칼슘이온은 다양한 생리학적 시스템의 중요한 구성요소이며, 생명체의 기능과 생존을 위해 매우 중요한 역할을 담당한다(Herberger and Loretz, 2013). 경골 어류의 주요 칼슘흡수 기관은 아가미이며, 아가미에 존재하는 염류세포가 세포 내 칼슘수송을 조절하는 역할을 하는 것으로 알려져 있다(Payan et al., 1981; Perry and Flik, 1988; Perry et al., 1992). 어류는 성장과 번식을 위하여 지속적으로 수중에 존재하는 칼슘을 섭취하며, 담수어의 경우 사육수 또는 먹이와 같은 주변 매개체로부터 칼슘을 획득하여 혈중 칼슘량을 약 2-4 mmol로 유지한다(Flik and Verbost, 1993).
일반적으로 수중에는 충분한 양의 칼슘이 녹아있으며 어류는 살아가는 동안 필요한 양의 칼슘을 수중으로부터 체내로 흡수하는 것으로 알려졌으나 아메리칸 시클리드(Cichlasoma urophthalmus) 및 나일 틸라피아(Oreochromis niloticus) 등의 어종에서는 추가적인 칼슘 공급이 필요한 것으로 나타났다(Chavez-Sanchez et al., 2000; Shiau and Tseng, 2007). 또한 체내로 흡수되는 칼슘량이 한정되어 있기 때문에 성장기와 성숙기 같이 칼슘 수요가 급격히 증가하는 시기의 어류는 칼슘을 보충하기 위하여 체내의 골조직과 비늘에서 칼슘을 재흡수 하여 이용 하는 것으로 알려져 있다(Flik and Verbost, 1993).
경골 어류는 체내 칼슘량의 약 95%가 뼈와 비늘에 포함되어 있는 것으로 알려졌으며(Flik et al., 1986), 담수어의 경우 금붕어(Carassius auratus), 송사리(Fundulus heteroclitus) 및 무지개 송어(Oncorhynchus mykiss)에서 산란기와 같은 특정시기에 비늘에서 칼슘이 재흡수 되는 것으로 보고되고 있다(Carragher and Sumpter, 1991; Mugiya and Watabe, 1977; Persson et al., 1994, 1995). 그러나 이처럼 뼈와 비늘에서 칼슘 동원이 이루어지게 되면 면역력이 약화 되어 질병에 걸릴 확률이 높아진다. 이를 보완하고자 사료에 칼슘을 첨가하는 연구가 다양하게 진행되고 있으나 사료를 통한 칼슘흡수는 장에서 이루어지는 것으로 알려져 있는 반면, 담수어는 장으로부터의 칼슘 흡수가 거의 이루어지지 않으며 주로 아가미를 통해 칼슘의 흡수가 이루어지는 것으로 보고되고 있다(Furuichi et al., 1997; Hossain and Furuichi, 1999; 2000, Nakamura and Yamada, 1980). 그러므로 담수어의 경우, 사료보다 사육수 내의 칼슘 농도 변화가 체내 칼슘 흡수에 유용한 영향을 미칠 것으로 여겨지나, 사육수 내에 직접적으로 칼슘 농도를 변화시켜 실시한 연구는 미미한 실정이다.
따라서 본 연구에서는 사육수 내의 칼슘 농도에 따른 변화를 관찰 하기 위하여 담수어인 시클리드(Maylandia lombardoi)를 이용하여 실험을 실시하였다. 시클리드(M. lombardoi)는 최근 국내외에서 열대 담수 관상어로서 인기가 많은 어종으로, 추가적인 칼슘 공급이 필요한 것으로 보고 된 나일 틸라피아(O. niloticus)와 같은 척색동물문(chordata) 조기아강(actinopterygian)에 속하는 어종이다. 이 종은 최대 15 cm까지 자라며 마우스 브리딩 하는 난생어종으로 암수가 색으로 구분되는 특징이 있으며, 구입이 쉽고 사육이 간편한 장점이 있기 때문에 본 연구에서는 시클리드(M. lombardoi)를 이용하여 사육수에 칼슘 농도를 인위적으로 조절하였을 때 나타나는 어류의 스트레스 및 면역학적 생리 변화를 관찰하였다.
