Two new genera of red algae, Madagascaria erythrocladioides West et Zuccarello and Pseudoerythrocladia kornmannii West et Kikuchi (Erythropeltidales, Compsopogonophyceae, Rhodophyta), were previously described using molecular analysis and confocal microscopy of isolates in laboratory culture. We examined the ultrastructure of both genera to compare with ultrastructure of other members of the class Compsopogonophyceae. Both genera had Golgi bodies not associated with mitochondria and chloroplasts with a peripheral encircling thylakoid similar to all other members of the class studied thus far. Confocal autofluorescence images showed that Madagascaria has a single round central pyrenoid while Pseudoerythrocladia has no pyrenoid. Our electron microscopic work confirms these initial observations. Tables and keys are presented that assist in interpreting cellular details of genera in the class Compsopogonophyceae.
Detailed molecular and culture studies coupled with ultrastructural observations enable us to discover significant new genera and species of microscopic and macroscopic red algae (e.g., West et al. 2007a, 2007b, Yokoyama et al. 2009). Recently two new genera in the Erythropeltidales (Compsopogonophyceae) were described from algae collected in France, Madagascar and Japan and maintained in unialgal culture before conducting molecular analyses (Zuccarello et al. in press). Very few members of the Compsopogonophyceae have been examined by transmission electron microscopy (Scott and Broadwater 1989, West et al. 2007b, Yang et al. in press). It is readily apparent that each genus studied thus far has two distinctive ultrastructural