경골어류의 난소발달 과정을 조절하는 주요 내분비호르몬은 뇌하수체의 생식소자극호르몬(GtHs, gonadotropins)인 여포자극호르몬(FSH, follicle stimulating hormone)과 황체형성 호르몬(LH, luteinizing hormone), 그리고 난모세포의 여포층에서 생성.분비되는 성 스테로이드 호르몬이다(Lubzens et al., 2010; Kagawa, 2013).
난소내 난황이 축적되기 시작하는 난황형성시기에 혈장 FSH 농도가 증가하면서 estradiol-17β(E2)가 이 시기의 주요 스 테로이드로 작용하고, 성숙과 배란시기에는 혈장내 LH 농도 증가와 함께 progesterone의 유도체인 17α, 20β-dihydroxy-4-pregnen-3-one(17α20βOHP)나 17,20β,21-trihydroxy-4-pregnen-3-one (17α20β21P)이 주요 스테로이드 호르몬으로 알려져 있다(Nagahama et al., 1994; Yoshizaki et al., 2001; Patino et al., 2003; Lubzens et al., 2010; Kagawa, 2013). 지금까지의 연구를 보면 난황형성기에는 난모세포에서 생성된 E2가 간에서 난황전 구물질(vitellogenin, Vg)을 합성한 후 난모세포내에 난황물질(vitellin)로 전환되어 축적되므로 난황형성과 E2의 상호 연관성에 대해서는 이미 잘 알려져 있으며, 최근에는 E2의 생성에 성장 호르몬의 관련성도 제기되고 있다(Babin et al., 2007; Lubzens et al., 2010). 그러나 난모세포의 성숙과 관련된 progestogens 생성과 기작에 대한 연구는 여전히 부족한 실정이다. 어류의 난성숙과정은 난황형성기에 비해 단기간에 이루어지며, 난핵포 붕괴(GVBD, germinal vesicle break down) 직후 배란이 일어나는 일련의 과정에 어떠한 호르몬들이 관련되어 상호작용하는지 또한 어류의 산란 패턴, 즉, 산란기에 한번 산란하는 어종, 2번 이상 다회 산란하는 어종에 따라서도 호르몬 종류와 작용 기작이 다르므로 더 많은 연구가 요구된다.
기름가자미는 가자미목 가자미과에 속하는 동해안의 상업종으로 우리나라 동·남해, 일본, 사할린, 동중국해에 분포한다(Choi et al., 2002; National Fisheries Research and Development Institute, 2004; Kim et al., 2005). 동해안의 기름가자미는 자원상태가 양호하나 최근의 기후변화 및 해양환경 변화로 자원관리가 필요한 종으로 인식되어 이를 위한 기초자료로 Cha et al. (2008)이 동해안 기름가자미의 성숙과 산란에 대하여, 그리고 Lee et al. (2014)이 어란과 자어 출현 등 기름가자미의 산란생태 특성에 대하여 조사하였다. 이와 같은 자료를 근거로 본 연구에서는 산란기의 개체들을 대상으로 성숙 관련 대사물질을 분석함으로써 기름가자미의 산란 패턴과 산란 회수를 정확히 파악하는데 기초자료로 제공하고자 한다.
실험어는 산란기인 2013년 5월과 6월에 경북 울진 부근 해역에서 어획된 기름가자미 개체들 중 성숙 단계라고 판단되는 암컷 4 개체를 대상으로 하였다. 실험에 사용된 개체의 전장은 30.6-32.0 cm, 체중은 198-269 g 이었다. 실험어는 전 채혈 후 무균상태에서 난소를 절취하여 ice-cold balanced salt solution (132.96 mM NaCl, 3.09 mM KCl, 0.28 mM MgSO4·7H2O, 0.98 mM MgCl2·6H2O, 3.40 mM CaCl2·6H2O, 3.65 mM HEPES)으로 세척하였다. 절취된 난소는 가는 핀셋을 이용하여 개체별로 난모세포를 각각 분리하였다.
