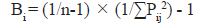어류의 초기생활사 연구는 알과 자어 및 치어 단계를 포함하며, 이 시기를 거쳐 성어로의 가입이 이루어지기 때문에 어업자원관리 차원에서 매우 중요하다. 해마다 변하는 수산자원의 어획량은 이러한 가입량의 변동에 기인한다는 사실은 1900년대 초반에 알려지기 시작했다(Hjort, 1914, 1926). 어류의 난과 자치어는 환경 변화에 민감하며, 특히 부화 후에 난황이 모두 흡수되고 외부섭식이 적절히 이루어지지 못하는 데서 기인한 사망률의 증가는 가입량에 크게 영향을 미치는 것으로 알려져 있다(May, 1974; Lasker, 1975; Hunter, 1976).
어류의 초기생활에서 먹이의 가용성(prey availability)과 섭식의 성공은 가장 중요한 요소라고 할 수 있다. 성어로 가입 전까지 높은 성장률과 생존을 유지하기 위해서는 최적화된 섭식 전략을 고수해야만 할 것이다( Østergaard et al., 2005). 그래서 초기 생활사 단계의 선호하는 먹이생물을 확인하는 것은 섭식 조건과 먹이 요건을 충족하는 자치어의 기회를 평가하는데 중요한 요소이며(Robichaud-LeBlanc et al., 1997), 어류의 초기생활사 모델에 유용한 자료를 제공할 수 있다(Fiksen et al., 1998; Rose et al., 1999).
하구(estuary)는 일반적으로 상류지역으로부터 유입되는 풍부한 영양염의 영향으로 기초 생산력이 매우 높은 장소로 알려지고 있다(Reid et al., 1976). 또한 담수와 해수가 혼합되는 기수역(brackish water zone)을 형성하여 다양한 먹이생물과 서식처를 제공, 기수어종 뿐만 아니라, 많은 해양성 어류가 섭이장, 산란장, 그리고 성육장으로 이용하고 있어 풍부한 어류상을 나타내므로 이에 대한 연구는 어류상을 조사하는 중요성도 있지만 수산자원개발의 측면에서 더욱 중요하다.
꼼치(
지금까지 국내 꼼치에 관한 생태적 연구는 낙동강 하구 꼼치 성어의 식성(Huh, 1997), 광양만 잘피밭에 서식하는 꼼치 유어의 식성(Kwak and Huh, 2003)에 관한 연구가 있었고, 초기 생활사에 대한 연구로는 꼼치와 미거지(Liparis ingens) 난자치어 단계의 형태적 특징(Kim et al., 1986a; Kim et al., 1986b)과 형태적 특징에 기반 한 초기생활사(Jung, 2005)에 관한 연구가 있을 뿐, 자치어기의 식성에 대한 연구는 없었다. 따라서 본 연구는 우리나라 남해안의 대표적인 어류 산란·성육장인 낙동강 하구역에 출현하는 어종의 초기생활사에 대한 종합적인 연구의 일환으로 중요한 상업성 어종 중 하나인 꼼치 자치어의 월별 공간 분포 현황과 주먹이생물을 파악하였고, 자치어의 성장에 따른 먹이변화 양상을 파악하였다.
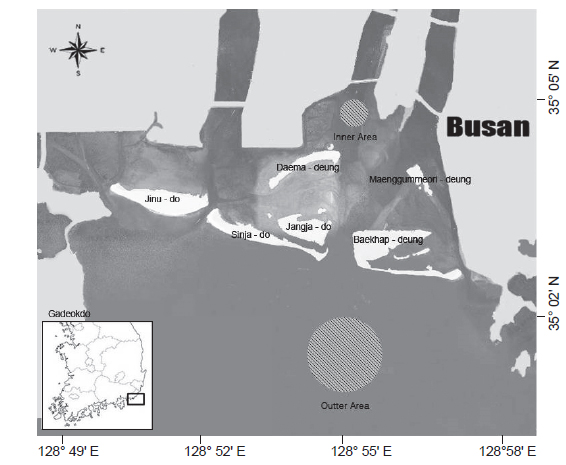
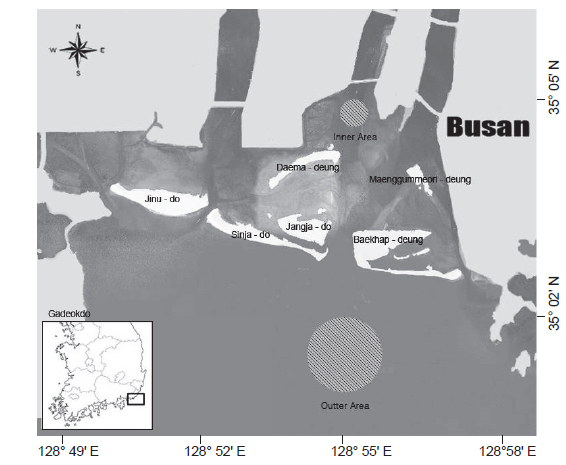
본 연구에 사용된 꼼치 자치어의 시료는 2010년 12월부터 2011년 4월까지 매월 소조기의 낮 동안 채집이 이루어졌으며, 수심이 5 m 내로 얕고, 담수의 영향을 많이 받을 것으로 예상되는 낙동강 하구역 안쪽에 위치한 정점에서 3-4회 반복채집 하였고, 수심이 5-10 m로 비교적 깊고 해수의 영향을 많이 받을 것으로 예상되는 바깥 정점에서 7회 반복 채집되었다(Fig. 1). 시료의 채집은 RN80 네트(망구: 80 cm; 망목: 330 μm)를 이용하여 수심 1-2 m에서 약 2.5 노트(kn)로 10분간 수평 채집을 실시하였고, 정량적 분석을 위하여 네트 입구에 유량계(Hydro-Bios Model 438 110)를 부착하였다.
