기후변화가 해양생태계에 미치는 영향에 대한 연구는 해양 생태학 뿐만 아니라 수산 자원관리 분야에서도 중요한 주제가 되고 있다. 북태평양해양과학기구(The North Pacific Marine Science Organization, PICES)는 북태평양에서 발생하였던 기후와 해양의 물리적 현상의 변이, 그리고 이에 따른 생태계변화에 대해 요약하였으며(PICES, 2004 and 2010), Otterson et al. (2010)은 기후변화로 인한 해양환경의 변동이 여러 해역의 생태계에 미치는 영향에 대하여 보고하였다. 세계적으로 수산자원 관리방침에서 해양생태계의 상태를 고려하는 것은 당연하다는 관점이 생기게 되었으며(FAO, 2003), 미국의 해양정책 입안자들은 1990년대 후반에 동부 태평양에서 발생한 기후체제 변화에 따른 해양의 변화가 무엇인지 규명하는 과학적 질문을 설정하는데 PICES과학자들에게 조언을 요청하기도 하였다(PICES, 2005). 그러므로, 환경변화로 일어난 해양생태계의 변화는 해양생태학자들의 관심분야로 떠오르기 시작했다(Kruse et al., 2006).
가다랑어(
생지화학 역학모델을 이용해 가다랑어의 먹이분포를 예측하였을 때 수렴대(convergence zone)의 동서 이동과 가다랑어의 분포변화는 유의한 상관관계를 보였다(Lehodey et al., 1998). 먹이분포는 웜풀의 가장자리에서 높게 예측되었으며, 가다랑어가 많이 어획되는 해역은 예측된 먹이분포 밀도가 높은 해역과 일치하였다. 지구 온난화 시나리오와 가다랑어 서식해역에 대한 예비 모델링 연구에서 대기 이산화탄소의 농도 상승에 의한 온난화는 열대해역의 가다랑어 먹이생물을 증가시키며 온대해역의 먹이생물은 감소시켜 해수면 온도의 상승으로 인해 가다랑어의 최적 서식해역이 확대되는 결과를 도출하였다(Loukos et al., 2003). 두 모델링 연구는 해양환경 변화로 인해 가다랑어의 분포에 변화가 있을 수 있다는 결과를 보여주지만, 해양환경 변화가 어떠한 생태학적 과정을 거쳐 가다랑어의 생물학적 변화를 일으키는지는 예측하지 못했다.
중서부태평양의 가다랑어는 20세기 중반부터 우리나라를 포함하여 일본, 파푸아뉴기니아, 대만, 그리고 미국의 어선에 의해 많이 어획되고 있다. 가다랑어의 어획량은 태평양 총 다랑어 어획량의 70%를 차지할 정도로 높은 어획량을 기록하고 있지만 다른 다랑어류에 비해 그 연구가 미비하다. 통계적 개체군 동태 모델링(statistical population dynamics model, MULTIFAN-CL)을 통해 추정된 지난 반세기 동안의 가다랑어 가입과 공간환경모델(spatial environmental population model, SEPODYM)을 이용한 가다랑어 서식환경을 비교한 결과로부터 기후와 해양의 지수가 가다랑어의 가입에 때로는 강하게 때로는 약하게 영향을 주고 있음을 알 수 있었다(Lehodey et al., 2003). 가령, 북태평양의 해면수온을 기본으로 한 태평양십년 주기진동(Pacific Decadal Oscillation, PDO)이 양의 값을 나타내었을 때 엘니뇨현상은 자주 발생하며, 이는 가다랑어 가입을 증가시킨다. 또한 온실가스가 산업화 이전의 수준의 4 배가 될 경우 엘니뇨 발생의 빈도가 2 배로 증가한다는 연구 결과가 있으므로(Collins, 2000), 미래 가다랑어의 가입과 어획은 지구 온난화와 상당한 밀접한 결과가 있을 것이라 예측된다.
기후변화가 해양생태계에 영향을 미친다는 사실은 이미 정설로 인식되고 있지만, 환경적 변화가 어떤 메커니즘에 의해, 어느 정도의 시간 차이를 가지고 생물학적 반응을 발현시키는지는 정확하게 연구되지 않았다. 본 논문에서는 한국의 가다랑 어 어획자료를 이용하여 ENSO가 가다랑어 어장의 이동과 어획에 미치는 영향을 고찰해 보았다. 또한 국립수산과학원에서 수집한 생물학적 자료가 기후지수의 변화와 어떤 시차를 가지고 변화하는지 알아보았다.