분리된 난모세포는 난경별(0.76, 0.82, 0.88, 그리고 0.91 mm)로 24-well plate에 well당 Leibovitz의 L-15 (Gibco, U.S.A.) 1 mL에 20개의 난모세포를 각각 분주한 후 18℃에서 24시간 배양하였다. Balanced salt solution과 L-15 배양액의 pH는 7.7, 삼투농도는 380 mOsmol kg-1로 조절하였다. 난모세포에서 생성되는 성 스테로이드 호르몬 종류를 분석하기 위해 방사 표지된 전구물질인 17α-hydroxyprogesterone ([3H]-17αOHP, Amersham Life Science, England)을 난경별로 well당 각각 55 kBq 첨가하여 배양하였다.
난모세포 발달 단계의 조직학적 관찰을 위해 각 개체로부터 적출한 난소를 Bouin's solution에 고정하여 파라핀으로 포매한 후 5-6 μm 두께의 절편을 만들어 Mayer's hematoxylin과 eosin으로 비교 염색하여 광학현미경(U-ND25-2, OLYMPUS, JAPAN)하에서 관찰하였다.
배양 후 난모세포와 배양액은 함께 80% ethanol로 균질화하여 원심분리한 후 상등액만을 모아 건조시킨 뒤 증류수 500 μL에 용해시켜 dichloromethane으로 2번 추출하여 유리 스테로이드만을 얻었다.
스테로이드 추출물은 스테로이드 표준물질과 동시에 실리카겔을 입힌 얇은 막 지지체(TLC plate, 60F254, Merck)에 점적시킨 후 밀폐된 혼합용매(benzene:acetone=80:20와 benzene:ethylacetate=80:20)속에서 전개시켰으며, 이후 TLC plate상의 대사물질 확인은 estrone (E1)과 estradiol-17β (E2)의 경우 요오드 증기로, 나머지 반점들은 254 nm의 자외선 하에서 이루어졌다. TLC plate에 나타난 대사물질들은 방사선 사진법(autoradiography, Fuji Bas 3000)으로 재확인하였으며, 재확인된 반점에 해당되는 실리카겔 밴드를 잘라 5 mL의 혼합 용매(dichloromethane : methanol = 9:1)로 용리하였다. 용리된 대사물질은 HPLC (high performance liquid chromatography, Waters)를 이용하여 표준물질의 retention time과 일치하는 분획만을 분리하였다. 분리된 분획은 Liquid Scintillation Counter (PACKARD TR-CARB-2100T, USA)를 이용하여 radioactivity 측정 후 대사율을 계산하였으며, 분리된 대사물질은 HPLC-radiodetector (β-RAM MODEL 4, USA)로 재확인하였다. HPLC-radiodetector로 확인된 대사물질은 gas chromatography-mass spectrometry (GC/MS) (GCMS-QP5050A, Shimadzu, Japan) 로 동정하였으며, GC/MS 작동조건은 table 1과 같다.