채집된 표본은 5% 중성 포르말린으로 고정한 뒤, 실험실에서 꼼치 자치어를 분리한 후, 척색장(notochord length: NL)을 0.1 mm까지 측정하여 해부현미경(Olympus SZ40, Olympus, Japan) 하에서 위장을 분리하였다. 장내용물은 쌍안실체현미경(Olympus CH2, Olympus, Japan)을 이용하여 먹이 종류별로 구분하였고, 출현량이 많은 먹이생물은 가능한 속(genus)수준까지 동정하였고, 그 외 먹이생물은 그 보다 상위 분류단계까지만 동정하였다. 먹이생물의 동정에는 Yamaji (1984), Cho (1993), Yoo (1995) 등을 참고하였다.
먹이생물은 종류별로 개체수를 계수한 뒤, 요각류 노플리우스 단계나 기타 소형 먹이생물은 17 μm, 요각류 성체나 기타 대형 먹이생물은 68 μm 단위로 먹이생물의 장축과 단축을 측정하였다. 그리고 먹이생물의 부피를 측정하기 위하여 먹이생물 종류에 관계없이 원기둥으로 가정한 뒤 장축은 높이, 단축은 밑면의 반지름을 구하는데 이용하였고, 원기둥 부피 식인 v=πr2*h 이용해 먹이 생물의 체적을 구하였다.
장내용물의 분석 결과는 각 먹이생물에 대한 출현빈도(frequency of occurrence), 먹이생물의 개체수비와 부피비로 나타내었으며, 출현빈도는 다음과 같이 구하였다.
%F = Ai / N × 100
여기서, Ai는 장내용물 중 해당 먹이생물이 발견된 꼼치 자치어의 개체수이고, N은 위장에 내용물이 있었던 꼼치 자치어의 개체수이다.
섭식된 먹이생물의 상대중요성지수(index of relative importance, IRI)는 Pinkas et al. (1971)의 식을 이용하여 다음과 같이 구하였으나, 먹이생물의 중량비(%W) 대신 부피비(%V)를 이용하였으며, 결과 값은 백분율로 환산하여 상대중요성지수비(%IRI)를 구하였다.
IRI = (%N + %V) × %F
성장에 따른 먹이생물의 변화를 알아보기 위하여 꼼치 자치어의 시료를 2 mm 간격으로 3개의 크기군(3-5 mm; 5-7 mm; >7 mm)으로 나누어 먹이조성을 조사하였고, 척색장과 섭식된 먹이생물 크기(단축) 사이의 관계는 선형회귀분석을 실시하였다. 그리고 성장에 따른 먹이섭식 특성 파악을 위하여 크기군별 개체 당 먹이의 평균 개수(mean number of preys per stomach, mN/GUT)와 부피(mean volume of prey per stomach, mV/GUT)를 구하였으며, 분산분석(analysis of variance, ANOVA)을 이용하여 유의성 검정을 실시하였다.
또한 먹이생물을 얼마나 다양하게 먹고 있는가를 파악하기 위하여 dietary breadth index (Bi)를 구하였다(Krebs, 1989).
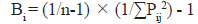
여기서, Pij는 포식자 i의 장내용물 중 먹이생물 j가 차지하는 비율이고, n은 총 먹이생물의 종 수이다. 이 지수는 0에서 1까지의 범위를 보이며, 1에 가까울수록 더 다양한 먹이생물을 섭식하는 종으로 볼 수 있다(Gibson and Ezzi, 1987; Krebs, 1989).