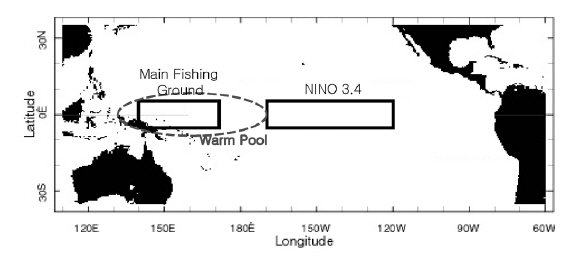
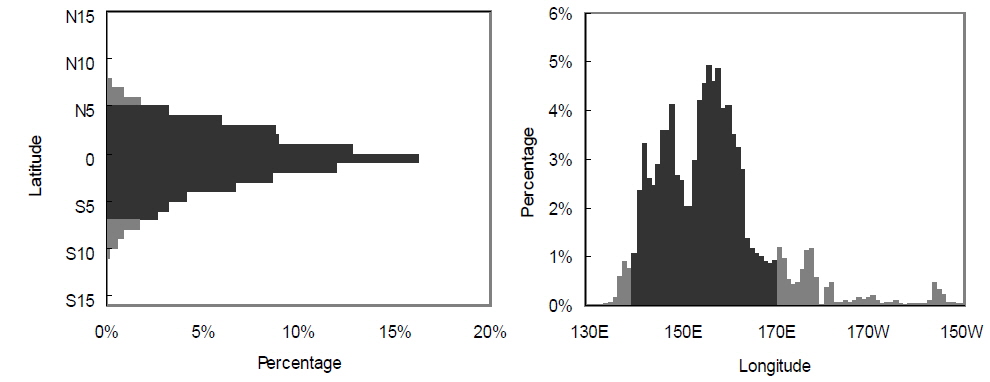
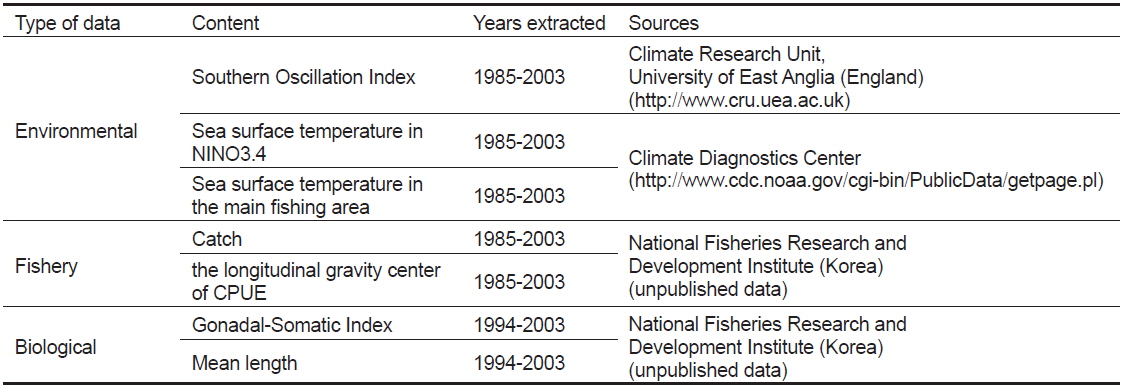
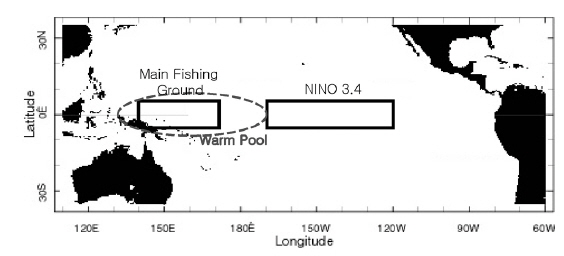
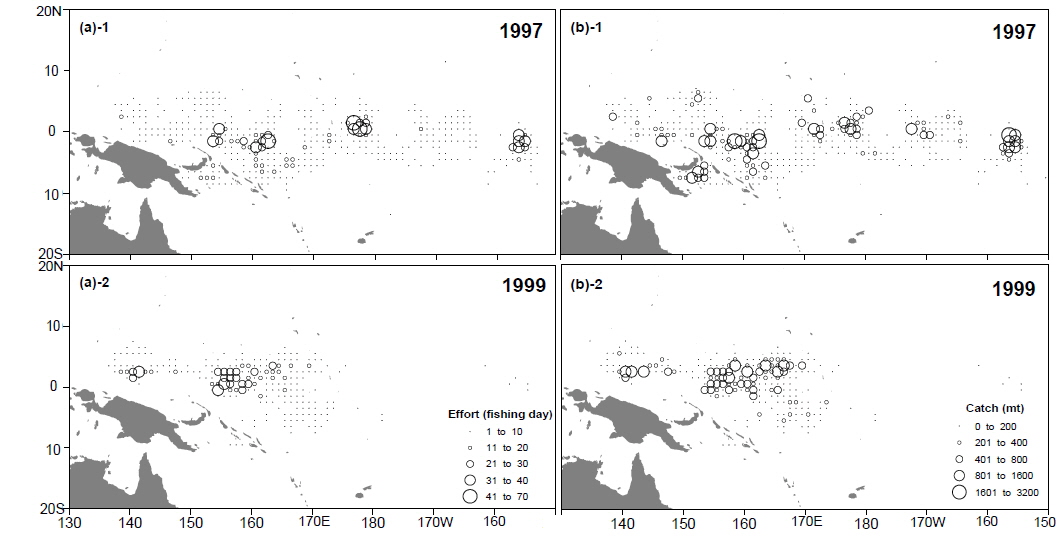
본 연구에서 사용한 환경 자료는 남방진동지수[Southern Oscillation Index, SOI, by Ropelewski and Jones (1987)]와 해면수온(sea surface temperature, SST)이다. 1985년 1월부터 2003년 12월까지의 SOI와어획된가다랑어의 생물학적 특성 을비교하였다. 또한 같은 기간의 NINO3.4 (5°N-5°S, 170°-120°W)해역과 한국 선망어업의 주 어획해역(main fishing area, MAFA; 5°S-5°N, 140° -170°E)의 월별 해면수온을 Climate Diagnostic Center (http://www.cdc.noaa.gov/cgi-bin/PublicData/getpage. pl)로부터 전송받아 본 연구에 사용하였다(Fig. 1). NINO3.4 해면수온(NINO3.4 SST)은 ENSO와 거의 동일한 변화를 보이고 있으므로(Hanley et al., 2003), 이를 ENSO의 지표로 사용하였다. 연구 기간동안의 어획노력이 80% 이상이 이루어지는 해역을 MAFA로 설정하였다(Fig. 2). NINO3.4 와 MAFA의 해면수온 아노말리를 산출하여 각각 NINO3.4 index, MAFA anomaly로 명칭하여 본 연구에 사용하였다.