characters: the association of Golgi bodies with endoplasmic reticulum instead of mitochondria as is typical of almost all red algae investigated with transmission electron microscopy (TEM) (Scott et al. 2008, Yokoyama et al. 2009), and the presence of a peripheral encircling thylakoid in the chloroplast. No other multicellular algae have the combination of these two characters except genera in the Stylonematophyceae. However, the unicellular algae Rhodosorus (Stylonematophyceae; Scott et al. 2008) and Corynoplastis (Rhodellophyceae; Yokoyama et al. 2009) also share these characteristics. The phylogenetic interrelationships of these classes of red algae are becoming more fully understood (Yang et al. in press). Since we enthusiastically believe that all applicable techniques should be utilized to identify new taxa, we have conducted an ultrastructural study on two new algae, Madagascaria erythrocladioides West et Zuccarello and Pseudoerythrocladia kornmannii West et Kikuchi (Zuccarello et al. in press).
Methods for collection, isolation and maintenance of cultures are presented in Zuccarello et al. (in press). In addition, several cultures were grown in 30 psu von Stosch enriched seawater (von Stosch 1964) in Pyrex No. 3250 dishes in front of north-facing windows (37o 27′N, 76o 71W). Filaments from cultures grown in window light at 20oC were prepared for TEM as described in Scott et al. (2008).
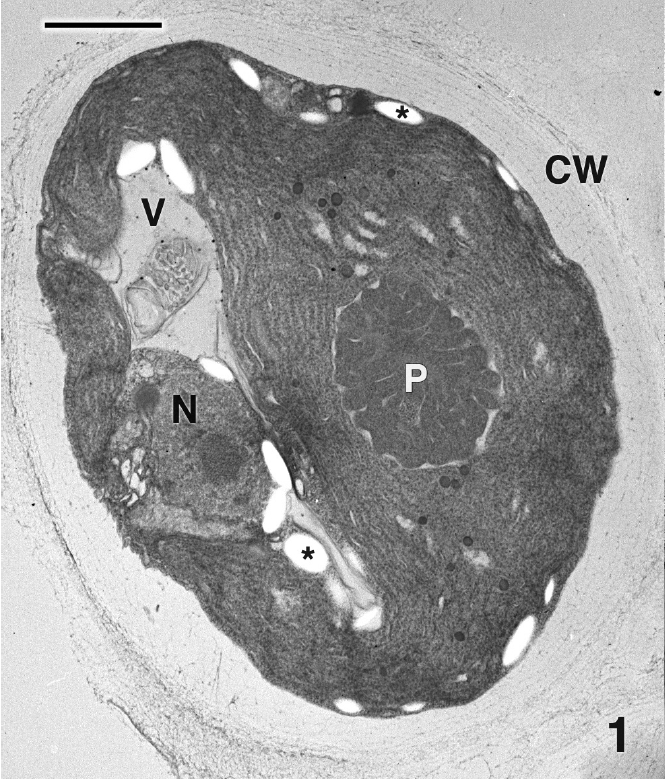
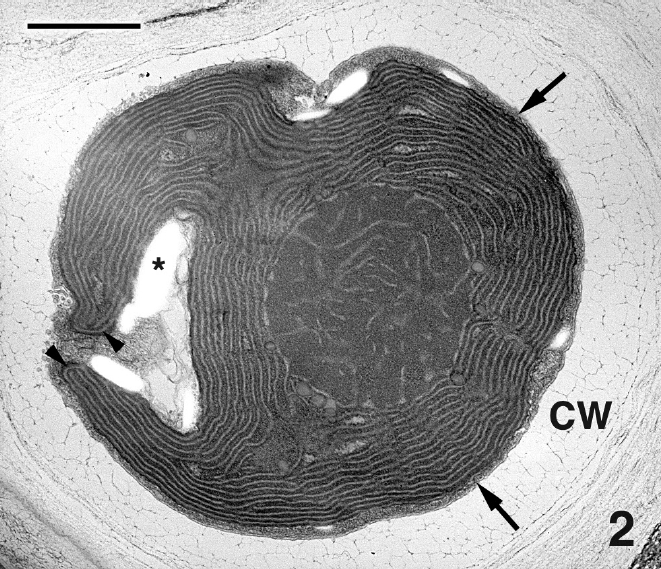
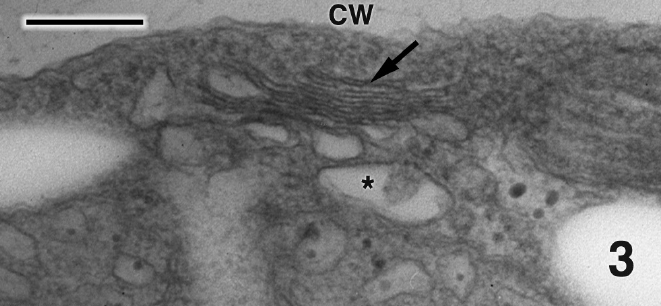
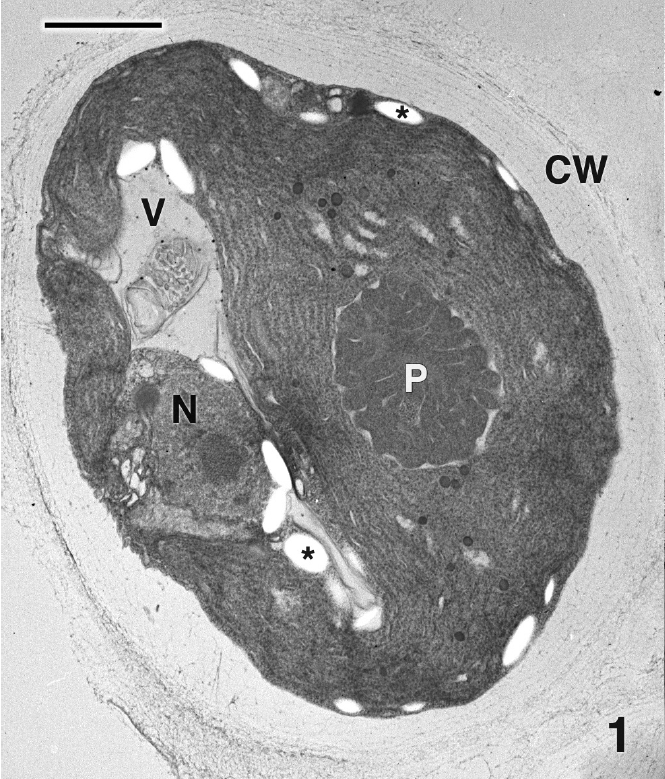
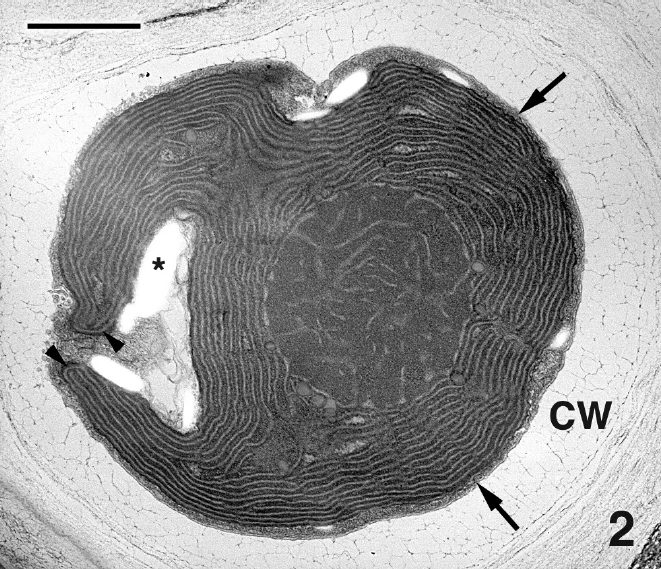
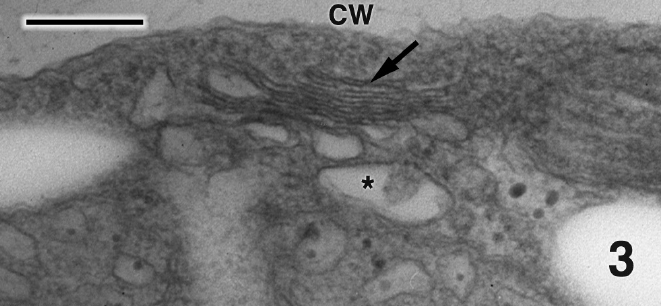