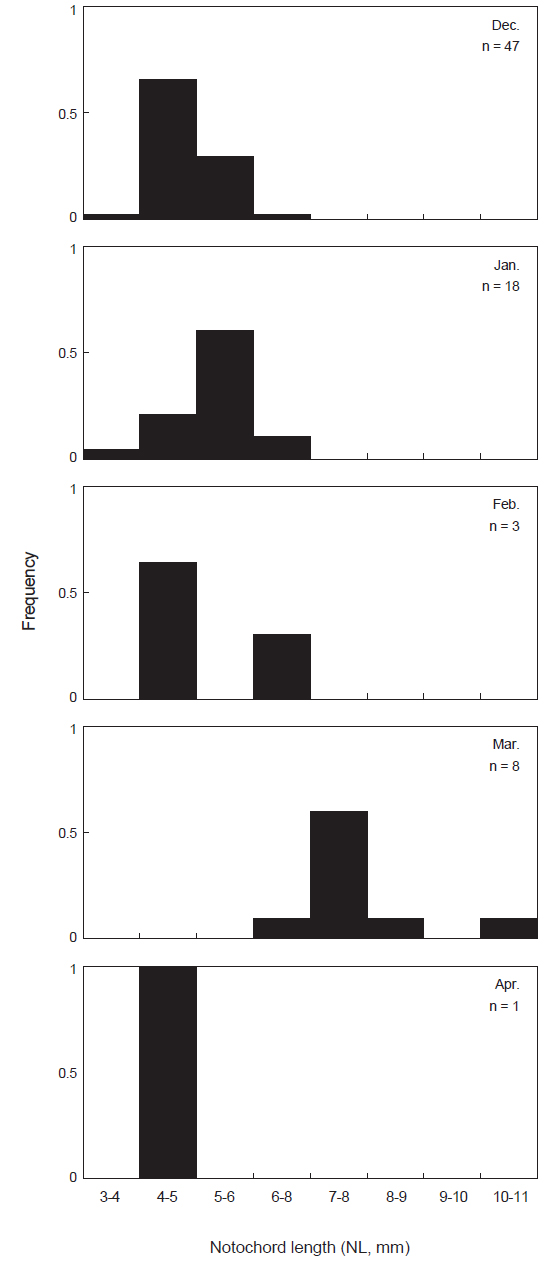
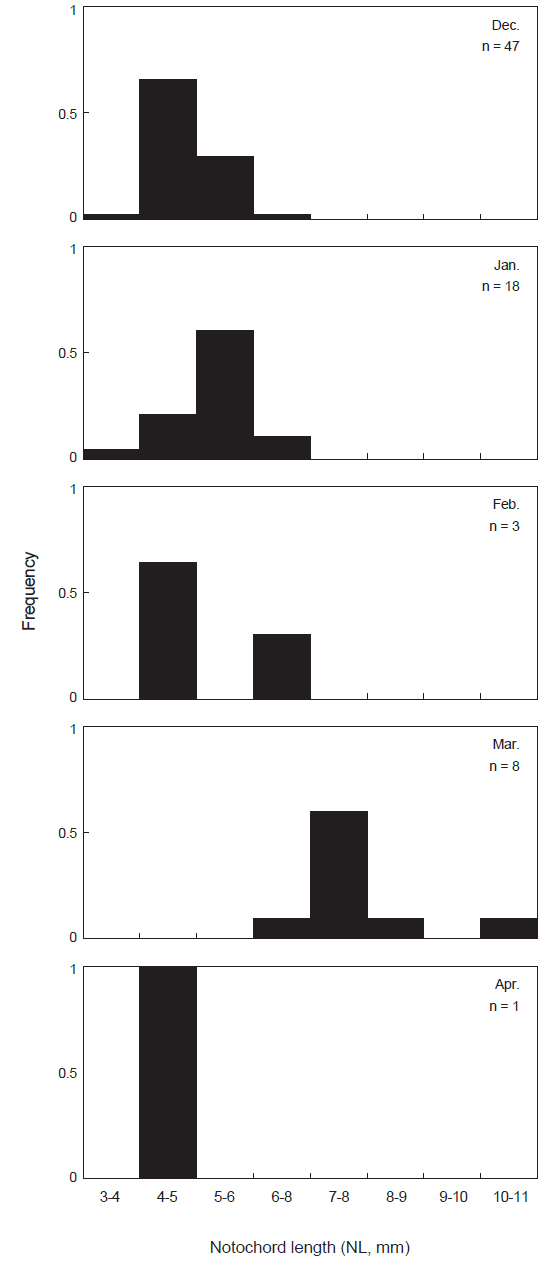
본 연구에서 꼼치 자치어는 2010년 12월부터 2011년 4월까지 총 77개체가 출현 하였으며, 척색장은 3.8-10.6 mm의 범위를 보였다. 월별 출현양상을 살펴보면(Fig. 2), 2010년 12월에는 출현 기간 중 가장 많은 47개체가 채집되어 전체 출현량의 61.0%를 보였다. 이 시기 동안 꼼치 자치어는 3.8-6.2 mm의 척색장 분포를 보였고, 그 중 4-5 mm 크기의 개체가 66.0%로 가장 많이 출현하였다. 2011년 1월에는 18개체가 출현하여 출현기간 중 두 번째로 많은 개체가 출현하였으며, 5-6 mm 크기의 개체가 많이 출현하였다. 2월에는 3개체가 채집되었으며, 3월에는 8개체가 채집되었고 출현하는 꼼치 자치어의 척색장이 점차 증가하는 양상을 보였다. 4월에는 오직 1개체만 채집되었고, 척색장도 4.6 mm로 작은 개체가 출현하였다.