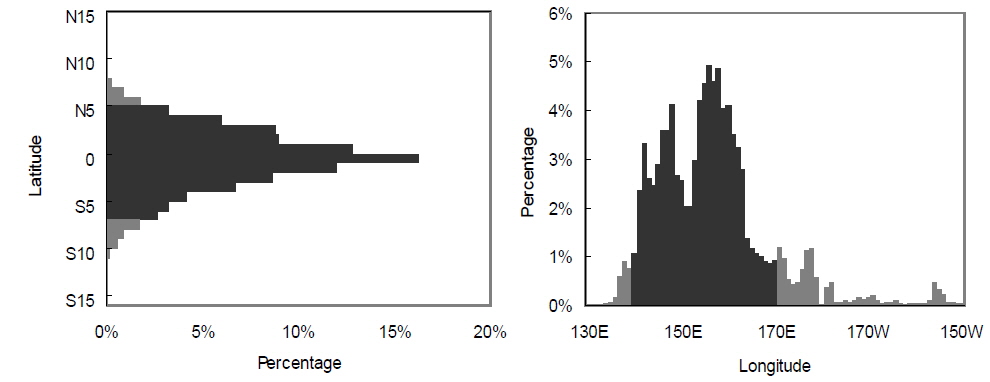
국립수산과학원에서는 우리나라 선망 어업자료를 1980년대 초부터 수집하여 기록하였다. 본 연구를 위해 1985년부터 2003년도의 어획량과 월별 단위노력당 어획량 (catch per unit effort, CPUE, 어획일수당 어획량)이 산출되었으며, 위도 5°S-5°N 내에서의 위경도 1도 단위의 구획으로 어획량을 나누었다. 이를 이용해 단위노력당 경도 어획 중심(

여기서
1994년부터 2003년까지 중서부태평양에서 어획된 가다랑어 중에서 매월 200-300개체를 암수로 나눈 후 가랑이체장, 체중을 측정하였다. 또한, 매월 40-60개체를 선별하여 생식선 중량을 측정하였으며 이를 이용해 생식소중량지수(GSI)를 다음과 같은 식으로 계산하였다.

여기서
본 연구에 사용된 자료는 Table 1에 요약하였으며, 각 자료의 편차보다는 환경자료와 어획, 생물자료 변화의 경향성을 확인하기 위해 모든 자료는 5달 기준으로 이동평균하여 나타내었다. 환경자료와 생물학적 자료의 상관관계는 SPSS 소프트웨어를 이용하여 분석하였다. 환경변화가 가다랑어의 어획과 생물학적 형태에 변화를 주는 시간차를 판단하기 위해 0에서 12개월사이의 교차상관관계(cross-correlation function, CCF)를 구하였다.