칼슘 농도가 다른 환경수에서 사육한 실험동물의 생리적 변화를 관찰하기 위하여 인공적으로 사육수를 제조하여 실험에 사용하였다. 사육수 제작은 Chen et al. (2003)의 연구를 참고하여 1차 증류수에 MgSO4, NaCl, K2HPO4, KH2PO4 및 CaO를 각각 첨가하여 제조하였다. Ca2+이 첨가된 실험군은 총 세 그룹으로 22 L의 사육수 안에 CaO를 2 mM, 4 mM 및 8 mM의 농도로 첨가하여 실험에 사용하였다. 모든 그룹의 Mg2+, K+ 및 Na+은 동일한 양으로 각각 첨가하였으며(Mg2+, 0.1 mM; Na+, 0.5 mM; Ka+, 0.02 mM), control군은 Ca2+을 첨가하지 않았다.
본 연구에 사용된 실험어는 대만에서 종묘 생산된 4-6개월령 시클리드(M. lombardoi)치어를 사용하였으며 실험실로 수송 후 3일 간 순응시간을 가진 뒤 실험에 사용하였다. 이후 실험을 위하여 control군과 칼슘 첨가 농도에 따른 2 mM군, 4 mM군 및 8 mM군으로 나누어 28×28×28 cm3의 수조에 한 군당 20마리씩 이송하였다. 칼슘 처리 후 사육 실험은 14일간 진행되었으며 사육 기간 동안 에어레이션과 여과를 지속적으로 시켜주었고, 실험 기간 동안 사육수의 수온은 26-28℃, 광주기는 12L:12D로 유지하였다. 사료 급이 시 사용된 사료는 시중에 일반적으로 판매되고 있는 열대어 전용 사료(tetra bits, tetra)를 사용하였으며 1일 2회 급이 하였다. 실험어의 평균 체장은 2.94±0.06 cm, 평균 체폭 0.88±0.1 cm 및 평균 무게는 0.57±0.04 g이었다.
어체 내 총 칼슘 함량 측정을 위하여 실험종료 후 각 그룹당 6 마리의 실험어를 무작위로 선별하여 실험어의 두부 신경을 빠르게 절단한 뒤, −80℃에 동결하였다. 이후 동결 건조기(PVTFD 10A)를 이용하여 건조 후 분말 형태로 분쇄하였다. 이후 분쇄된 샘플 각각 0.2 g을 질산 10 mL와 과산화수소(electronic grade) 1 mL를 사용하여 PTFE digestion vessel에 옮긴 뒤, 100–180℃에서 가수분해 시켰다. Blank solutions으로는 10 mL 질산과 1 mL 과산화수소를 혼합하여 사용하였다.
칼슘분석은 Agilent 7700s ICP-MS (Agilent, USA)를 사용하여 측정하였다. 무선주파수는 1500 W로 설정하였으며, 아르곤 가스의 속도는 0.5 L min1−1로 조정하였으며, 데이터 수집은 Agilent mass hunter software for ICP-MS (Version A.01.02)를 사용하여 실시하였다.
사육 실험 종료 후, 각 그룹당 14마리의 실험어를 2-Phenoxy-Ethanol 200 μL/L를 이용하여 3분 동안 마취 후, heparine을 처리한 1 mL 주사기를 이용하여 미부 정맥에서 혈액을 채혈하였다. 이후 3,000 g에서 3분간 원심분리 후, 상층액을 분리하여 −80℃에서 동결하여 분석 전까지 보관하였다.