조사 해역에서 꼼치 자치어의 월별 정점별 출현양상을 살펴보면, 12월의 하구역의 안쪽 정점에서는 3마리만 출현하여 12월 전체 출현량의 6.4%를 차지하였고, 대부분이 수온과 염분이 더 높은 하구역의 바깥 정점에서 출현하였으며, 이런 특징은 전 출현 시기에 걸쳐 나타나 꼼치 자치어는 하구의 바깥 정점에서 주로 출현하는 경향을 보였다.
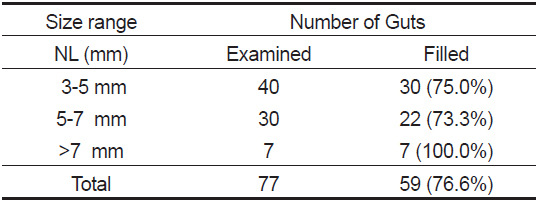
장내용물을 분석한 77개체 중 먹이를 섭식한 꼼치 자치어는 59마리로 76.6%의 섭식률을 보였다(Table 1). 척색장에 따른 섭식참여율은 3-5 mm에서 75.0%로 비교적 높은 섭식률을 보였고, 5-7 mm에서는 73.3%, >7 mm에서는 100%의 섭식률을 보여 척색장이 증가함에 따라 증가하는 경향을 보였다.
[Table 1.] Feeding incidence of Liparis tanakae collected in the Nakdong River estuary
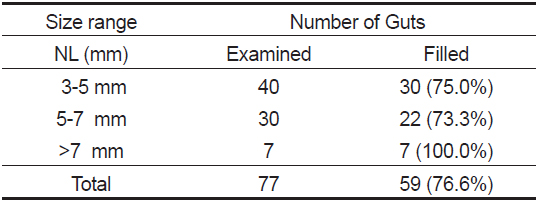
Feeding incidence of Liparis tanakae collected in the Nakdong River estuary