[Table 1.] Sources and types of information used in the analyses
Sources and types of information used in the analyses
한국 대형 선망어업은 중서부태평양에서 1980년도 초부터 조업해왔다. 비록 이 해역에서 1985-2003년 동안 한국 어획량은 전체 가다랑어 어획량의 4-24%에 불과하지만, 세계 선망 어획량과 한국 선망 어획량의 변화는 아주 높은 상관관계를 보이고 있다(r=0.901,
[Table. 2.] *: Significant at the 5 % level **: Significant at the 1 % level

*: Significant at the 5 % level **: Significant at the 1 % level
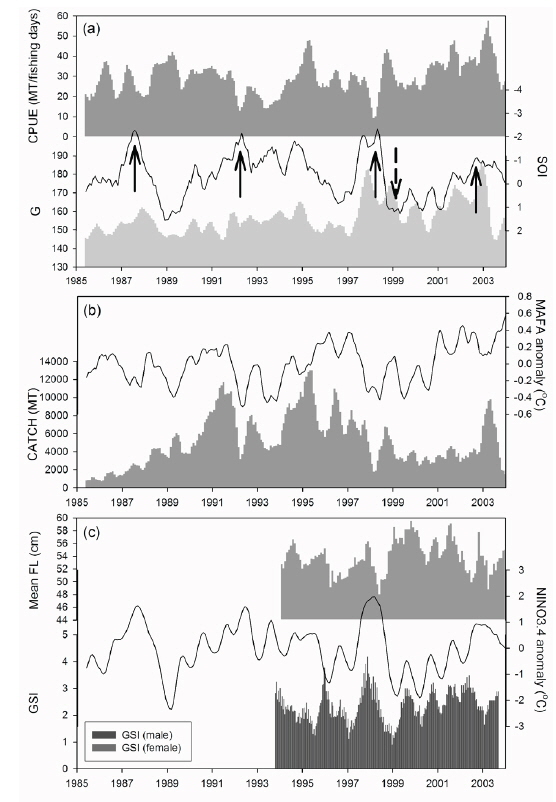
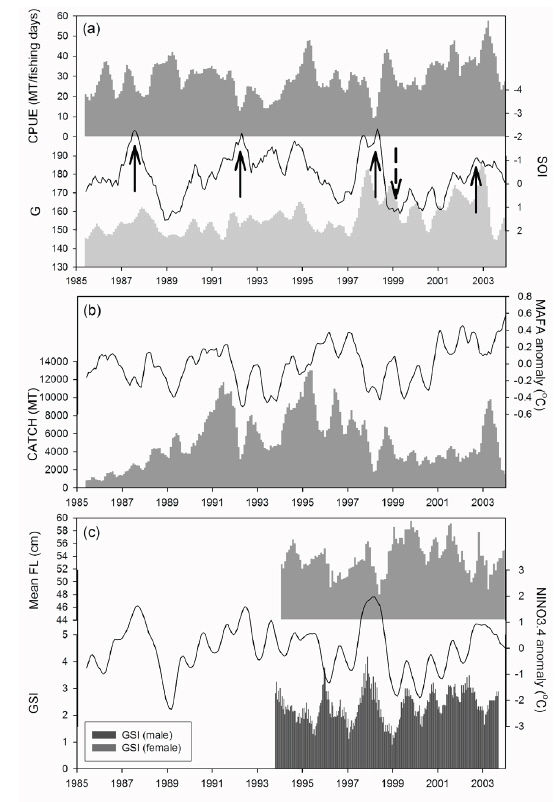