혈액 내 칼슘 함량 분석은 자동 분석기용 Ca2+ 측정용 시약(아산제약)을 사용하여, 제조사의 분석 방법에 따라 생화학 분석기(CH 100 PLUS)로 측정하였다.
칼슘이 스트레스 호르몬인 cortisol에 미치는 영향을 확인하기 위하여 혈액 내 cortisol분석을 시행하였다. Cortisol 분석은 cortisol kit (R&D system)에 포함된 시약들을 사용하여 제조사의 분석 방법에 따라 진행되었다. 우선 RD5-43을 이용하여 혈액샘플을 400배 희석 시킨 뒤, conjugation 시약을 50 μL 첨가한 후, antibody를 50 μL 분주하여 2시간 동안 상온에서 shaking (300 rpm)하며 반응 시켰다. 이후 plate를 wash한 후, 발색 시약인 color regent를 200 μL씩 각 샘플에 첨가하여 빛을 차단한 상태에서 30분간 반응 시켰다. 이후 반응을 멈춰주는 stop solution을 50 μL첨가한 뒤 ELISA (enzyme-linked immunosorbent assay) reader를 이용하여 450 nm에서 측정하였으며, Soft max pro 프로그램을 이용하여 분석하였다.
칼슘이 비특이적 면역계인 lysozyme활성에 미치는 영향을 확인하기 위하여 lysozyme 분석을 실시하였다. 각 실험군의 혈청 15 μL를 96 well plate에 분주한 후 150 μL micrococcus lysodeikticus solution (0.1 M phosphate buffer saline, pH 6.8)을 넣은 뒤 25℃에서 5 분간 반응 후 540 nm에서 흡광도를 측정하고, 다시 5분 간 반응 시켰다. 이후 동일한 파장에서 흡광도를 측정하여 lysis 전 후의 흡광도 차를 비교하여 측정하는 방법을 통해 lysozyme activity 변화를 분석하였다.
모든 분석 결과는SPSS version 21 (SPSS Inc., USA)을 활용하여 통계적 분석을 실시하였다. One-way ANOVA-test로 통계 분석 하였으며, 데이터 값의 유의차는 Duncan’s multiple test 사후분석을 실시하였다. 모든 통계 결과 값은 P<0.05에서 유의성을 판단하였다.
본 연구에서는 수용성 칼슘인 CaO를 이용하여 사육수 내에 인공적으로 칼슘 농도를 조절하였을 때 시클리드(M. lombardoi) 어체 내에서 나타나는 변화를 생리적으로 분석하였다. 우선 어체 내 칼슘 함량과 혈액 내 칼슘 함량을 각각 측정하여 사육수 내의 CaO가 어체 내에 흡수되는 정도를 파악하였다.
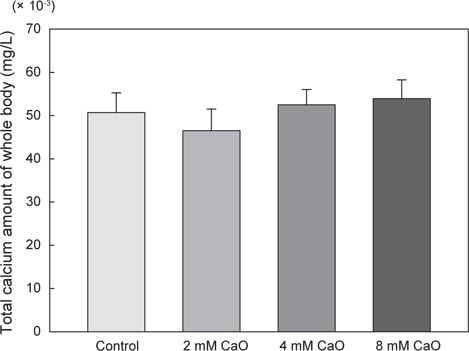
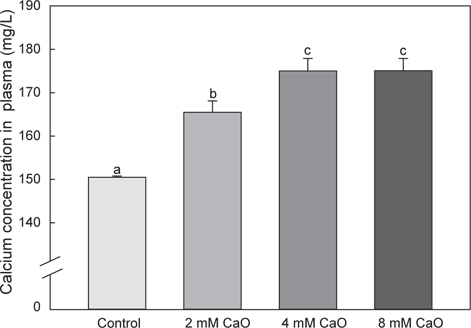
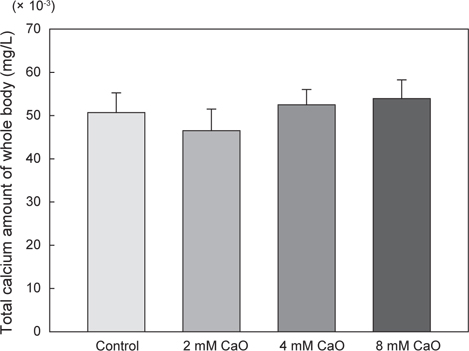
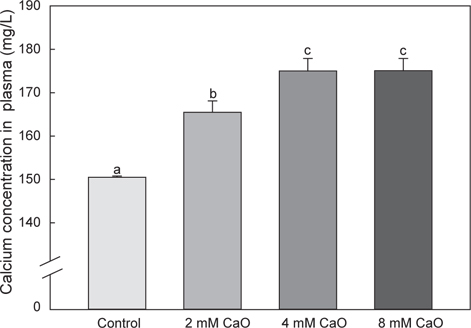