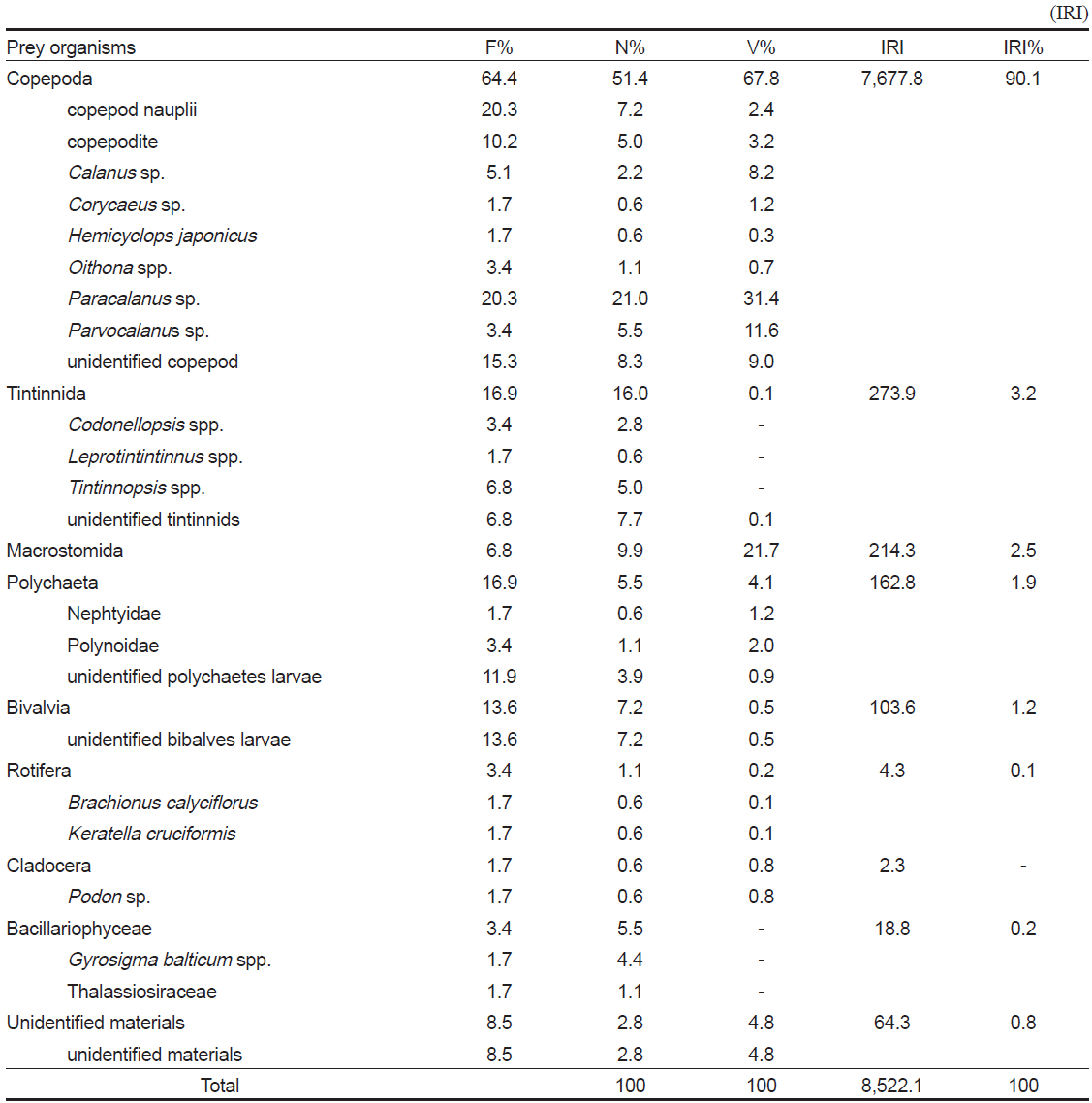
먹이를 섭식한 59개체의 장내용물 분석 결과(Table 2), 꼼치 자치어의 가장 중요한 먹이생물은 출현빈도 64.4%, 개체수비 51.4%, 부피비 67.8%를 보여 상대중요성지수비 90.1%를 차지한 요각류(Copepoda)이었다. 요각류 중에서
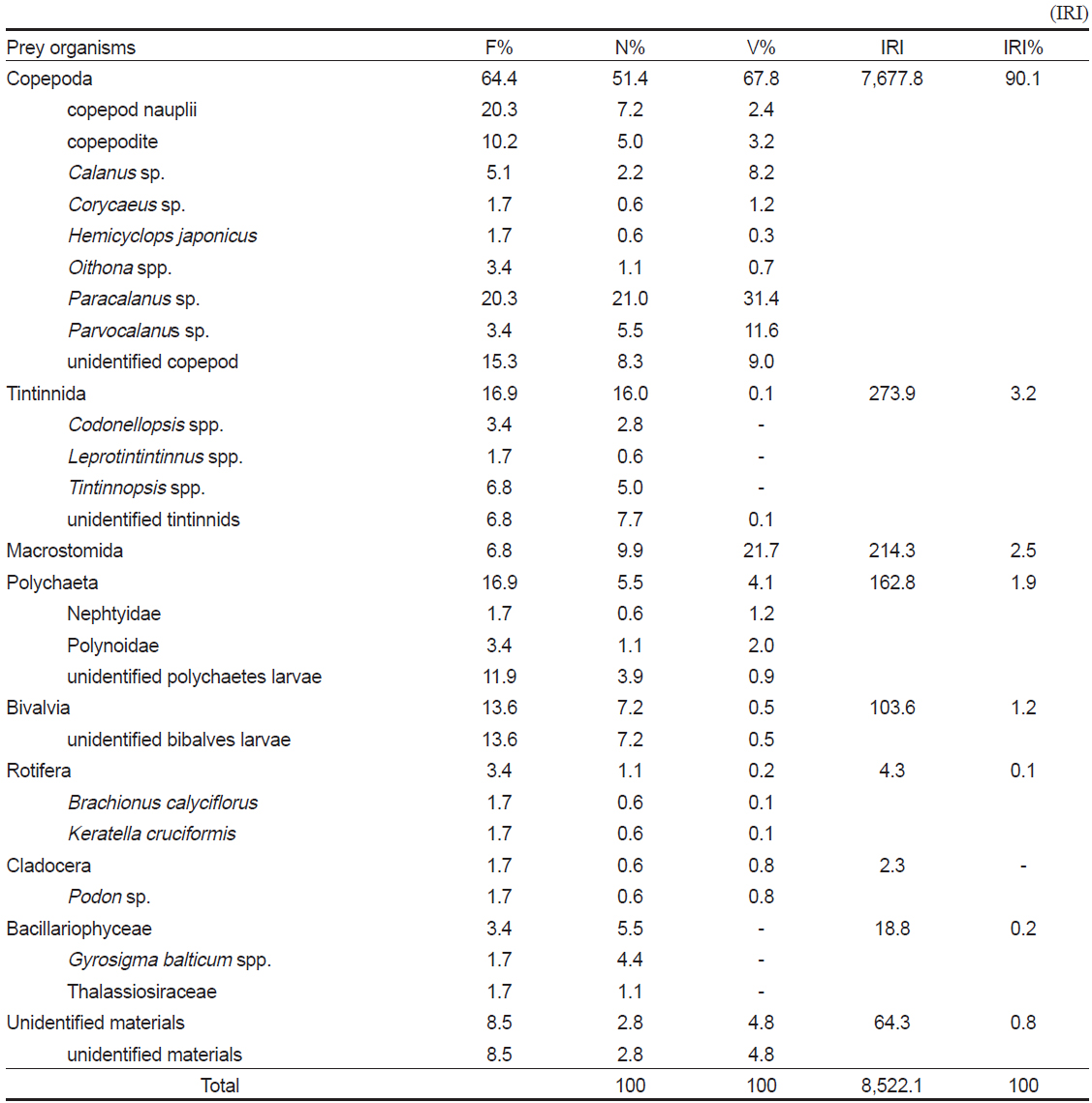
Composition of the gut contents of Liparis tanakae by frequency of occurrence, number, volume and index of relative importance
요각류 다음으로 많이 섭식된 먹이생물은 출현빈도 16.9%, 개체수비 16.0%, 부피비 0.1%를 차지하여 상대중요성지수비 3.2%를 보인 유종섬모충류(Tintinnida)이었다. 유종섬모충류 중에서 미동정 개체가 가장 많았고, 그 외
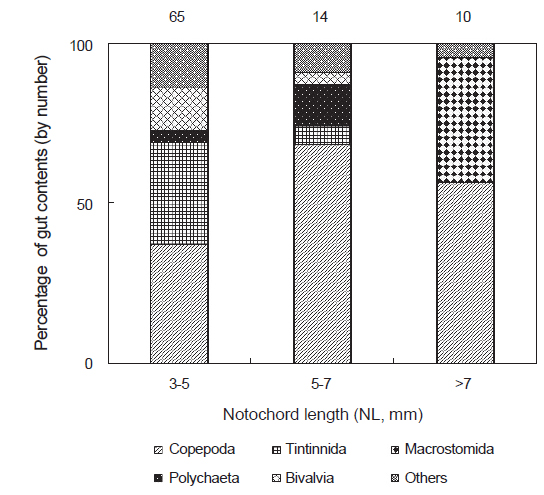
