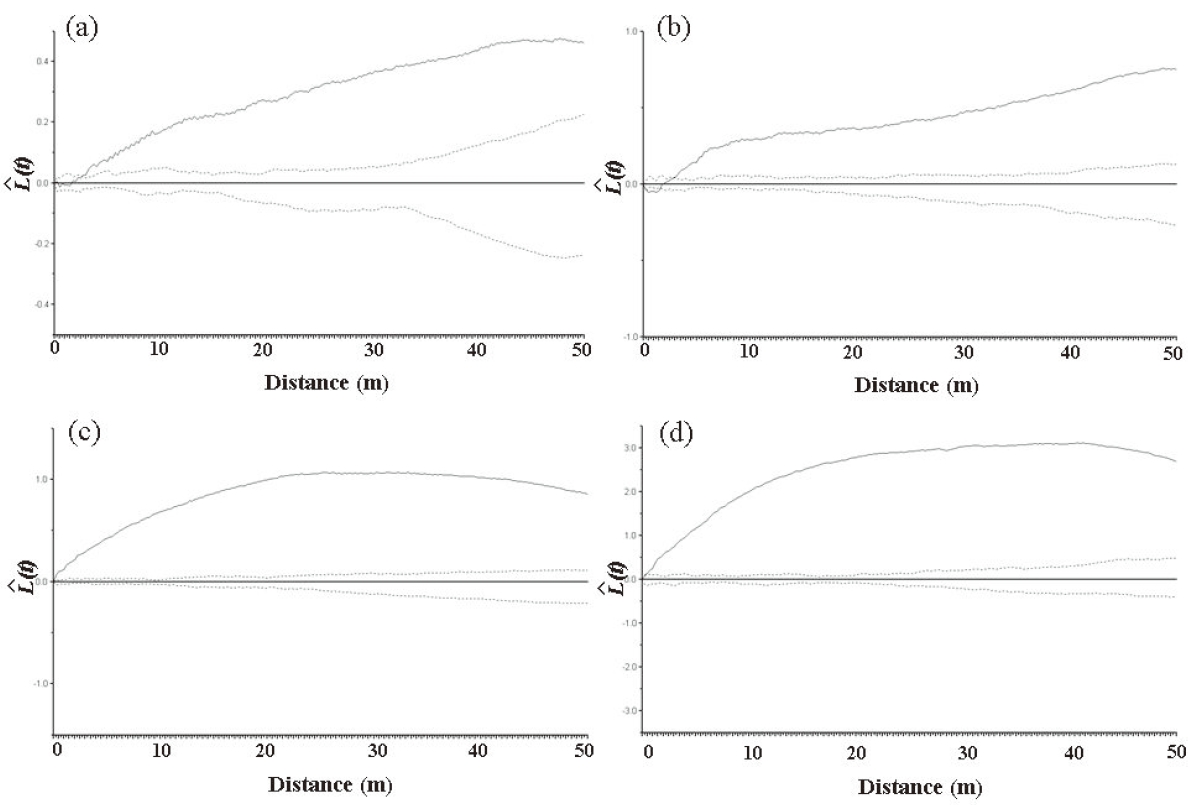
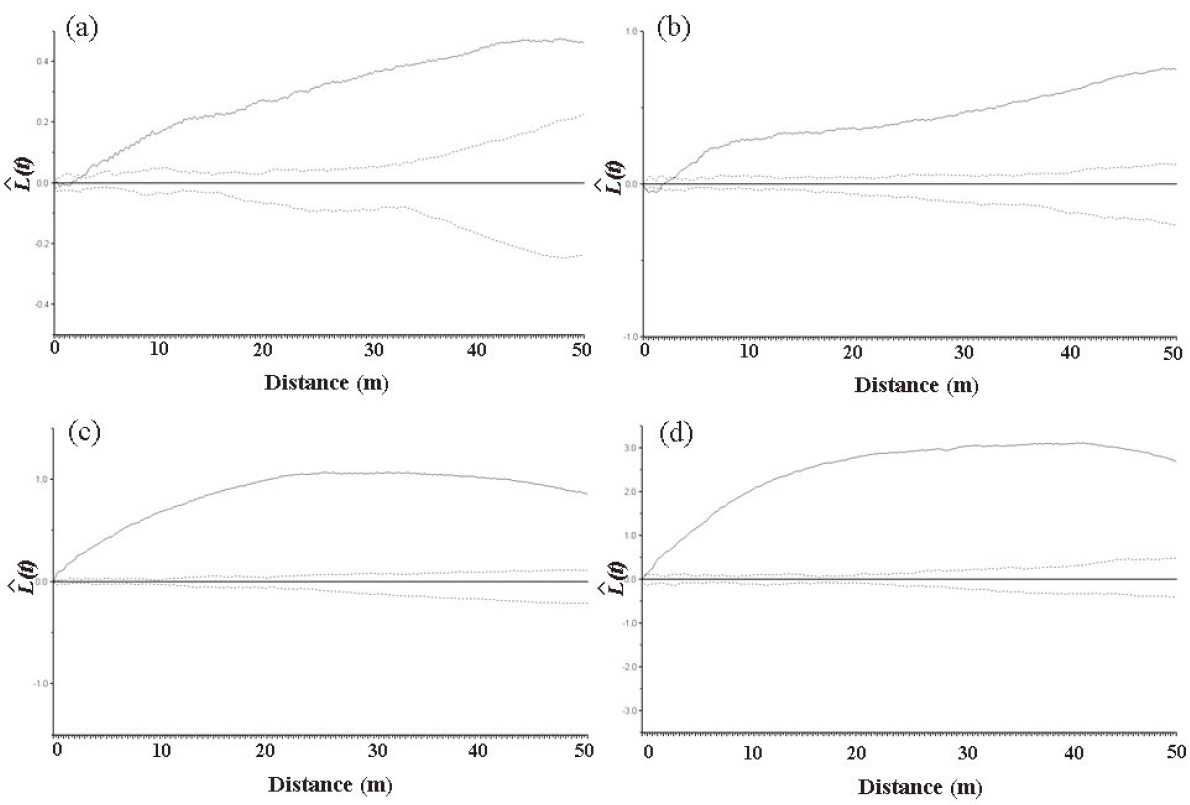
In the Imgyulleong site, all trees showed a clumped distribution for distances > 3 m. For distances < 3 m, most trees showed a random distribution (Fig. 4a). All trees showed a clumped distribution for all distances at the Simwon site (Fig. 4c). Q. mongolica showed a clumped distribution for all distances at both sites (Fig. 4b, d).
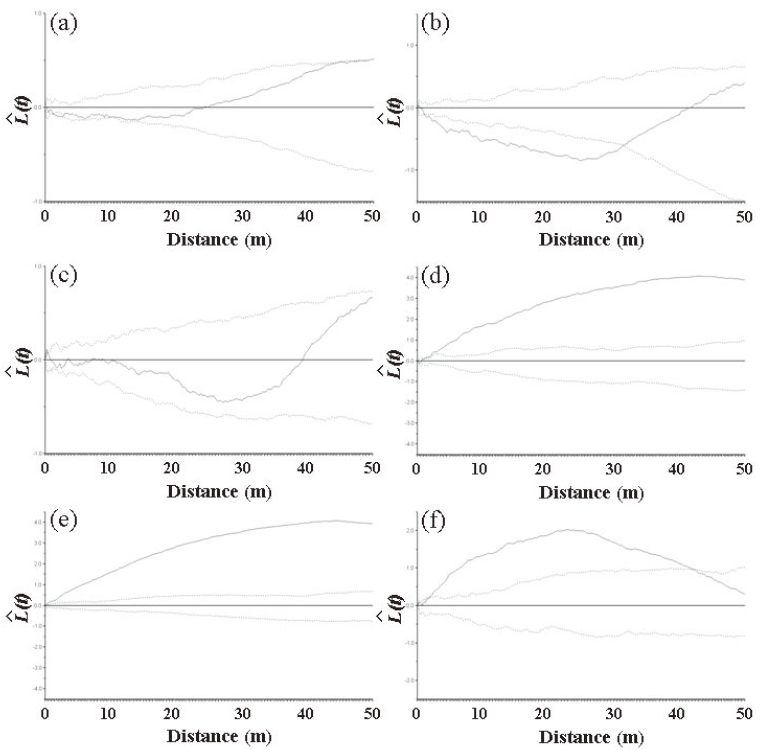
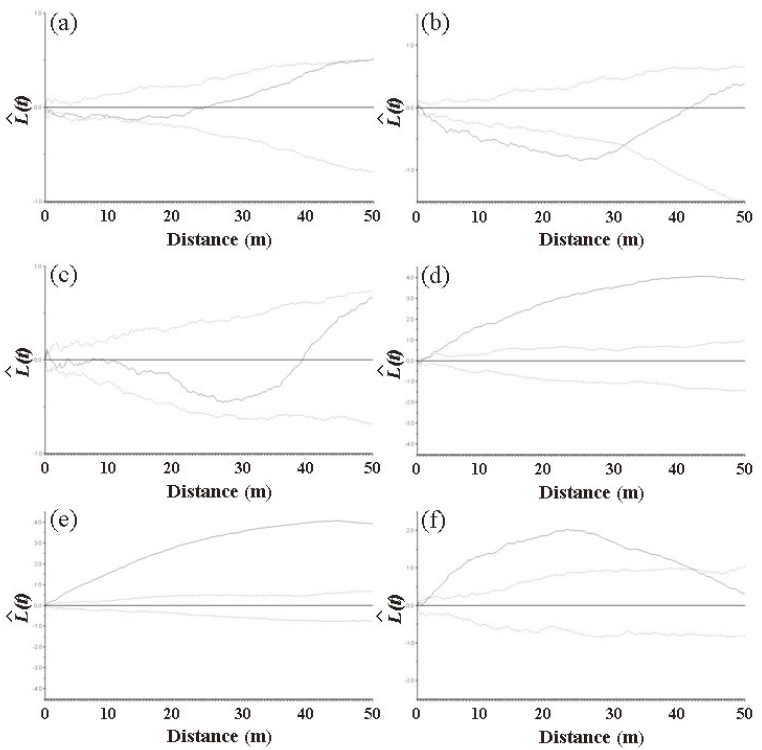
At Imgyulleong and Simwon, the bivariate spatial patterns differed between Q. mongolica and other major species (Fig. 5). Q. mongolica at Imgyulleong had a distribution that was independent of T. amurensis and negatively related with that of A. koreana (Fig. 5a, b), while Q. mongolica at Simwon was positively related with A. hirsuta var. sibirica and F. sieboldiana (Fig. 5d, e). The spatial relationship between Q. mongolica and snag was random at Imgyulleong and positive at Simwon (Fig. 5c, f).
Species composition and stand structure
The Imgyulleong site could be characterized as a Quercus mongolica-Tilia amurensis community and the Simwon site was occupied by a Quercus mongolica- Fraxinus sieboldiana community following the categories established by Song et al. (2003). Previous studies of the environmental characteristics of Q. mongolica communities have shown that Quercus mongolica-Tilia amurensis communities have rich soil nutrients, low available soil P, and