어체 내 총 칼슘 함량의 변화를 측정한 결과, control군에서의 총 칼슘 함량은 50.71±4.5 mg/L이었으며, 2 mM 칼슘 첨가 실험군에서는 46.52±4.98 mg/L, 4 mM 칼슘 첨가 실험군에서는 52.5±3.54 mg/L, 8 mM 칼슘 첨가 실험군에서는 53.92±4.36 mg/L로 8 mM칼슘 첨가 실험군에서 가장 높은 수치를 나타내었으나 각 그룹간에 유의적인 차이는 없었다(Fig. 1, P>0.05). 반면에 혈액 내의 칼슘 함량 변화를 측정한 결과, control군에서 는 150.50±0.29 mg/L이었으나 2 mM 칼슘 첨가 실험군에서 165.50±2.6 mg/L, 4 mM칼슘 첨가 실험군에서 175.0±2.89 mg/L 그리고 8 mM 칼슘 첨가 실험군에서 175.05±2.86 mg/L로 어체 내 총 칼슘 함량과 유사하게 8 mM칼슘 첨가 실험군에서 가장 높은 칼슘 함량을 나타내는 것을 확인할 수 있었다(Fig. 2). 특히 칼슘이 2 mM, 4 mM 및 8 mM의 농도로 증가할수록 혈액 내 칼슘 함량과 환경수 내 칼슘 농도간의 유의적인 차이가 확인 되었으며(P<0.05), 모든 칼슘 첨가 실험군에서 control군에 비해 칼슘량이 농도 의존적으로 증가하는 경향을 나타내었다(Fig. 2). 일반적으로 다양하고 광범위한 수중 환경에서 서식하는 어류의 칼슘 흡수와 조절은 서식지의 환경 요인 등에 의하여 영향을 받게 되는 것으로 알려졌다(Chen et al., 2004). 이러한 수중 환경에 존재하는 칼슘은 주로 탄산칼슘(CaCO3)으로 체내로 흡수되기에 어려운 형태이기 때문에 어류의 혈액 내로 흡수되는 칼슘량이 매우 제한적이다. 따라서 어류는 성숙기 또는 성장기와 같이 칼슘 수요가 증가하는 시기에는 뼈와 비늘에서 칼슘을 재흡수하여 부족한 칼슘을 보충 하는 것으로 보고되었다(Carragher and Sumpter, 1991; Mugiya and Watabe, 1977). 그러나 본 연구에서 수용성 칼슘인 CaO를 이용하여 체내로 흡수된 칼슘의 변화를 관찰한 결과, CaO가 고농도로 갈수록 체내 칼슘량이 증가하는 것을 확인할 수 있었으며, 이것은 이온화된 형태의 칼슘인CaO가 체내에 보다 쉽게 흡수되어 어체 내 칼슘 흡수가 용이하게 이루어져 농도 의존적으로 칼슘 축적량이 증가한 결과로 판단된다. 기존에 진행된 칼슘 수요가 증가하는 시기인 성숙기와 성장기의 해산어 넙치(Paralichthys olivaceus)에 CaO가 첨가된 사료를 공급한 결과, 체내로의 칼슘 흡수가 용이하게 이루어져 비늘에서 칼슘 동원이 이루어지지 않은 것으로 나타났다(Moon et al., 2014). 본 연구에서도 이와 동일하게 담수어인 시클리드(M. lombardoi)를 CaO에 순치한 결과 칼슘 흡수가 용이하게 이루어진 것을 확인할 수 있었다. 따라서 이후 성숙기 어류를 대상으로 하여 CaO가 포함된 사육수에서 순치 할 경우, 칼슘 수요가 급격히 증가하여도 체내로의 칼슘 흡수를 빠르게 유도하여 뼈와 비늘에서의 칼슘 동원이 이루어지지 않고 체내 칼슘량을 일정하게 유지할 것으로 기대된다.
