The distribution of five cold-water coral taxa (Scleractinia, Zoanthidae, Antipatharia, Octocorallia, Stylasteridae) is influenced by various environmental factors, including depth, slope, temperature, substratum, and current velocity (Freiwald et al., 2004; Roberts et al., 2009). The specific morphological characteristics of Octocorallia in the class Anthozoa and Stylasteridae in the class Hydrozoa that have developed in response to the cold-water conditions are reviewed below.
Two gorgonian families of the class Anthozoa, specifically, Paragorgiidae within the suborder Scleraxonia and Primnoidae within the suborder Calcaxonia, are well known as cold-water coral taxa. These octocorals have a certain degree of flexibility, with their non-rigid internal axis allowing them to bend with water currents. This characteristic is not found in the Scleractinia or Stylasteridae (Roberts et al., 2009).
The paragorgids or “bubblegum corals” of the family Paragorgiidae have dimorphic polyps, and are among the largest corals (up to 10 m in height). These corals represent the most ecologically important sessile benthic deep-water organisms in the ocean (Sánchez, 2005), and are broadly distributed, occurring as deep as 4,152 m (Freiwald et al., 2004; Roberts et al., 2009). While the medulla of these corals is not well consolidated, it contains a dense concentration of calcareous spicules that are bound by strands of gorgonin. Furthermore, the medulla is not separated from the cortex by a ring of boundary canals (Dunn, 1982). There are two genera of paragorgids,
Five species of primnoids from the family Primnoidae have been recorded in Korea (Song, 1981, 2004):
One family of the class Hydrozoa, the Stylasteridae or “hydrocorals” within the order Stylasterina, is the second most species-rich of the 77 hydrozoan families. This family contains 224 valid species, including 24 exclusively fossil species. Some stylasterid species have commercial value in jewelry manufacturing. Consequently, all stylasterids are listed in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) in 1990 (Cairns, 2011a).
The stylasterids or “lace corals” are a significant cold-water taxon. They are fragile, usually small, uniplanar to slightly arborescent colonial hydrozoans of the phylum Cnidaria. This taxon is distributed from the Antarctic to the Arctic circle and occurs at depths ranging from 0 to 2,789 m (Cairns, 2011a). The genus
This study provides a detailed description of two new records of cold-water corals from Korea based on their morphological characteristics,
All specimens examined in this study were collected in fishing nets at depths ranging between 20 and 200 m along the coasts of the East Sea, Korea, from 1976 to 1993. The specimens were anesthetized with menthol for 6-8 h and fixed in 4%-5% (v/v) formalin with seawater. The specimens were then washed with tap water and preserved in 70% alcohol (v/v).
For identification, the internal and external morphological characteristics of each specimen were examined using stereomicroscopes (Semi SV-6 and SV-11; Carl Zeiss, Jena, Germany; and S8APO; Leica, Wetzlar, Germany) and a light microscope (Eclipse 80i, Nikon, Tokyo, Japan). Characteristics included growth form, branching pattern, size of each part of the colony, arrangement of polyps on stems and branches, coloration, and shape and size of sclerites in the polyps and coenenchymes. To examine the sclerites, a small piece of tissue from each body part was dissolved in a diluted solution of Clorox for 5 min.
Before fixation, the specimens were photographed using a digital camera (G7; Canon, Tokyo, Japan). Photographs of the arrangement of the polyps and branches, as well as cyclosystem orientation, were taken using two separate stereomicroscopes (Semi SV-11; Carl Zeiss and S8APO; Leica) mounted with two different digital cameras respectively (5060-WZ; Olympus, Tokyo, Japan and DFC 290; Leica). The sizes and intervals of the polyps and cyclosystems were then measured using an image analyzer (LAS Version 3.6; Leica). Images of sclerites were taken using a light microscope (Eclipse 80i; Nikon) mounted with a camera (DS-5Mc; Nikon). Then, the sizes of sclerites were determined using an image analyzer (NIS-Elements BR 3.0; Nikon). In addition, an image editing program (HeliconFocus 5.1 Pro; Helicon Soft, Kharkov, Ukraine) was used to create a single completely focused image of a sclerite from several partially focused images by combining the focused areas.