[Table 1.] Analytical conditions of GC/MS for identification of steroid metabolites
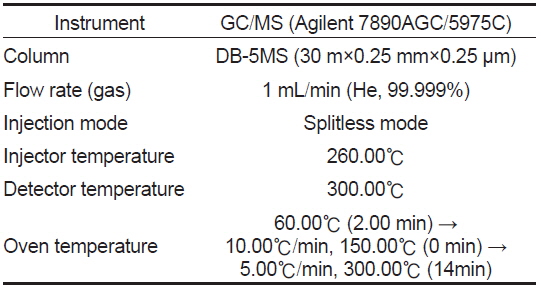
Analytical conditions of GC/MS for identification of steroid metabolites
>
성숙 난모세포의 난경별 조직 관찰과 스테로이드 대사 물질 분석
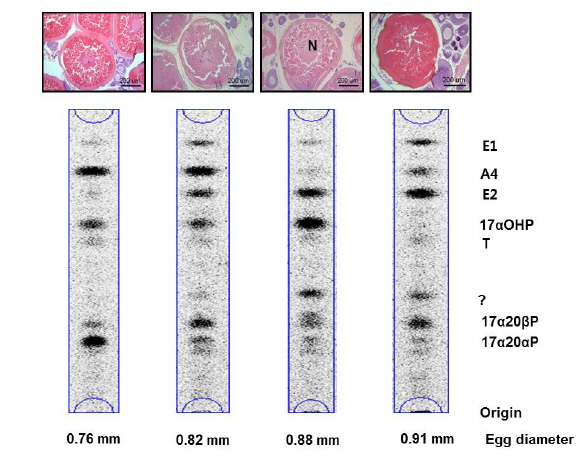
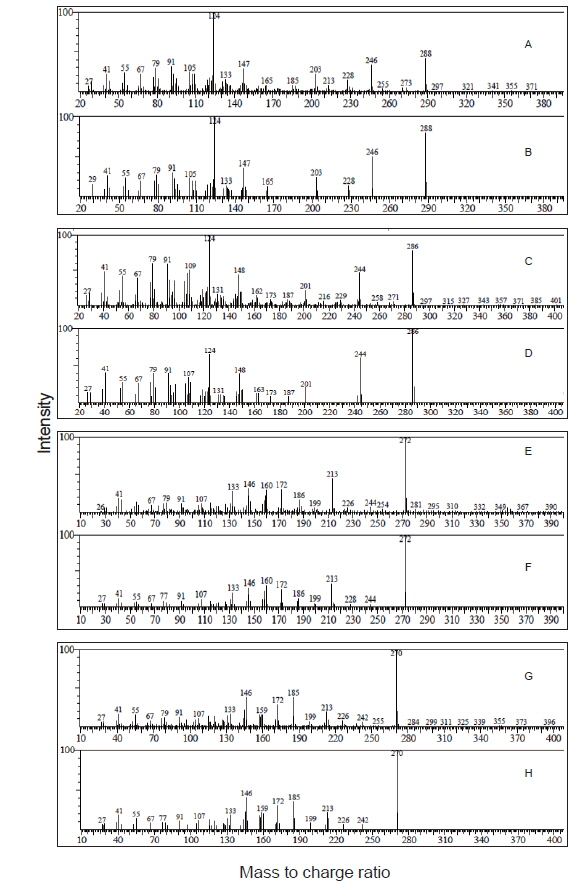
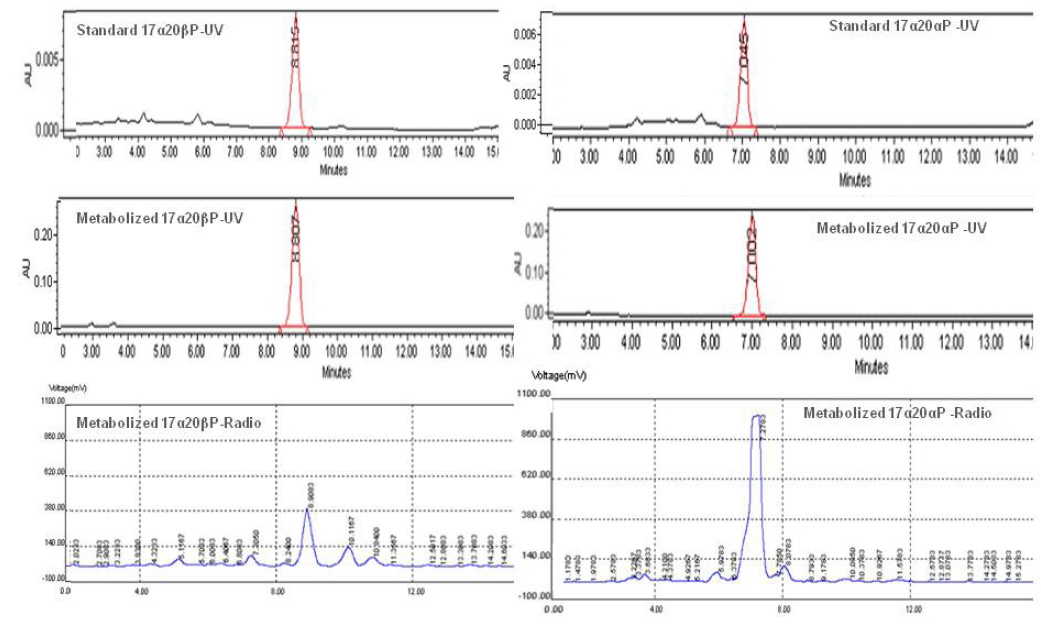
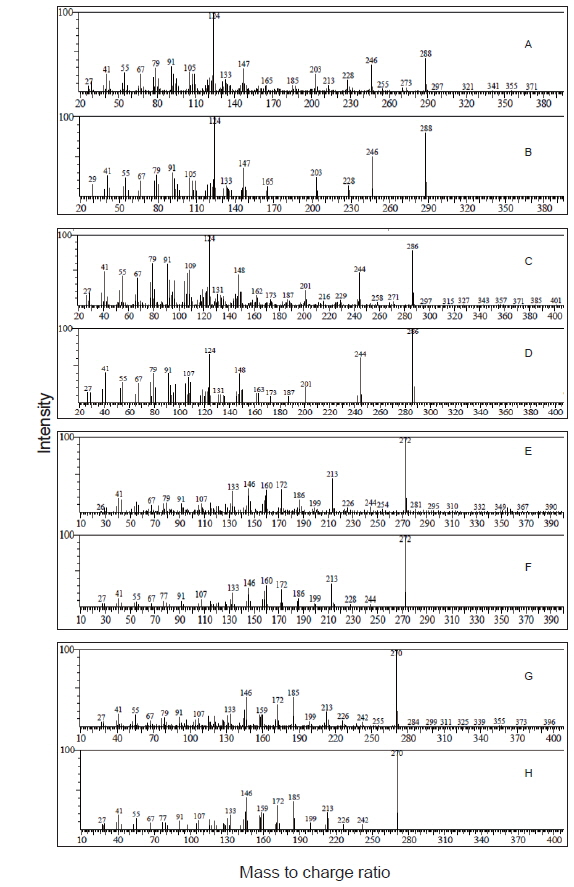
기름가자미 난소내 난모세포 배양직전의 조직 관찰은 Fig. 1과 같다. 평균 난경 0.76, 0.82, 0.88 그리고 0.91 mm에 해당하는 난모세포들의 세포질 내에는 난황과립(yolk granule, Yg)으로 충만하였으며, 성숙 정도를 판단할 수 있는 핵(nucleus, N)의 이동상을 보면 0.76 mm과 0.82 mm 의 난모세포는 세포질 중앙에 