The pierid butterfly
One common approach to evaluating the effects of tempera ture on insect development is to monitor the duration of the various developmental stages. This permits the determination of two vital parameters of development: the thermal constant (K) and the minimum temperature of development (Tmin). The thermal constant is expressed as the number of degree-days (in ℃) and provides an alternative measure of the physiological time required for the completion of a process or a particular developmental event (Damos and Savopoulou-Soultani, 2008).
Developmental rates decrease with a decrease in environmental temperature and development ceases at very low temperatures. Similarly, developmental rates increase with an increase in temperature up to a certain maximum, and subsequently decrease until reaching a minimum temperature at which development ceases.
The temperature-dependent developmental rate curve of an insect is an important feature of its life history (Taylor, 1981). Using degree-day accumulations to predict a wide variety of events such as egg hatch, adult emergence, or migratory flights may be feasible for insect rearing systems. Degree-day accumulations require a knowledge of both insect developmental responses to temperature and the lower developmental threshold (Woodson and Jackson, 1996).
Butterflies belong to the order Lepidoptera and can be used as indicators of habitat alteration because they are affected by anthropogenic activities, such as urbanization and changes to the natural landscape. Butterflies are also relatively easy to monitor and can therefore be used as an umbrella species when considering local habitat status, environmental conditions, biodiversity, and climate change (Blair, 1999; New, 1997). Butterflies are thought to be sensitive to changes in the environment that can lead to drastic changes in their numbers. These phenomena could justify further study into the changes that occur and the remediation that may be required. Climate change is predicted to lead to an increased mean temperature and more frequent climatic extremes (Pollard
The relationship between temperature and development should be investigated to determine how best to regulate reproduction in the mass rearing of butterflies
Insects. The larval stage of
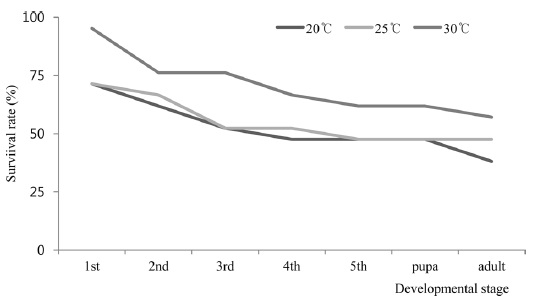
Developmental time and survival. Temperature development studies of
Developmental rate and degree-day requirement. Developmental times at different temperatures were analyzed using the general linear model procedure for each instar separately, as well as for all instars combined. When the effects of temperature were significant (
Linear Model. For larvaal stage, the linear portion (20~30℃) of the developmental rate curve [R(T) = a + bT] was modeled using least squares linear regression, where T is temperature, and a and b are estimates of the intercept and slope, respectively. The base temperature threshold was estimated from the intersection of the regression line at R(T) = 0 and
Statistical Methods. Data were analyzed with analysis of variance (ANOVA) at α = 0.05. Means were separated by the Tukey-Kramer honestly significant difference (HSD) test (Sokal and Rohlf, 1995)
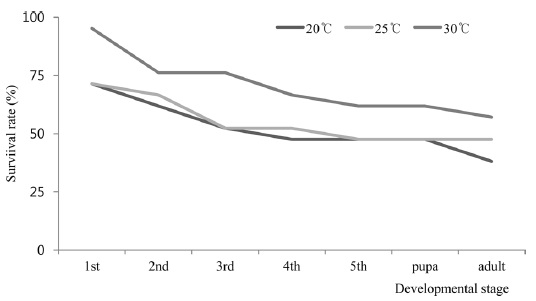
The survival rates of
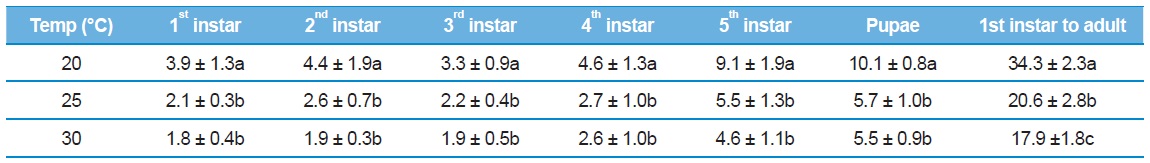
[Table 1.] Developmental duration (mean ± SE) of Eurema hecabe at various constant temperatures
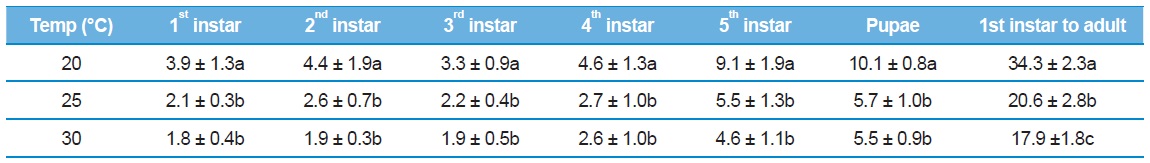
Developmental duration (mean ± SE) of Eurema hecabe at various constant temperatures
The reciprocal of days required for each stage to develop at different temperatures was used to represent the developmental rate (1/d for development). The relationship between the developmetal rate (Y) and the rearing temperature (X) is represented by the regressionformula. The thermal constant is 1/a in Y = aXb, and X is the developmental zero in Y = 0.