Close observation of the cyclosystems and coenosteal texture was achieved using a scanning electron microscope (Model JSM 35CF; JEOL, Tokyo, Japan) at 25 kV accelerating voltage. An ion sputter (Model JFC-1000; JEOL) was used for gold coating.
For identification and classification, the generic revision of the Stylasteridae by Cairns (2011a) and the systematic revision of the Paragorgiidae by Sánchez (2005) were used, along with the review of phylogenetic patterns in the phylum Cnidaria by Daly et al. (2007). Some of the samples examined in this study were deposited in the Korean Coral Resource Bank (KCRB), while other samples were housed in the Natural History Museum, Ewha Womans University, Seoul, South Korea.
Diagnosis. Colonies with an axis or internal axial-like layer (medulla) composed predominantly of sclerites that are either unfused or fused with calcite. Medulla contains endodermal canals and solenia, without a central cord. Colonies generally cylindrical, composed of two layers separated by a ring of longitudinal boundary canals. Proximal portion of polyps embedded in coenenchyme (cortex) with endodermal canals and solenia.
Diagnosis. Dimorphic octocorals with reproductive siphonozooids and feeding autozooids, without axial skeletal structure, other than medulla formed by unfused sclerites. Medulla surrounded by a cortex containing gastric cavities of autozooids and siphonozooids. Profusely branched colonies.
Diagnosis. Medulla perforated by 3 or more large canals in the terminal branches, with numerous smaller canals occurring in both the medulla and subsurface/outer medulla. Autozooid tentacles with sclerites. Surface sclerites 6 to 8 radiates, always less than 0.1 mm in length, with smooth, globular, grooved, or lobulated ornaments. Axial medulla with unfused spindles up to 0.6 mm in length; other sclerites less than 0.15 mm in length.
>
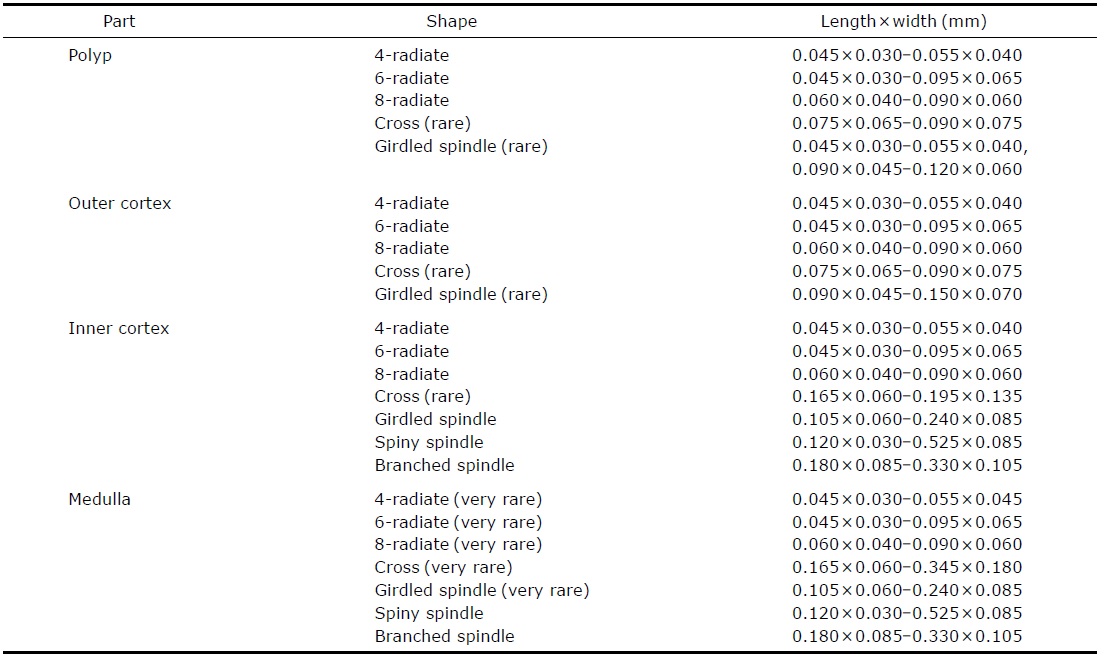
[Table 1.] Measurements (mm) and shapes of sclerites in Paragorgia arborea

Measurements (mm) and shapes of sclerites in Paragorgia arborea
Material examined. Korea: 1 large and 1 small fragments, Gangwon-do: off Samcheok, 37°26′08.53″N, 129°11′25.24″E, 29 Jun 1988, Lee JW, Lee JE (EWZS6312, EWZS6313), 20-30 m deep by fishing nets.