핵이 위치하고 있었으며, 난경 0.88 mm에서는 핵이 동물극 방향으로 이동 중이며, 0.91 mm인 난모세포는 GVBD 직전 단계인 것으로 관찰되었다. Fig. 1에서 보는 바와 같이, 이들 난모세포들을 대상으로 성숙 정도에 따른 스테로이드 호르몬 존재 여부를 알아보기 위하여 전구물질인 [3H]-17αOHP 으로부터 생성된 스테로이드 대사물질을 분석하였다. TLC상에 전구물질로부터 전환된 스테로이드 반점은 17α20αP, 17α20βP, A4, T, E2 그리고 E1 등의 표준물질과 일치하였으며 평균난경 0.88과 0.91 mm에서는 unknown (?) 물질도 관찰되었다. TLC 상의 반점들은 용리(elution)후 HPLC를 이용하여 스테로이드 분획을 분리 후 HPLC-radiodetector를 거쳐 GC-MS에 의해 최종 동정되었다. [3H]-17αOHP로부터 전환된 대사물질들 중 A4, T, E2 그리고 E1 만이 GC-MS상에서 최종 동정되었다(Fig. 2). 17α20αP와 17α20βP은 HPLC상에서만 재확인되었으며(Fig. 3), 0.88과 0.91 mm 에서 관찰된 unknown 물질은 GC-MS에 의한 동정에도 불구하고 확인되지 못하였다.