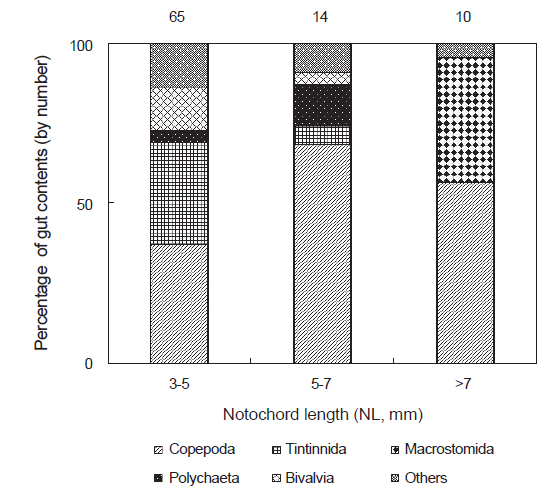
채집된 꼼치 자치어는 3-5 mm, 5-7 mm, >7 mm 3개의 크기군으로 나누어 장내용물의 개체수를 기준으로 먹이생물 조성변화를 조사하였다(Fig. 3). 가장 작은 크기군인 3-5 mm에서는 요각류가 전체 먹이생물 개체수의 37.0%를 차지하여 가장 중요한 먹이생물로 나타났으나, 유종섬모충류 또한 32.1%의 높은 개체수비로 많이 섭식되었다. 그 다음으로 이매패류와 기타 먹이생물이 각각 13.6%로 비교적 많이 섭식되었으며, 다모류가 3.7%로 소량 섭식되었다. 5-7 mm 크기군에서는 요각류가 더욱 증가하여 전체 먹이생물 개체수의 68.5%를 차지하였다. 한편 유종섬모충류는 크게 감소하여 5.6%를 차지하였고, 이매패류와 기타 먹이생물도 감소하였다. 반면 갯지렁이류 유생은 13.0%를 차지하여 이전 크기군에 비하여 증가하였다. 가장 큰 >7 mm 크기군에서는 요각류가 개체수비 56.5%로 다시 감소하는 경향을 보인 반면, 이전 크기군에서 출현하지 않았던 거구목의 편형동물이 개체수비의 39.1%로 중요한 먹이생물이었다. 그 외 기타 먹이생물이 4.3%로 소량 섭식되었다.