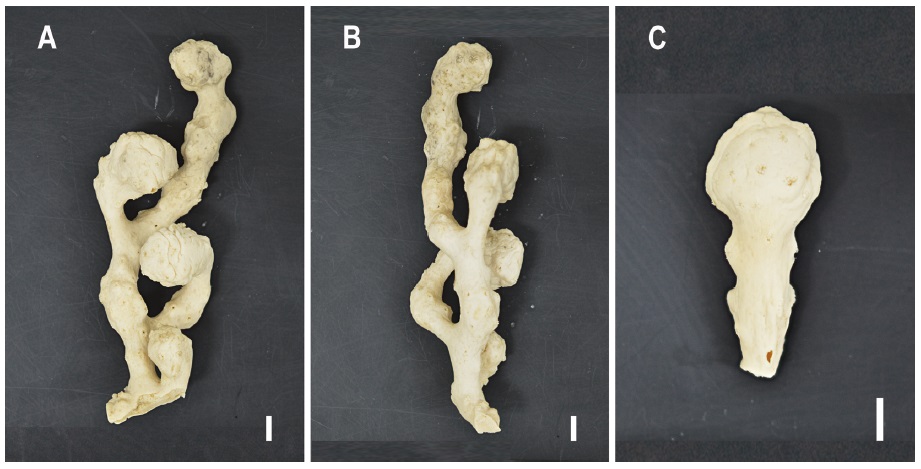
Description. One large fragment (EWZS6312) 205×50×42 mm (height×width×thickness). Colonies sparsely branched at right angles, with each bubble-shaped branch curved toward front side of colony. Branches sympodial, arising 15-55 mm apart. Larger branch tips 22-26 mm in diameter, 25-28 mm in length, and narrow parts 9-11 mm in diameter, 50 mm in length. Smallest branch tip 12 mm in diameter, 18 mm in length, and narrowest part 9 mm in diameter, 6 mm in length.
Medullar region of axis 4.0-5.0 mm in diameter, composed of separable spicules and projections bound together with horny material. Medulla and cortex separated by a ring of canals (boundary solenia). Cortex with 2 distinct layers: outer thinner layer (0.2-0.5 mm in thickness) composed of compact spicules, and inner thicker layer (3.0-7.0 mm in thickness) packed with spicules and solenia.
Polyps dimorphic, autozooids and siphonozooids. Autozooids, 3.0×2.5-4.0×3.0 mm in diameter, completely retracted into coenenchyme, forming rather large hemispherical verrucae with 8-rayed orifices, 4×4-8×9 mm in diameter, clustered in groups on frontal faces of colonies and on tips of distal or lateral branches, and composed of terminal and lateral zooids. Autozooid nodules 15-25 mm in diameter, irregularly situated on narrow parts of branches. Coelenterons on all sides connected with cortex-solenia. Basal part of coelenterons reaching as far as, but not penetrating, medulla.
Inter-nodular surface with numerous and uniformly distributed tiny siphonozooid apertures, appearing as simple pores in rind. Siphonozooids present all over colony, irregularly scattered in vicinity of autozooids, penetrating far into cortex, but not reaching medulla.