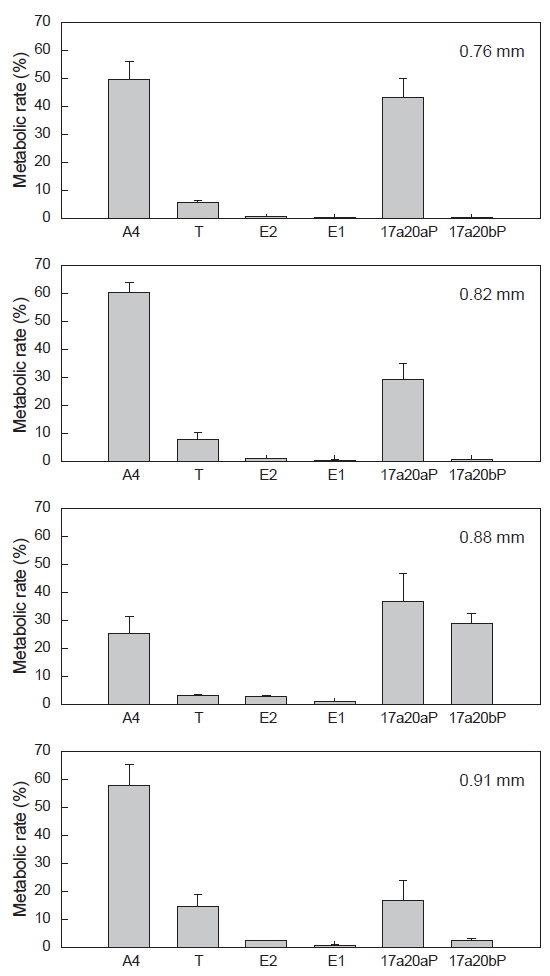
전구물질, [3H]-17αOHP로부터 전환된 스테로이드 대사물질들의 대사율은 Fig. 4와 같다. A4의 대사율은 평균난경 0.76, 0.82 mm인 난모세포에서 각각 49.7, 60.2% 였고 0.88 mm인 난모세포에서 25.1%로 낮았으며, 0.91 mm에서 다시 57.7%의 대사율을 보였다. T로의 대사율에서는 평균난경 0.88 mm에서 3.07%로 가장 낮게 나타났다. E2의 대사율은 평균난경 0.76과 0.82 mm에서 각각 0.7과1.0%였고, 0.88과 0.91 mm 난모세포에서 2.7과 2.4%의 대사율을 나타냈다. E1의 대사율은 난경 모두에서 0.18-0.96%로 나타났다. 프로제스테론의 유도체인 17α20αP와 17α20βP의 경우, 17α20βP는 핵이 동물극 방향으로 이동 중인 난경 0.88 mm에서 29.04%로 가장 높은 대사율을 보이다가 GVBD 직전인 0.91 mm에서는 낮은 수치(2.61%)를 나타냈다. 17α20αP은 0.76-0.88 mm 난모세포에서 비교적 높은 대사율(29.3-43.0%)을 보이다가 0.91 mm에서 16.8%로 감소하였다.
본 연구에 사용된 성숙기 난모세포는 기름가자미(
Canario and Scott (1990, 1991)는 17α20αP가 marine flatfish
난황형성기의 주요 스테로이드인 estrogens (E2와 E1)은 성숙 단계의 난경 크기(0.76-0.91 mm)에서는 낮은 대사율을 보인 반면 testosterone (T)은 핵이동 이후의 최종성숙 단계인 0.91 mm에서 높게 나타났다. 연어과 어류에서 혈장 T 농도는 난황형성기 동안 E2와 유사하게 증가하다가 성숙과 배란 시기에 높은 농도로 유지되는 것은 T가 E2의 전구물질로 T→ E2로 전환되는 aromatase의 활성 감소와 관련된다고 하였다(Kagawa, 2013). 참돔(
본 연구에서 성숙 과정중인 난모세포를 대상으로(0.76, 0.82, 0.88 그리고 0.91 mm) 성 스테로이드 대사물질 종류와 이들 대사율 분석 결과, 핵이동 시기인 0.88 mm 난모세포 크기에서 대사물질의 변화가 일어나 최종성숙인 GVBD에 반응하는 단계의 난경이라고 판단되며, 비록 이 시기에 관찰된 unknown 물질을 동정하지 못했지만 기름가자미의 최종성숙 유도 물질은 17α20βP인 것으로 생각되었다. 아울러 난모세포의 성숙과 배란 메카니즘 연구를 위해 또 다른 스테로이드의 존재 가능성에 대해서도 계속적인 연구가 요구된다.