gentle slopes and that Quercus mongolica-Fraxinus sieboldiana communities have high available soil P, low soil nutrients and steep slopes (Song et al. 2003). The Imgyulleong site had a gentler slope and a deeper soil A layer than the Simwon site, corresponding to the typical characteristics of Quercus mongolica-Tilia amurensis communities, and the Simwon site had characteristics similar to typical Quercus mongolica-Fraxinus sieboldiana communities.
Both study sites had higher stem density, lower basal area and evidence of past fires, indicating that they have become established after recent fire disturbance relatively recently compared with other stands in Mt. Jiri (Park et al. 2006). The Imgyulleong site had a higher stem density and more Q. mongolica individuals than the Simwon site, while the Simwon site had more large trees. The Imgyulleong site seemed to have been disturbed more recently than the Simwon site. Although Q. monglica showed a reverse-J shaped DBH distribution and the DBH distribution was continuous in both sites, this does not mean that Q. monglica has been continuously regenerating. Rather, the majority of Q. mongolica individuals may belong to a single cohort. Large numbers of Q. mongolica in the lower DBH classes might be suppressed trees, and small number of trees > 30 cm DBH might be the fastest-growing trees or remnant trees that survived past disturbances.
Although Q. mongolica is known to have a relatively loose crown structure, providing better light conditions for shade-tolerant late successional species to become established and survive in the understory (Jang and Im 1985), the high densities of Q. mongolica and relatively low importance values of other species in both study sites suggest that the dominance of Q. mongolica in both study sites is likely to continue for a while. However, the continuous DBH distributions of A. koreana in most DBH classes in the Imgyulleong site, the relatively low shade tolerance of Q. mongolica (Lee and Lee 2001), and the results of previous studies of succession from Q. mongolica to A. koreana (Park and Chung 1990) all suggest that A. koreana has the potential to increase in dominance in this area. In the same way, the importance values of F. siboldiana, F. chiisanensis and Stewartia pseudocamellia Maxim. may increase resulting in increased codominance of those species at the Simwon site under Q. mongolica dominance (Park et al. 2006).
The univariate spatial point pattern analysis showed that trees in both study sites had clumped distributions (Fig. 4). The clumped distribution of trees can be partly explained by stand developmental stages in the study sites. As a stand develops after a disturbance, the spatial distribution of trees changes from being clumped to being random or regular distribution. The stand structure and evidence of fire in both sites indicate that these stands have not yet reached the old growth stage (Kim and Kuen 1991).