In polyps and cortex, shape and size of sclerites similarly developed: 4-, 6-, 8-radiates, crosses, and girdled spindles. In surface of cortex, 6- or 8-radiates with subdivided and grooved ornaments developed. In inner cortex and medulla, large spiny spindles with branching processes developed. Ratio of radiates to spindles 1 : 1 in inner cortex and 1 : 9 in medulla. After decalcification, spindles of medulla separated as other parts, not anastomosed. On polyp bodies, sclerites densely arranged, extending from calyx margin to back side of tentacles.
Colonies orange-yellow when alive, but milky white with pale yellow white medulla when preserved in alcohol. All spicules colorless.
Remarks. The characteristics of the specimens mostly agree with those of Verseveldt (1940) and Sánchez (2005); however, there was variation in the sizes of the sclerites from the different parts of the colonies.
Distribution. Pacific Ocean: Korea (East Sea), Japan (Sagami Bay), New Zealand, Alaska; Atlantic Ocean; Abyssal.
Diagnosis. Gorgonians with an axis of gorgonin containing large amounts of non-scleritic calcite (as internodes or embedded in gorgonin), without a hollow, cross-chambered central core.
Diagnosis. Gorgonians with highly calcified axis and nonretractile polyps heavily armored with calcareous scales.
Diagnosis. Colonies dichotomously branched, usually bushy. Axis densely calcareous, polyps closely spaced and randomly arranged (not in pairs or whorls), usually facing downward. Polyps covered with 6 longitudinal rows of scales, adaxial side largely naked, 8 highly concave marginal scales, adaxials often quite small. Well-developed operculum present, operculars keeled on inner surface. Rind sclerite scales present.
>
Primnoa pacifica
Previous record. Korea: Gyeongsangbuk-do: Ulleung-do (Sadong and Jeodong) (Song, 1981).
Material examined. Korea: Gangwon-do: 28 fragments, Samcheok-si, Jeonglajin, 37°26′08.53″N, 129°11′25.24″E, 29 Jun 1989, Park JH (KCRB1999), by fishing nets (unknown depth).
Remarks. The characteristics of these specimens mostly agree with those of Aurivillius (1931) and Cairns and Bayer (2005); however, the specimens present some differences in the sizes of the scales from the different parts of the colonies. When alive, colonies are assumed to be pink (Cairns and Bayer, 2005) or orange-yellow (Mortensen and Buhl-Mortensen, 2005), but when preserved in alcohol, the colonies are milky white with an axis of glossy metallic yellow. In a previous study (Song, 1981), 1 specimen collected from the deep waters of Ulleung-do Island in the East Sea, Korea, in July 1976, was very large (up to 2 m in height) and retained gonads in the polyp bodies.
Distribution. Pacific Ocean: Korea (East Sea), Japan (Sagami Bay, Misaki, Okino); Aleutian Islands; Okhotsk Sea; Bering Sea; Alaska; California, 9-600 m.
Diagnosis. Gastrozooids short, cylindrical, with filiform tentacles or lacking tentacles; dactylozooids small, finger-shaped, without tentacles; sexual generation by gonophores that remain attached to colony.
Diagnosis. Gastrozooids having filiferan tentacles with desmoneme and eurytele nematocysts.
Diagnosis. Massive calcareous exoskeleton, often brightly pigmented.
Diagnosis. Gastropores and dactylopores arranged in distinct cyclosystems.
Diagnosis. Similar to Group C. In addition to the regularly sympodially arranged cyclosystems at the branch tips, cyclosystems present on the anterior and posterior branch faces, especially on the large-diameter branches. Colonies usually slightly more robust, sometimes bushy.