스트레스를 받은 어류의 경우 혈중 cortisol 분비가 증가 되기 때문에 스트레스 평가지표로서 주로 cortisol이 사용되어지고 있다(Barcellos et al., 1999; Nakano et al., 2013; Wendelaar Bonga, 1997). 일반적으로 어류는 수중에 살고 있기 때문에 다양한 스트레스 요인들에 항상 노출되어 있으며, 스트레스는 어류의 생리기능에 변화를 주어 면역기능을 저하시키는 것으로 알려졌다(Espelid et al., 1996; Pickering, 1998). 그러나 스트레스를 주지 않으면서 어류를 취급하는 것은 매우 어렵기 때문에 최근에는 어류의 스트레스 자극 최소화 및 안정화 유도에 관한 연구가 진행되고 있다(Volpato and Barreto, 2001).
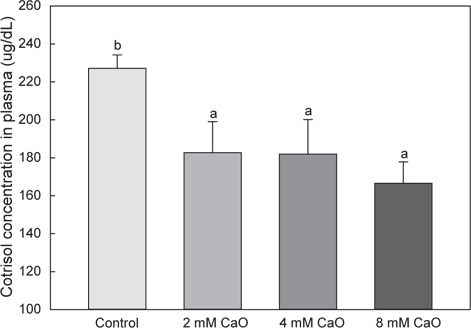
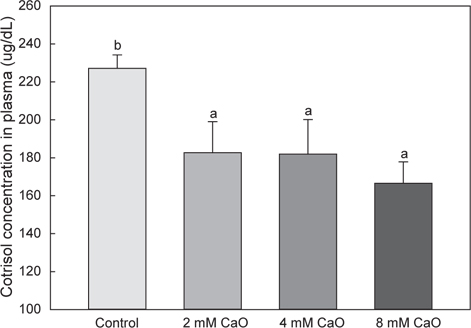
칼슘은 체내의 항상성을 유지시켜주는 물질로서, 스트레스를 받을 경우 체내의 칼슘 항상성에 불균형이 초래되며 이로 인해 항산화 스트레스 및 세포사멸을 가져오는 것으로 알려졌다(Ermak and Davies, 2001). 따라서 본 연구에서는 칼슘이 어체 내 스트레스에 미치는 영향을 파악하기 위해 cortisol 분석을 실시하여 그 변화를 비교하였다. 본 실험 결과 칼슘이 첨가되지 않은 control군의 경우 227.14±7.11 μg/dL, 2 mM칼슘 첨가 실험군에서 182.66±16.41 μg/dL, 4 mM 칼슘 첨가 실험군에서 182.00±18.13 μg/dL 그리고 8 mM 칼슘 첨가 실험군에서 166.54±11.31 μg/dL으로 나타나 모든 칼슘 첨가 실험군에서 control 군에 비해 유의적으로 낮은 수치를 나타내었다(Fig. 3, P<0.05). 어류에서 체내 안정화가 이루어질 경우 혈중 cortisol 수치가 낮아지는 것으로 알려져 있다(Barcellos et al., 1999; Nakano et al., 2013). 따라서 본 연구에서 칼슘이 첨가된 실험군에서 control군에 비해 유의적으로 낮은 cortisol 수치를 나타낸 것은 칼슘이 체내의 안정화를 유도하였기 때문에 어체 내 cortisol의 분비가 감소된 것으로 판단된다.