우리나라 선망어업의 가다랑어 총 생산량은 1980년대 중반부터 1990년대 초반까지는 계속 증가하였으나, 그 이후에는 변화폭이 컸다. 특히 1991년에 높은 어획량을 보이던 것이 1992년에는 크게 감소하였으며, 다시 1995년에 크게 증가하였고, 1996년에 감소하는 변동을 보였다(Fig. 3b). 그 이후, 대체로 감소 경향을 보이던 어획량은 다시 2003년에 증가하였다. 하지만, 가다랑어 어획량은
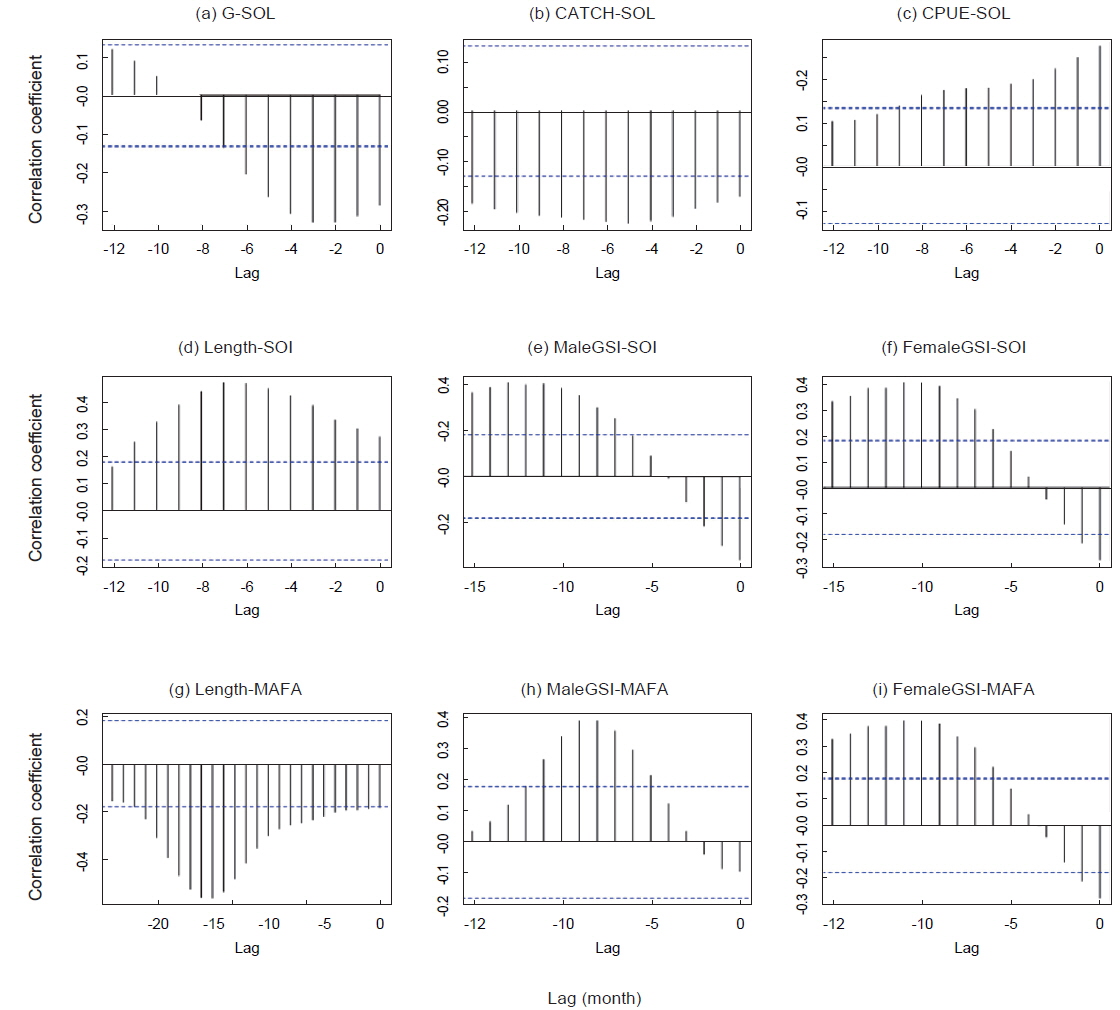
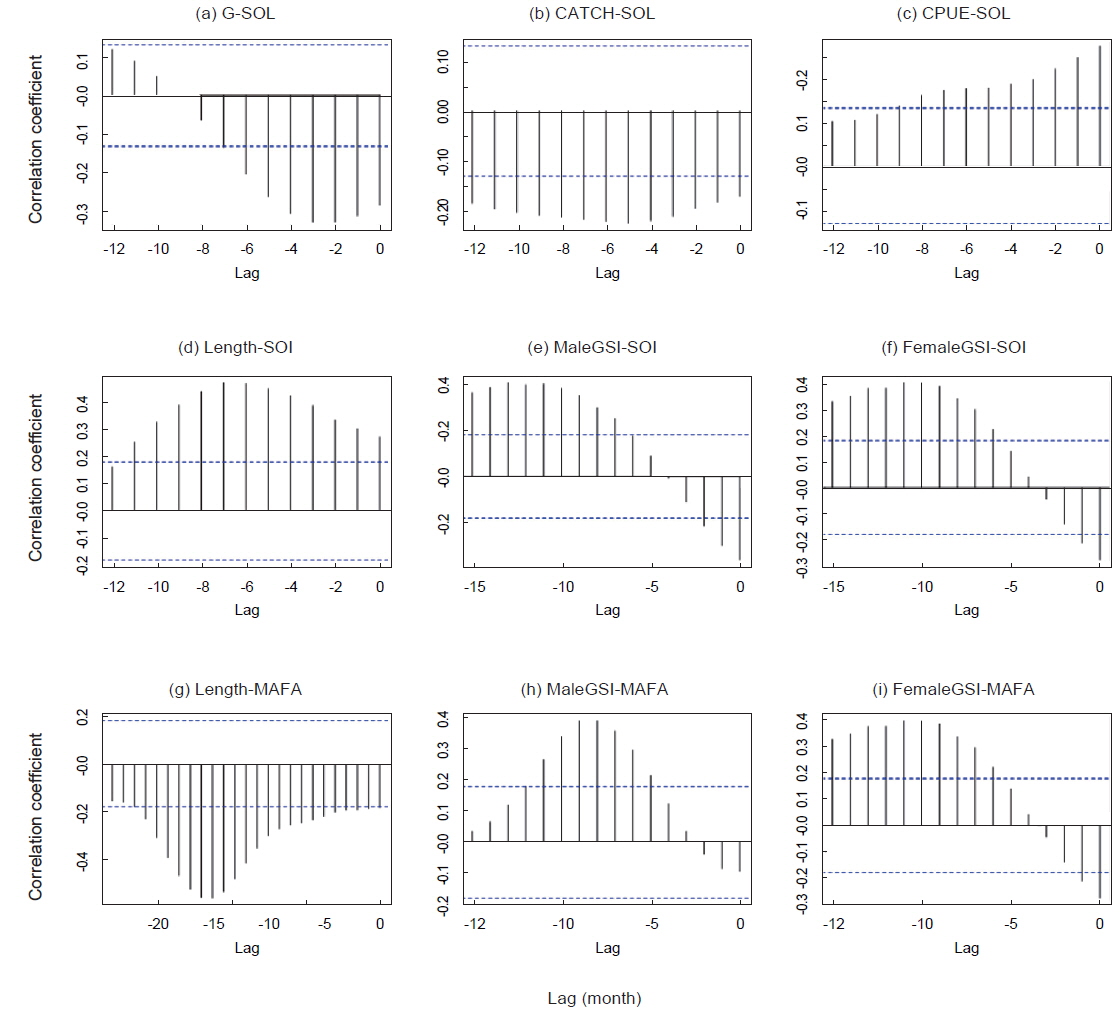
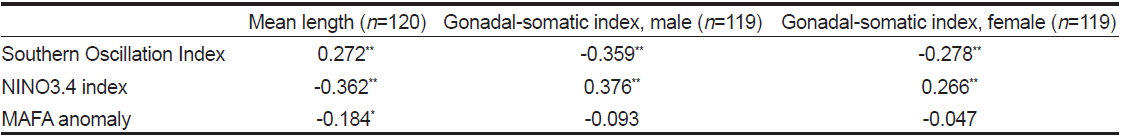
연구기간동안 가다랑어 단위노력당 생산량은 10-57톤이었다(Fig. 3a). 남방진동지수가 음의 값을 가지는 경우(1991/1994) 단위노력당 생산량은 점차 감소하였고 낮은 생산량을 보였다. 가장 낮은 단위노력당 생산량은 엘니뇨 현상이 가장 강했던 1997/1998년에 나타났다. 비록 어획량과 ENSO 지수와는 유의한 상관관계를 보이지 않았지만, 단위노력당 생산량은 모든 환경 지수와 유의한 상관관계를 보였다(Table2).