>
[Table 2.] Number of dactylopores per cyclosystem in Stylaster profundiporus
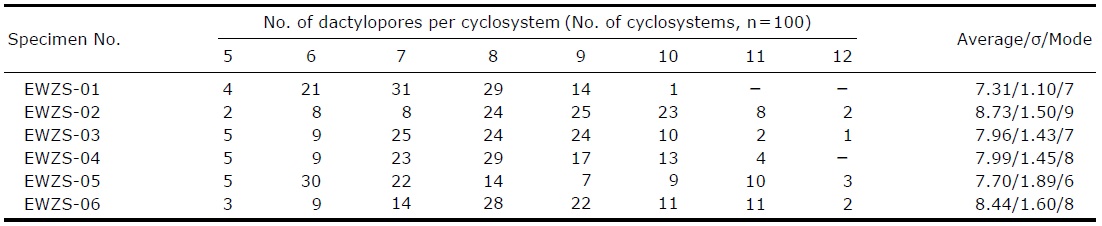
Number of dactylopores per cyclosystem in Stylaster profundiporus
Material examined. Korea: 2 individuals, Gyeongsangbuk-do: Ulleung-do, off Jeodong, 37°29′42.67″N, 130°55′29.13″E, 26 Jul 1976, Song JI (EWZS6314, EWZS6315); 2 fragments, Gyeongsangbuk-do: Ulleung-do, off Dodong, 37°28′44.72″N, 130°54′32.35″E, 27 Jul 1976, Song JI (EWZS6316, EWZS6317); 1 individual and 2 fragments, Gyeongsangbuk-do: off Pohang, 36°03′45.29″N, 129°27′42.03″E, 14 Feb 1990, Ham KH (EWZS6318); 1 individual, Gyeongsangbuk-do: Ulleung-do, Dok-do, off Seo-do, 37°27′27.16″N, 130°53′19.56″E, 22 Mar 1993, Choe BL (EWZS6319), 200 m deep by fishing nets.
Description. Colonies usually flabellate to slightly bushy, irregularly branched, the largest corallum (EWZS6314) 105 mm in height, 85 mm in width, 18 mm in stem diameter. Branches 8-11 mm, twigs 1.2-2 mm in diameter. Branching sympodial, most cyclosystems alternately arranged in series on both sides and on frontal faces of twigs, but not on dorsal faces of corallum. Branch anastomoses rare. Coenosteum polished, porous, reticulate-granular in surface texture.
Cyclosystems circular or oval, slightly protruding above surface, 0.1-0.3 mm in height. Cyclosystems 0.6-1.2 mm, gastropores 0.2-0.5 mm in diameter. Number of dactylopores per cyclosystem (n=100) 5-12, mostly 7-9, mode 8 (Table 2). Dactylopores slit-like, narrow, and arranged in a radial ring like spokes of a wheel, length shorter than diameter of gastropore mouth. Interspaces between slit-like dactylopores shaped like petals of a plume flower. Gastropore tubes cylindrical, straight to curved, about 1.0-1.8 mm deep, each bearing a lanceolate gastrostyle (0.6-0.8 mm in length, 0.2 mm in diameter) covered with sharp spines having a short stalk, 1/2-1/3 of tubes in length. Dactylostyles sometimes rudimentary.
Ampullae superficial hemispheres clustered on anterior and posterior branch faces, 0.5-0.9 mm in external diameter, ampulla cavity below coenosteal surface. Ampullae similar to coenosteum in surface texture, mainly located near cyclosystems at branchlets and twigs.
Colonies mostly salmon pink or pastel peach with twigs pale pink in alcohol or under dry condition.
Remarks. The characteristics of the specimens mostly agree with those of Broch (1936) and Eguchi (1941, 1968), but differ slightly in the number of dactylopores per cyclosystem and in the depth of the gastropore tubes.
Distribution. Pacific Ocean: Korea (East Sea), Japan (Sagami Bay, 82-165 m; off Soto Bosyu, 105 m; off Izu Peninsula, 80-145 m; Kurose, 198 m; south of Shikoku Island, 212 m).
Korean name:1*측산호과 (신칭) Korean name:2*측산호속 (신칭) Korean name:3*나무측산호 (신칭) Korean name:1*히드라충아강 (신칭) Korean name:2*꽃민컵히드라충목 (신칭) Korean name:3*실꽃히드라충아목 (신칭) Korean name:4*의산호과 (신칭) Korean name:5*의산호아과 (신칭) Korean name:6*의산호속 (신칭) Korean name:7*구멍의산호 (신칭)