Mature thalli of both Madagascarensis erythroclad-ioides and Pseudoerythrocladia kornmannii are mono-stromatic to distromatic crusts approximately 0.1-2 mm in diameter, depending upon age. In M. erythrocladioides cells are rectangular, approximately 6-12 μm in length and 5-6 μm in width. Figure 1 shows a transverse section through a peripheral crust cell of M. erythrocladioides. A thick fibrillar cell wall surrounds each cell and pit plugs were never seen. The parietal chloroplast occupies most of the cell and contains a large embedded pyrenoid, 1.5-2.0 μm in diameter. Using confocal microscopy Zuccarello et al. (in press) clearly showed large, circular non-fluorescent regions which they interpreted as being pyrenoids in M. erythrocladioides from both Madagascar and Japan. Our TEM images confirm their confocal work. The pyrenoid matrix is moderately electron dense, containing numerous thylakoids derived from thylakoids that delimit the pyrenoid periphery (Figs 1 & 2). Phycobilisomes are present on chloroplast thylakoids but appear absent from those in the pyrenoid matrix. A peripheral encircling thylakoid is present (Fig. 2). The cell contains a small peripheral nucleus, several starch grains lining the chloroplast, a small vacuole containing membranous debris (Fig. 1), and a thin layer of cytoplasm at the cell periphery (Figs 1 & 2). Surprisingly, conspicuous mitochondria were not seen in any of our preparations. Figure 3 shows one of several Golgi bodies detected. Mitochondria were never seen in close association with Golgi bodies, which are usually located at the cell periphery with the cis-region adjacent to the cell membrane.