The large size of Q. mongolica acorns may contribute to relatively the small dispersion range of this species, which also results in clumped distributions (Venable and Brown 1988). Wildlife such as rodents, which are major acorn-dispersing agents, have limited ranges of movement (Miyaki and Kikuzawa 1988), contributing to the small dispersion ranges of Q. mongolica and resulting in a clumped distribution pattern. Another explanation for the clumped distribution of Q. mongolica could be the irregular distribution of proper habitats for Q. mongolica, especially in the Simwon site. A large part of the Simwon site was steep and rocky, and Sasa borealis (Hack.) Makino covered quite a large portion of the area, limiting the opportunities for establishment of Q. mongolica from seeds (Chung and Chung 2004).
A high proportion of Q. mongolica seemed to originate from sprouts in the study sites. Sprouting often produces short, crooked, and multi-stemmed tree forms, which may be favored over tall, straight, single-stemmed trees in ridges or mountain tops where wind damage is prevalent, such as the study sites. The deep root systems of Q. mongolica that allow them to exploit soil water more effectively also enable them to become distributed in upper slopes or ridges where soil water is often deficient. Sprouting and the deep root systems of Q. mongolica gives this species the ability to dominate communities in ridges and mountaintops. However, sprouting also limits the spatial dispersion of this species, resulting in clumped distributions (Peterson and Squiers 1995). Where dispersion is more limited, interspecific associations with other species may become more important as the species compete for light and soil moisture and nutrients within their limited distribution ranges.
The bivariate spatial pattern analysis for the Imgyulleong site showed that Q. mongolica and T. amurensis are distributed independently while Q. mongolica and A. koreana were negatively correlated, which may indicate repulsion. Although covariation analysis showed slightly negative associations between Q. mongolica and T. amurensis, the correlation was not significant, because most T. amurensis were in the lower DBH classes, so T. amurensis was not an important competitor for Q. mongolica in the upper canopy layer, whereas A. koreana was as competitor with Q. mongolica, as indicated by a significantly negative correlation for their interspecific spatial association (r: -0.596, P < 0.01). The random spatial association between snags and Q. mongolica indicated that trees died out from competition on an individual basis, not as a result of interspecific competition with Q. mongolica.
The bivariate spatial pattern analysis for Q. mongolica and A. hirusta var. sibirica and F. sieboldiana in Simwon showed evidence of attraction, suggesting that those species share similar environmental preferences with Q. mongolica. The results of attraction between Q. mongolica and F. sieboldiana in the Simwon site were different from the previous study reporting that Q. mongolica and F. sieboldiana had different habitat preferences (Song et al. 2003). The attraction among species at Simwon might be due to the irregular site conditions at Simwon, as available tree establishment sites were limited by exposed rock area or dwarf bamboo, and trees were concentrated only in available locations. The interspecific spatial associations between Q. mongolica and A. hirusta var. sibirica and F. sieboldiana in Simwon site were slightly negative (Table 4), but the associations were not significant. We could not exclude the possibility of competition between Q. mongolica and A. hirsuta var. sibirica as they exhibited negative relationships in other study sites (Ahn and Lee 1998). Unlike the Imgyulleong site, the positive spatial association of snags and Q. mongolica at Simwon indicated that the mortality of trees might be due to suppression of Q. mongolica in the upper canopy.
An interaction between the developmental stages of the stands, Q. mongolica acorns and ecological characteristics, and the environmental characteristics of the study sites might result in the current clumped distribution of Q. mongolica in the study sites. Considering that the spatial distribution of Q. mongolica in temperate old growth forests is random (Hao et al. 2007), the spatial distribution of Q. mongolica in the study sites may shift to a random distribution if conditions change with stand development. However, the enhanced competitiveness of trees resulting from sprouting and neighboring species interacting with Q. mongolica in the study sites may result in ongoing Q. mongolica dominance and a clumped distribution of species in the study sites for years into the future.
Q. mongolica stands in the western part of Mt. Jiri showed different species compositions and spatial association depending on site and stand characteristics. On the upper slope of Mt. Jiri near ridge areas, A. koreana and T. amurensis were associated with Q. mongolica, and A. koreana had the potential to compete against Q. mongolica. In the lower elevational limit of Q. mongolicadominant stands, A. hirsuta var. sibirica and F. sieboldiana were codominant species. Trees in the study sites showed a clumped distribution, indicating either that the stands have not reached a mature developmental stage yet or that a clumped distribution was favored by the environmental characteristics in the stands. The continuous reverse J-shaped DBH distribution of Q. mongolica demonstrates that the dominance of Q. mongolica in this area is likely to continue for a while.