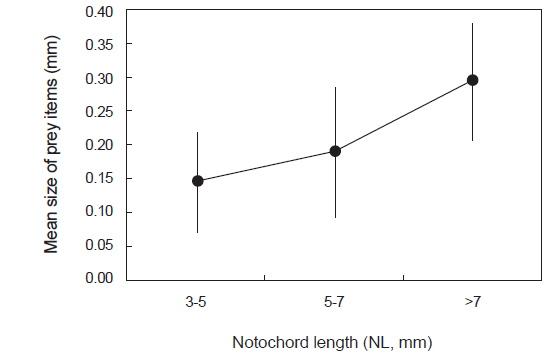
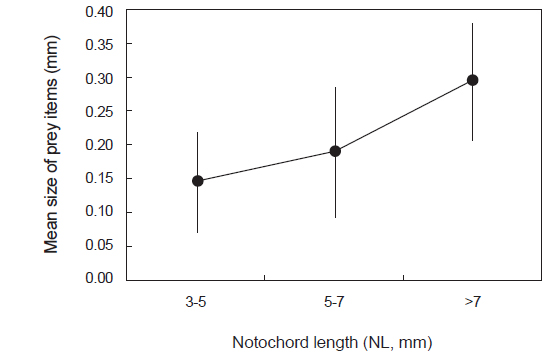
섭식된 먹이생물의 크기를 살펴보면(Fig. 4), 3-5 mm 크기군에서는 평균 먹이생물 크기가 0.15 mm이었으며, 5-7 mm 크기군에서는 0.19 mm, 가장 큰 크기군인 >7 mm에서는 0.29 mm로 자치어의 척색장 증가에 따라 섭식된 먹이생물의 크기는 통계적으로 유의하게 증가하였다(F2,88=10.449,
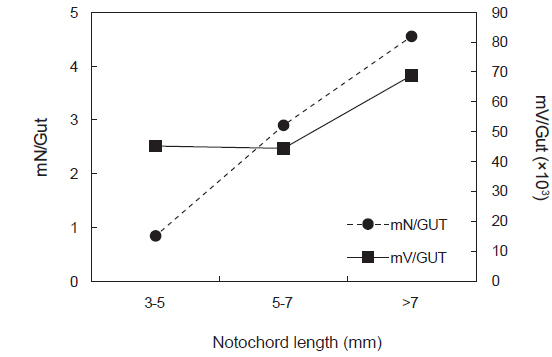
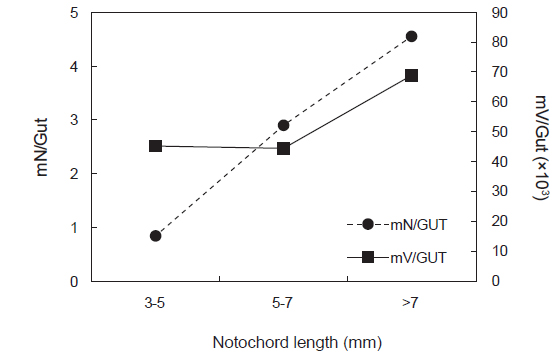
꼼치 자치어의 크기군별 개체 당 평균 먹이생물 개체수(mN/GUT)는 유의한 차이를 보였으며(ANOVA, F2,56=11.118,
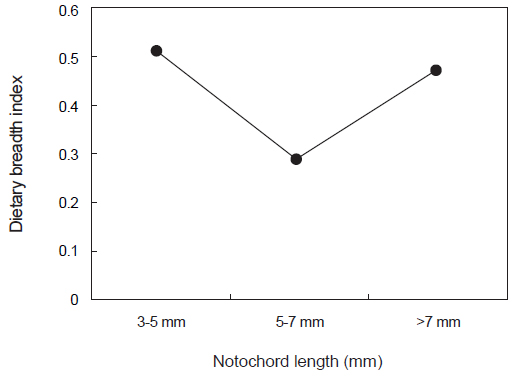
꼼치 자치어에 의해 섭식된 장내용물의 dietary breadth index의 범위는 0.29-0.51이었다(Fig. 6). 가장 작은 크기군인 3-5 mm 크기군에서 가장 높은 값인 0.51을 보였으며, 이후 크게 감소하여 5-7 mm 크기군에서는 가장 낮은 지수 값인 0.29를 보였으나, >7 mm 크기군에서는 0.47로 다시 증가하는 경향을 보였다.