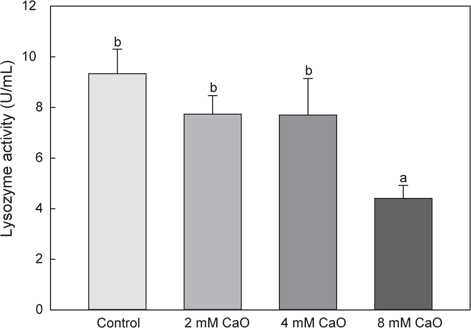
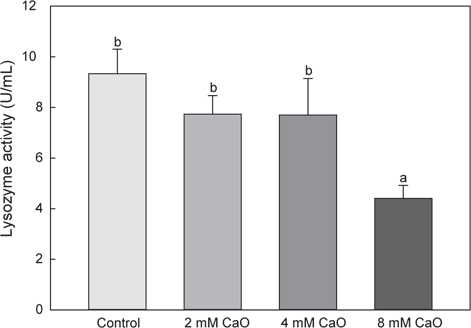
혈액 내의 lysozyme activity의 변화는 control군의 경우 9.33±0.97 U/mL으로 나타났으며, 2 mM칼슘 첨가 실험군에서는 7.73±0.73 U/mL, 4 mM칼슘 첨가 실험군에서는 7.7±1.44 U/mL 그리고 8 mM칼슘 첨가 실험군에서는 4.4±0.52 U/mL로 각각 나타났다(Fig. 4). Lysozyme은 외부 물질에 초기 방어역할을 하고 비특이적 면역활성을 하는 것으로 알려진 비특이적 면역계로, 그 중 어류에 존재하는 lysozyme으로 알려진 calcium-binding lysozyme (c-type lysozyme)은 칼슘과 결합하는 특징을 가지고 있다(Saurabh and Sahoo, 2008). 이전의 보고에 따르면, 무지개송어(O. mykiss)와 나일틸라피아(O. niloticus)의 lysozyme이 스트레스에 의존하는 것으로 보고되고 있으며(Caruso and Lazard, 1999; Mock and Peters, 1990; Yildiz, 2006), 스트레스 관련 호르몬인 cortisol과 마찬가지로 스트레스를 받을 경우 활성이 증가되었다가 시간이 흐른 후 감소하는 것으로 보고되어있다(Ma et al., 2013). 본 연구에서의 lysozyme activity역시 고농도 처리된 8 mM 칼슘 첨가 실험군에서 유의적으로 낮은 활성을 나타낸 것을 확인 할 수 있었으며(P<0.05), cortisol 측정 결과와 동일한 경향을 나타내었다.
이상의 연구 결과를 요약해보면 수중 환경에서의 CaO가 담수어인 시클리드(M. lombardoi) 체내로의 칼슘 흡수를 증가시키고 스트레스 완화 및 면역력을 증강시키는 효과를 가지는 것으로 판단된다. 담수어에 대한 CaO의 효과를 보다 명확히 밝히기 위해서는 이후 CaO의 처리 사육수에서 장기간으로 순치하였을 경우 나타나는 체내의 생리적 변화 및 칼슘 흡수 조절에 관한 연구가 진행되어야 할 것으로 보이며, 특히 8 mM의 CaO를 처리한 칼슘 첨가 실험군의 경우에는 비교적 단기간의 순치임에도 불구하고 체내를 안정화시키는 효과를 나타낸 것으로 판단되기 때문에, 어류 이송과 같은 물리적 스트레스 환경이나 화학적 스트레스에 대한 고농도의 CaO의 효과에 대해 추가적으로 실험을 실시할 계획에 있다.