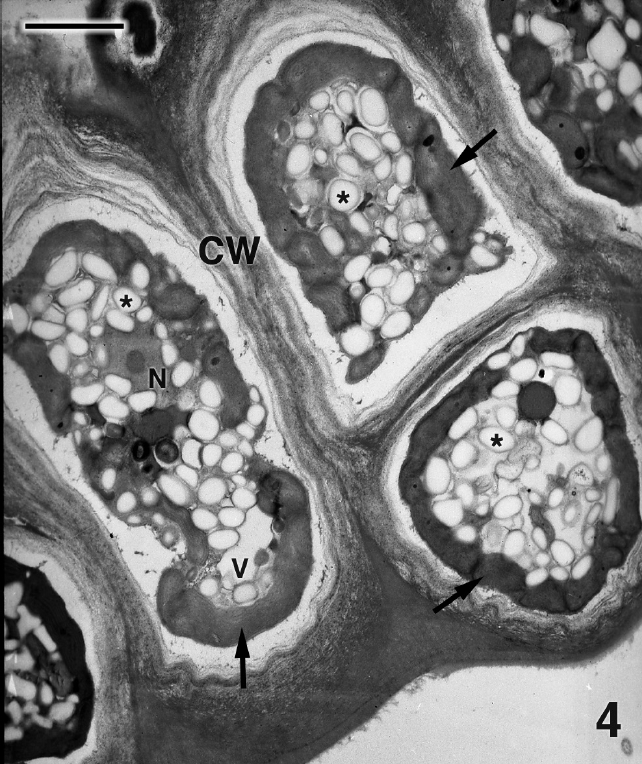
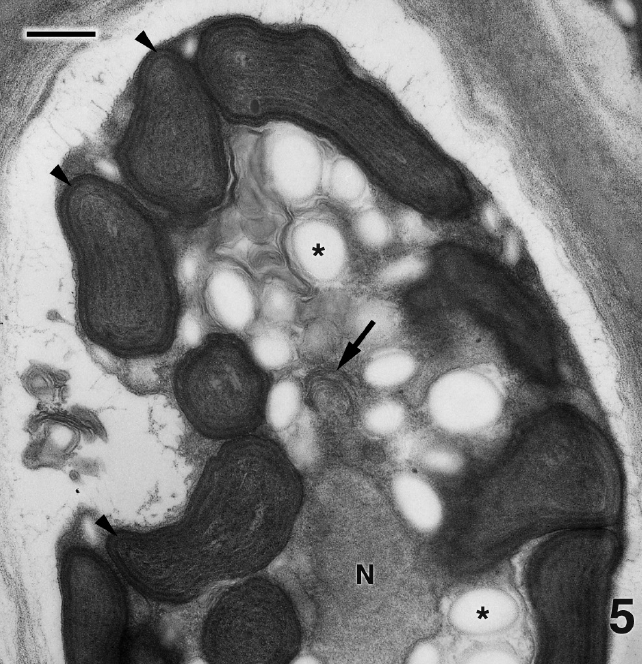
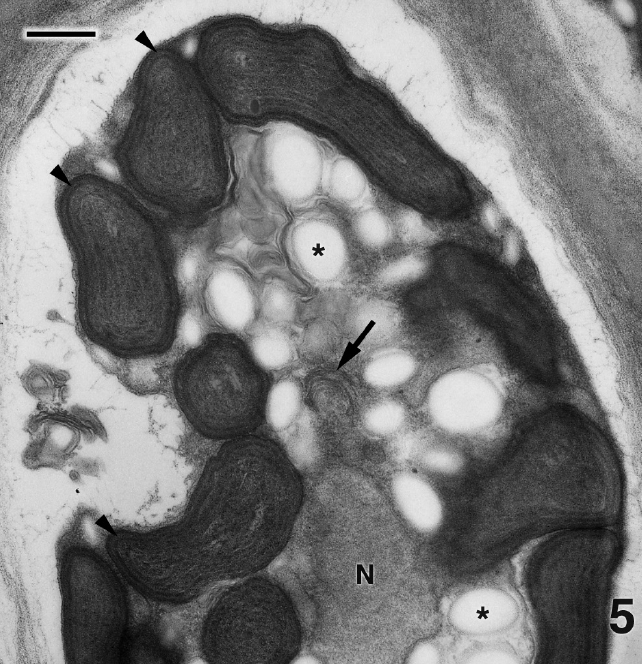
Figure 4 is a glancing section through several cells of P. kornmannii. Cells are rectangular, approximately 6-20 μm in length and 4-8 μm in width. Over 200 cells were examined by TEM. The cell wall consists of several moderately electron dense fibrous layers along with an inner electron transparent layer that most likely represents an artifact of fixation (Figs 4 & 5). Our results confirm the absence of pyrenoids (Figs 4 & 5) reported by Zuccarello et al. (in press) based on confocal microscopic images. One or more parietal chloroplasts are located at the cell periphery and peripheral encircling thylakoids are apparent (Fig. 5). A single nucleus is found in the cell center along with abundant starch, an occasional mitochondrion (not shown), and Golgi bodies not associated with mitochondria (Fig. 5). Very few Golgi bodies were found, even after intensive searches, and more than one Golgi body was never detected in the same cell, although several hundred cells from several preparations were examined. Each cell appeared to have one or more small vacuoles (Fig. 4).