![Autoradiograms of steroid metabolites produced from [3H]17α-hydroxyprogesterone after 24 hours incubation with isolated oocytes (20 oocytes/mL/well) from blackfin flounder Glyptocephalus stelleri. The metabolites were separated by thin layer chromatography developed with a benzene : acetone (4:1) and benzene : ethyl acetate (4:1) solvent system. A4,androstenedione; E1, estrone; E2, estradiol-17β; T, testosterone; 17α20αP, 17α,20α-dihydroxy-4-pregen-3-one; 17α20βP, 17α20β-dihydroxy-4-pregnen-3-one; ?, unknown. N, nucleus.](http://oak.go.kr/repository/journal/17554/KSSHBC_2015_v48n4_483_f001.jpg)
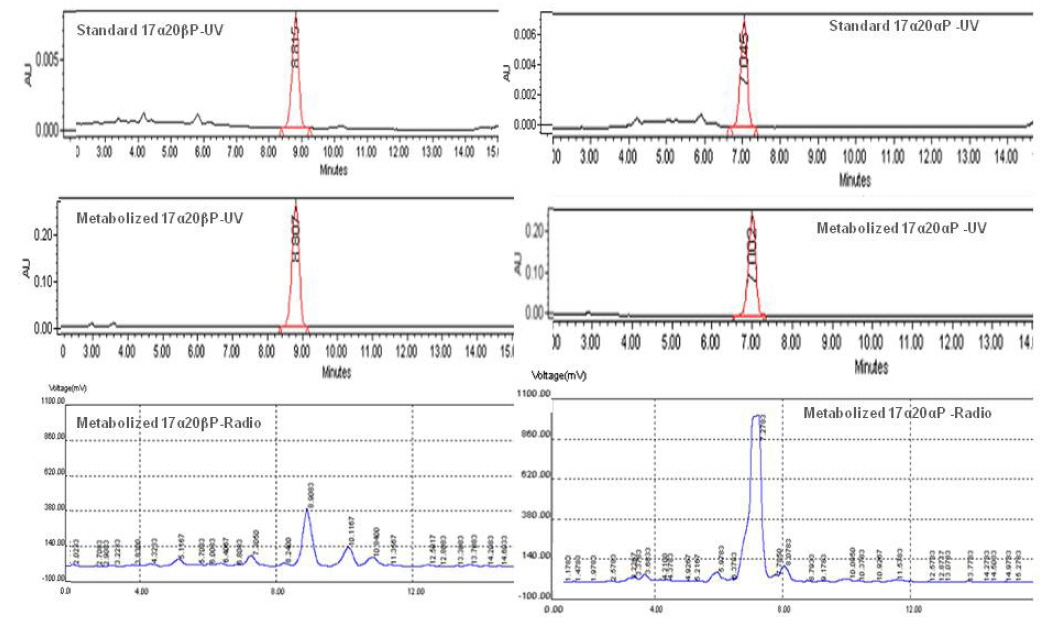
![Radioactivities of steroid metabolites from [3H]17α-hydroxyprogesterone in blackfin flounder Glyptocephalus stelleri. oocytes. The percentage of radioactivity associated with each isolated steroid was calculated to the percentage of total steroid recovered from initial TLC. Values are mean±SE (in duplicate wells, 20 oocytes/well). A4, androstenedione ; E1, estrone; E2, estradiol-17β; T, testosterone; 17α20αP, 17α,20α-dihydroxy-4-pregen-3-one17α20βP, 17α20β-dihydroxy-4-pregnen-3-one.](http://oak.go.kr/repository/journal/17554/KSSHBC_2015_v48n4_483_f004.jpg)