꼼치 자치어의 섭식 초기인 척색장 3-5 mm 크기군의 섭식률은 75.0%였으며, 자치어 기간 동안 전체 섭식률은 76.6%로 비교적 높은 섭식참여율을 보였다. 국내에서 연구된 다른 종들의 자치어 섭식률을 살펴보면, 농어목에 속하는 보구치(
본 연구에서 꼼치 자치어는 상대중요성지수비 90.1%를 보인 요각류를 주로 섭식하였다. 요각류는 많은 종류의 어류들이 선호하는 먹이생물 중 하나로, 주로 어린 시기에 주 먹이생물로 섭식되는 중요한 동물플랑크톤 분류군이다. 광양만(Park, 1999)에서 채집된 망둑어과, 풀반지속의 후기자어 역시 요각류의 노플리우스 알과 유생을 주로 섭식하였으며, 보구치(Cha and Park, 2001a), 멸치(Park and Cha, 1995)와 전어(Park et al., 1996)의 후기자어도 유종섬모충류와 함께 요각류를 많이 섭식하였다. 또한 인도양 동부의 Andaman해에서 채집된 4종의 자치어(
꼼치 자치어의 가장 큰 크기군에서 요각류 다음으로 많이 섭식된 먹이생물인 Macrostomida는 편형동물문(phylum Platyhelminthes) 와충강(class Tubellaria)에 속하는 중형저서생물(meiobenthos)이다. 와충강은 크기가 작고 납작하며 길쭉한 형태의 벌레모양 생물로 개체수가 매우 풍부하게 나타난다(Nybakken and Bertness, 2005). 본 연구해역에서 겨울철에 주로 출현하는 망둑어과, 청어(
본 연구에서 꼼치 자치어는 성장에 따라 주먹이의 섭식량과 기타먹이 생물의 조성이 변하는 결과를 보였다. 꼼치는 전체 크기군에서 요각류를 주로 섭식하였으나, 가장 작은 크기군에서는 유종섬모충류와 이매패류가 각각 개체수비 32.1%, 13.6%로 비교적 많이 섭식되었다. 한편 5-7 mm 크기군에서는 요각류가 68.5%의 개체수비로 가장 많이 섭식되었고, 이전 크기군에서 비교적 많은 섭식량을 보였던 유종섬모충류와 이매패류의 섭식량은 크게 감소하였다. 대신 갯지렁이 유생이 개체수비 13.0%를 보여 비교적 많이 섭식되었다. 그러나 가장 큰 크기군에서는 요각류 외에 Macrostomida가 39.1%의 개체수비를 보여 갯지렁이 유생 및 기타먹이를 대체하는 경향을 보였다. 또한 자치어 성장에 따른 먹이 조성변화 외에도 척색장과 먹이생물의 크기 사이에 통계적으로 유의한 양의 상관관계를 보였으며(Fig. 4), 다양한 연구에서 이와 유사한 결과들이 보고된바 있다(Young and Davis, 1990; Pepin and Penny, 1997; DeVries et al., 1998; Sabates and Saiz, 2000; Østergaard et al., 2005). 이런 결과는 자치어가 성장함에 따라 더 많은 에너지 요구에 부합해 더 큰 먹이생물을 잡아먹을 수 있는 능력을 가지게 되면서 먹이전환을 하는 것으로 생각된다.
국내외 연구 중, 꼼치 자치어 시기의 식성 연구는 찾아 볼 수 없었지만, 광양만에서 채집된 꼼치 유어(2.2-5.8 cm SL; SL:Standard Length)의 식성과(Kwak and Huh, 2003), 본 연구 해역과 동일한 낙동강 하구역에서 채집된 꼼치(4-45 cm SL)의 식성(Huh, 1997)에 대한 연구가 이루어져 있다. 꼼치 유어는 단각류(Amphipoda)를 가장 많이 섭식하였고, 그 중 카프렐라류(Caprellidea)를 많이 섭식하였다. 그 외 옆새우류(Gammaridea)와 새우류(Caridea) 등의 갑각류를 주로 섭식하였다. 그러나 체장 2.5 cm 미만의 개체들은 요각류가 전체 위내용물 중량의 45.2%를 차지하여 우점하였고, 이 후 4 cm 이상의 크기에서는 새우류를 주로 섭식하여 Huh (1997)의 연구와도 일치하였다. 따라서 꼼치는 성장에 따라 요각류에서 단각류로, 단각류에서 새우류로 이어지는 먹이전환을 겪는 것으로 생각되며, 이런 먹이전환은 종 내의 먹이 경쟁을 감소시켜 다양한 크기의 개체들이 공존하게 하는 기작으로 작용하는 것으로 생각된다(Langton, 1982).