해양의 물리적 변화인 엘니뇨 현상은 시간 지연을 가지고 열대 태평양의 가다랑어 어업에 영향을 나타내었다. 어장위치의 변화인
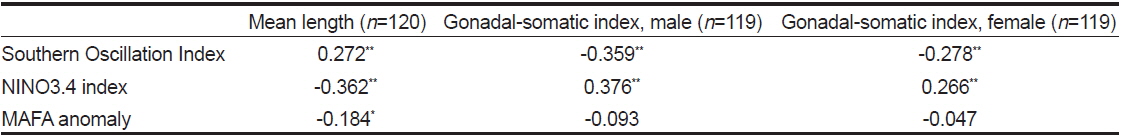
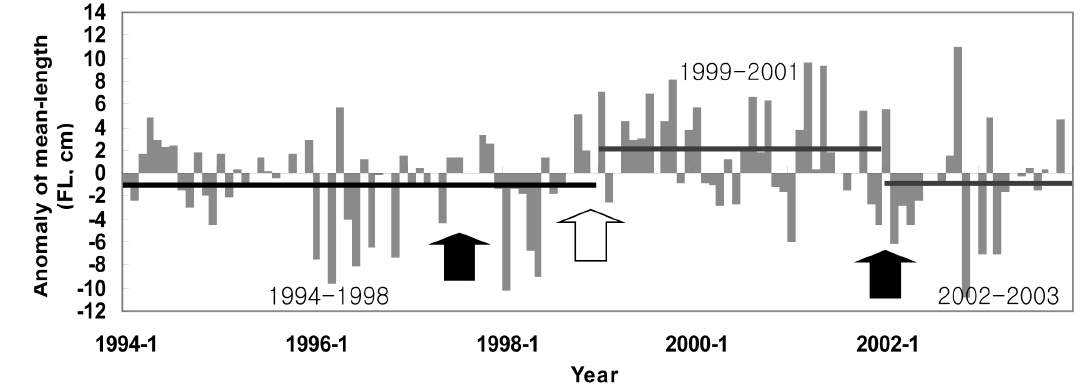
가다랑어 가랑이체장(length)과 생식소 중량지수(GSI)는 시간에 따라 변동하였지만 계절적 변화는 확인할 수 없었다(Fig. 3c). 하지만 이들의 변화는 ENSO와 연관성이 보였다. 1997/1998년의 강력한 엘니뇨 현상 이후, 2001년 후반기에 시작하여 2002년 전반기로 이어진 엘니뇨 기간동안에 평균체장은 감소하였다. 반면 라니냐 기간(1998/1999) 동안 평균체장은 증가하는 경향을 보인다. 엘니뇨가 가다랑어의 성장에 미치는 영향을 알기위해 두 현상의 상관관계를 알아보았다. 평균체장과 생식소 중량지수는 ENSO 지표와 유의한 상관관계를 보였다(Table 3). 남방진동지수와 NINO3.4 index는 평균체장과 양과 음의 상관관계를 보였으며, 이는 엘니뇨 현상이 나타날때, 즉 남방진동지수가 감소하고 NINO3.4 index가 증가할 때, 엘니뇨 현상이 나타나지 않을 때에 비하여 평균체장이 감소하였다. 그리고, 평균체장과 남방진동지수 사이의 교차상관관계는 7개월일 때 가장 높게 나타났다(r = 0.469,
Correlation coefficients between environmental indices and biological factors during 1994-2003
1994/1995년을 제외하고 생식소중량지수의 변화는 남방진동지수의 변화와 유사하다(Fig. 3c). 특히 1997/1998년 엘니뇨 현상 때 생식소중량지수가 가장 높았다. 남방진동지수와 생식소중량지수는 유의한 음의 상관관계를 나타냈으나(Table 3), 시간차이를 고려한 교차상관관계에서는 양의 상관관계를 나타냈다. 암수의 경우 모두 11-13개월의 시간차를 두고 가장 높은 상관관계를 보였다[r = 0.403(수) and r = 0.402(암),
MAFA anomaly는 생식소중량지수와는 유의한 상관관계가 없었으나 평균체장과는 음의 상관관계를 보였다(r = −0.184,
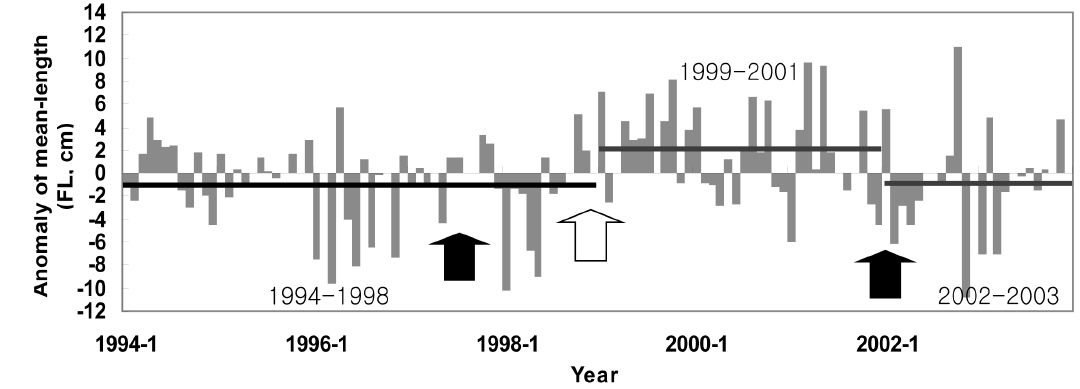
본 연구 기간동안 가다랑어 평균체장은 지속적으로 변동했으며(Fig. 5), 특정 기간 동안의 또렷한 증가 또는 감소를 보이고 있는데 그 시기는 1994-1998, 1999-2001, 그리고 2002-2003년이다. 이 기간 동안의 평균 아노말리는 각각 −0.99 cm, 2.09 cm, 그리고 −0.90 cm이다. 연구기간 동안 1991/1992, 1997/1998, 그리고 2002년에 엘니뇨, 그리고 1998/1999년에 라니냐 현상이 발생했다. 1999-2001년 동안 양의 아노말리를 보이던 체장은 2002년 엘니뇨를 지나며 음의 아노말리를 보였다. 반대로, 1994-1998년 동안 음의 아노말리를 보이던 체장은 1998년 라니냐를 겪으며 양의 값을 보였다. 특히 1998년 급격한 체장 감소는 가장 강력했던 1997/1998년의 엘니뇨 이후 발생했다.
1988년 태평양 섬 국가들의 배타적 경제수역의 발효로 우리나라를 비롯한 열대 태평양 선망어업국들의 어장이 제한되었다. 이러한 어장 제한으로 인한 영향을 최소화 하기 위해 Lehodey et al. (1997)는 어장에 제한이 없었던 미국 선망어업 자료만 이용해 가다랑어 자원의 동서 방향 분포 변화에 대하여 연구하였다. 우리나라 선망어업은 어장이 태평양 열대서부 해역에 치우쳐 있다는 한계를 가지고 있지만, 본 연구결과에서는 우리나라 어선의 가다랑어 어장도 ENSO의 영향을 받고 있는 것을 보여주었다. 우리나라 선망어선은 엘니뇨 현상이 나타난 뒤 2-3개월의 시간차를 두고 동쪽으로 이동했으며 이는 Lehodey et al. (1997)가 미국 선망어선 자료만 가지고 연구한 결과와 일치하였다.