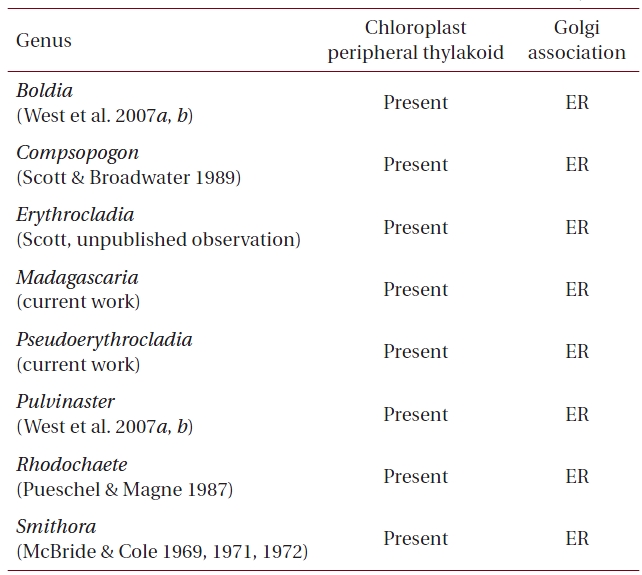
Our results have shown that, as expected, both M. erythrocladioides and P. kornmannii conform to previous TEM work that shows that cells of all members of the Compsopogonophyceae possess Golgi not associated with mitochondrion as well as a peripheral encircling thylakoid in the chloroplasts (Tables 1 & 3 ). What was not well established prior to this work was an awareness of pyrenoid distribution among the genera in this class. Table 2 lists pyrenoid presence and absence in genera of the three orders comprising class Compsopogonophyceae. Pyrenoids are absent in all genera of the Compsopogonales and the Rhodochaetales but present in eight of the ten genera of the Erythropeltidales. Pseudoerythrocladia and Porphyropsis are the only algae lacking pyrenoids.
Ultrastructural studies are remarkably consistent with the most recent red algal supraordinal classification schemes based largely on molecular technology (Saunders and Hommersand 2004, Yoon et al. 2006). Appendix is a dichotomous key based mainly on ultrastructural characters along with algal morphology type (i.e., unicellular vs. multicellular) and life history stage. It is apparent from this table that an unknown macro-stage of a multicellular red alga could readily be assigned to one of three “groups” of classes by examining ultrastructural character states of its Golgi bodies and chloroplasts. 1) If the Golgi is associated with a mitochondrion and a peripheral encircling thylakoid is present, the alga is in class Florideophyceae. 2) If the Golgi is associated with a mitochondrion and a peripheral encircling thylakoid is absent, the alga is in
[Table 1.] Ultrastructural characters of genera in the Compsopogonophyceae
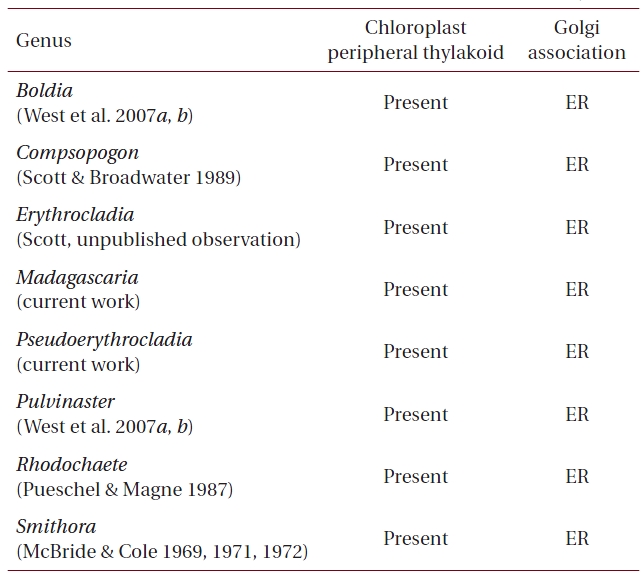
Ultrastructural characters of genera in the Compsopogonophyceae
[Table 2.] Pyrenoid presence or absence in the Compsopogonophyceaea

Pyrenoid presence or absence in the Compsopogonophyceaea
class Bangiophyceae. 3) But if the Golgi is not associated with a mitochondrion and a peripheral encircling thylakoid is present, the alga could be a member of either class Compsopogonophyceae or Stylonematophyceae. We are not suggesting use of this key as a diagnostic guide to solving red algal phylogenetic questions and instead, present it as continued evidence that when molecules speak, cells listen!