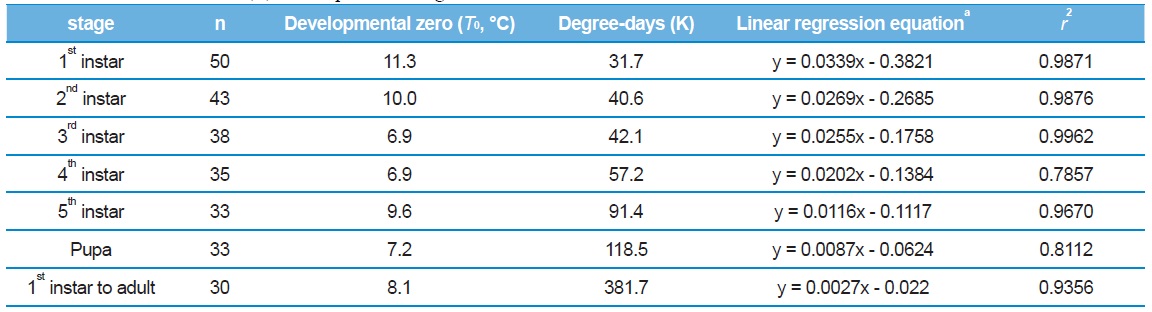
Larval development time decreased between 20 and 30℃, corresponding to a linear increase in development rate (Table 2), with a theoretical threshold value of
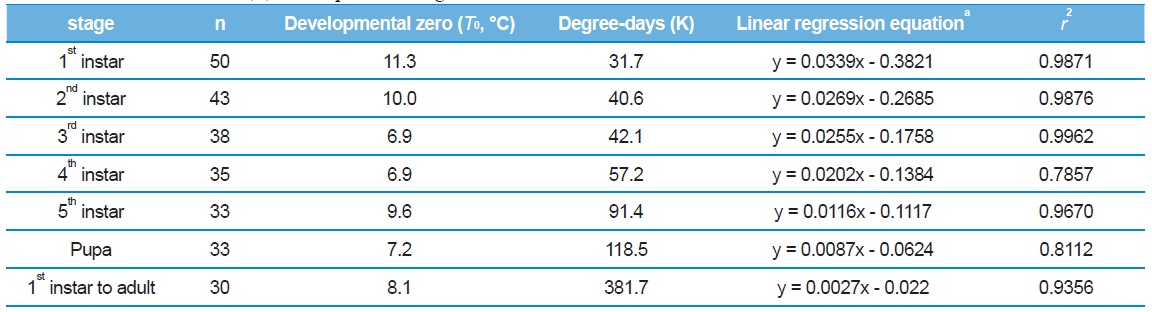
Mean developmental temperature (℃) thresholds (T0), thermal constants (K) in day-degrees above base, regression equations, and coefficients of determination (r2) for the pre-adult stage of Eurema hecabe
According to (Lee
In the mass rearing of any insect, a well-organized work sequence must be planned; for example, adult rearing, egg collection, and rearing from egg to emergence (Koyama
The influence of temperature during development and its effect on developmental stage of
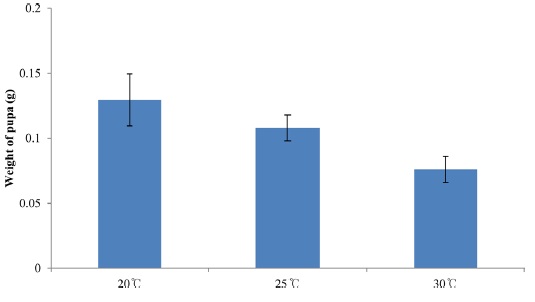
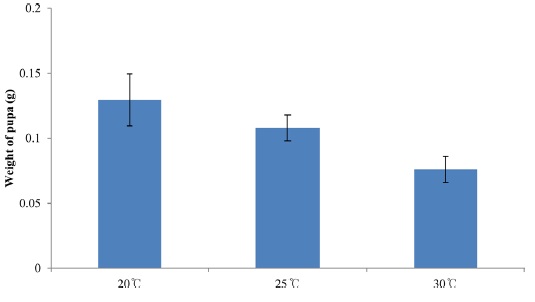
Body size is also plastic and can change in response to different environmental conditions (Stern, 2001). For example, insects that develop at higher temperatures are generally smaller than those that develop at lower temperatures (Atkinson, 1994), and well-fed organisms are typically larger than those fed a poorquality diet (Chapman, 1998). Given a particular growth rate, the final size of an insect should be proportional to its duration of growth. This means that either the developmental duration of an insect may be shortened at the cost of body size, or it may attain a larger size at the cost of prolonged development.
In insects, female lifetime reproductive success (mating ability in particular), is more closely correlated with body size (Honek, 1993) than the major components of male reproductive success are. Thus, in terms of fitness, larger females tend to have more attract.
As an insect larva grows, it normally progresses through a fixed number of instars. During its final instar, the larva attains a critical size that sets in motion the endocrine events that trigger metamorphosis.
Developmental thresholds and degree-day requirements can provide information about life stage events that might aid in developing efficient management strategies.