열대태평양의 웜풀은 빈영양해역이지만 세계 최대 다랑어 어획지역으로 알려져 있다. 이는 웜풀의 가장자리에 형성되는 수렴대에 플랑크톤이 풍부하게 서식하기 때문이다(Yoder et al., 1994; Power, 1996). 아직 우리는 플랑크톤에서 가다랑어의 먹이생물이 되는 소형 어종이나 갑각류, 두족류까지의 연결되는 먹이망에 대해 잘 알지 못한다. 그러나, Lehodey et al. (1998)는 일차생산자인 식물플랑크톤에서 가다랑어의 먹이생물을 연결하는 생지화학모델을 통해 일차생산자의 분포가 웜풀과 적도 용승해역에서 높게 예측됨을 보여 주었다. 그리고 먹이생물은 적도해류의 수렴대에서 높게 예측되었으며 이는 가다랑어가 가장 많이 어획되는 곳과 일치했다.
열대태평양의 가다랑어 어장은 아주 넓기 때문에 어획효율을 높이기 위해서는 가다랑어의 분포편화를 예측하여야 한다. 본 연구의 결과는 ENSO가 가다랑어의 어획에 긍정적인 영향을 보이며, 또한 환경변화로 인한 어획량의 변화를 예측할 수 있음을 보여준다. 하지만, 본 연구에는 1985년부터 2003년까지의 어획량이 사용되었고 이 기간동안에 우라나라 선망어선의 수가 변화하였으며, 선망어획자료의 수집이 1997년 이후부터 50% 이하로 크게 감소하였다. 이로인해 어획량과 환경지수와의 상관관계는 정량화된 단위 노력당 어획량보다 유의하지 않게 나타난 것으로 생각된다(Table 2)
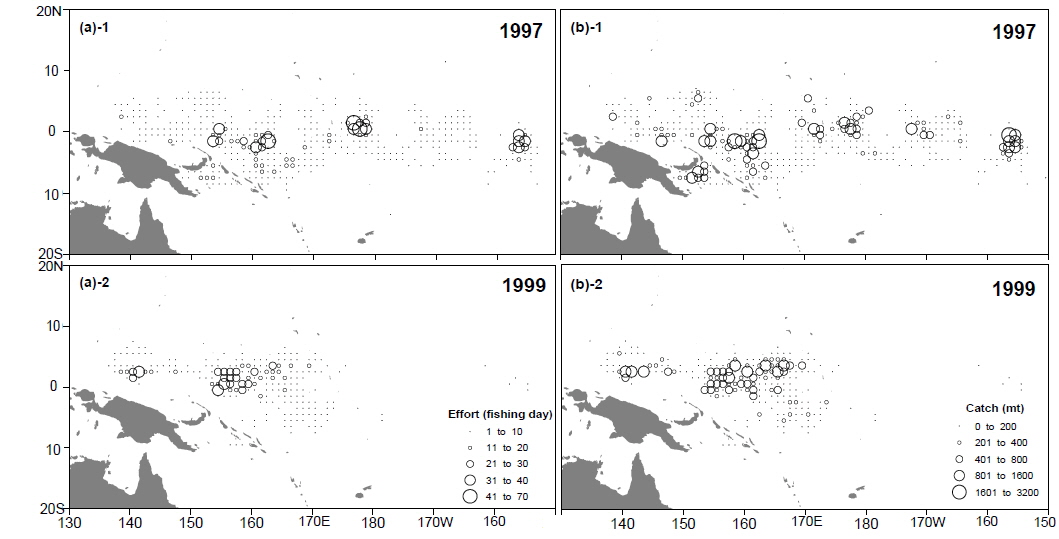
단위노력당 어획량은 엘니뇨기간 동안에 급격히 감소하였다. 가장 낮은 단위노력당어획량은 엘니뇨 현상이 가장 강했던 1997/1998년에 관찰되었다. 단위노력당 어획량과 남방진동지수는 시간차이 없이 가장 큰 상관관계를 보였으며, 이는 엘니뇨 기간 동안에 바로 어획효율이 감소되었음을 의미한다. 웜풀은 엘니뇨동안에는 동쪽으로 확장되며 라니냐 기간에는 서쪽으로 후퇴하는 경향을 나타낸다(Picaut et al., 1996; Matsuura and Iizuka, 2000). 이러한 웜풀의 확장과 수축의 변동으로 가다랑어 어장에서의 어획효율에 영향을 미치는 것으로 추측된다. 엘니뇨 기간 동안에는 웜풀와 적도해류의 사이에 형성된 수렴대가 웜풀의 확장으로 인해 동쪽으로 이동되며, 가다랑어의 먹이생물의 분포를 쫓아 동쪽으로 이동하는 것으로 알려져 있다(Lehodey et al., 1997; Lehodey et al., 1998; Lehodey, 2001). 그러나 우리나라 선망어업의 어획활동은 계속해서 웜풀 안에서만 이루어지고 있다. 강력한 엘니뇨가 일어났던 1997년에는 어획활동이 열대태평양 전역에서 활발하였으나, 1999년 라니냐의 경우 어획노력은 주어획해역(MAFA)안에서만 일어나는 것을 확인할 수 있다(Fig. 6). 그러므로 웜풀이 동쪽으로 확장하면서 가다랑어의 어획 해역도 넓어지는 것으로 보인다. 가다랑어의 분포수온은 20˚C에서 30˚C사이로 매우 넓은 편이다. 그러나 가다랑어의 분포 밀도가 높은 곳은 해면수온이 28˚C이상이 되는 열대 해역이다. 그러므로 웜풀의 확장은 가다랑어의 분포 밀도를 낮추어 단위노력당 어획량을 감소시키는 것으로 보인다. 1997년에 우리나라뿐만 아니라 다른 중서부태평양의 선망어업국의 단위노력당 어획량 감소는 이 가설을 뒷받침해준다(SCTB, 1998). 그러나 서부 열대 태평양의 해면수온과 어획량의 관계는 아직 불확실하다. 서부 열대태평양의 해면수온의 변화는 아주 미비하다. 엘니뇨 기간 동안 이 해역의 해면수온은 현저히 감소되는 것을 알 수 있다. 엘니뇨 기간 동안 어획 감소는 어획노력의 지리적 분산으로 인한 것으로 추측된다. 특히 어선의 이동은 이동 시간과 경비, 그리고 어획물의 변화를 감안하여 결정되는데, 모든 어선이 주 어획해역로부터 동쪽으로 이동하지 않는 것으로 보인다.
가다랑어의 평균 체장과 남방진동지수의 교차상관관계를 통해 엘니뇨가 평균체장을 줄인다는 가설을 고찰해 보았다. 가다랑어 자치어의 분포는 수온에 의해 결정된다(Forsbergh, 1989). 엘니뇨 발달로 인해 따뜻한 물의 분포가 확장되어 가다랑어 자치어에 적합한 환경이 조성된다. 또한 엘니뇨 기간 동안 웜풀과 콜드텅(무역풍에 의한 동부 태평양의 용승으로 인한 영양 염류가 풍부한 차가운 해류)의 경계가 사라지거나 약해지며(Vialard and Delecluse, 1998), 웜풀이 중부 태평양으로 확장되므로 서부태평양의 두꺼운 웜풀이 얕아져 일차생산력이 상승한다. 이로 인해 중서부 열대태평양에 서식하는 가다랑어 가입이 증가하는 것으로 보인다. 본 연구에서는 엘니뇨와 평균 체장의 변화와는 7개월의 시간차이가 나타났다. 다랑어 자원평가에 사용되는 통계모델인 MULTIFAN-CL의 결과를 보면 강력한 1997/1998년 엘니뇨 동안 가다랑어 가입이 높아질 것이라 추정했다(Hampton, 2002). 또한 1998년 후반기에 서부 태평양에서 많은 어린 개체의 가다랑어(20-35 cm)가 발견되었다(Lehodey et al., 2003). 엘니뇨 이후 높은 가입으로 인해 평균 체장은 감소하고 어획량은 증가한 것으로 보인다.
다른 많은 열대 어종과 마찬가지로 가다랑어는 지속적인 산란을 하는 것으로 알려져 있으며 체장이 40 cm 이상의 경우 성숙한 생식선이 연중 발견된다. Matsumoto et al. (1984)는 가다랑어 알의 지름 변화를 조사하여 이들이 연중 산란한다는 것을 밝혔다. Stequert and Ramcharrum (1996)의 연구에서는 70% 이 상의 암컷 가다랑어에서 최종 발달단계의 난소가 관찰되었으며, 생식소 중량지수(GSI)로도 연중 산란을 확인하였다. 본 연구에서도 GSI를 보면 특정한 산란 기간이 없이 변화하는 것을 확인할 수 있었다. GSI와 남방진동지수는 11-13개월 시차에서 가장 유의한 상관관계를 보인 것은 엘니뇨가 발생하고 약 1년이 지나면 가다랑어 생식소 발달에 부정적 영향이 나타난다는 것을 의미한다. 이 현상은 엘니뇨가 가다랑어 가입량을 높이는 것과 관련이 있는 것으로 보여진다. 가다랑어 가입의 증가로 종내 먹이경쟁이 치열해진다면 밀도 종속적 반응으로 생식소 발달에 영향을 줄 수 있다. 보통 가다랑어의 성성숙 체장이 40 cm (1세 이상, Matsumoto et al., 1984)인 것을 고려했을 때 엘니뇨가 GSI에 미치는 시간차가 11개월인 것은 이해될 수 있다. 하지만 본 연구에서는 엘니뇨가 일차 생산력에서 가다랑어의 생산력까지 미치는 메커니즘을 파악할 수는 없었으며, 높은 가입이 항상 높은 생산력으로 이어지지는 않았다. 가령, 1991/1992년 엘니뇨 이후의 어획량은 상승하지 않았다.
본 연구에서는 주어획해역(MAFA)의 해면수온은 가다랑어 평균체장과 생식소 중량지수와 유의한 상관관계를 보여주었다. 주어획해역(MAFA)에서 높은 수온은 9-11개월 뒤에 가다랑어의 성성숙도를 증가시켰다. Forsbergh (1989)는 수온이 1˚C 증가할 때 자치어의 밀도는 2 배로 증가한다고 서술한 바 있다. 만약 가다랑어가 높은 수온을 선호한다면, 주어획해역의 높은 수온이 가다랑어의 성숙도를 증가시켜 많은 산란을 유도하지만, 밀도종속적 반응으로 가입량은 오히려 감소될 수 있기 때문이다.
우리는 본 연구에서 우리나라 선망어업의 가다랑어 단위노력 당어획량(CPUE)을 중서부태평양의 가다랑어 분포의 표본으로 삼았다. 하지만 선망어업의 단위노력으로 사용된 조업일수는 어선 크기나 다른 사회경제학적 요소를 고려하지 않아 이상적인 단위노력의 기준이 되지 못한다고 생각한다. 그러나 우리나라 열대 태평양 선망어선들은 비슷한 크기(800-1,400톤)와 비슷한 장비를 보유하고 있으므로, 본 연구에서 사용한 우리나라 열대 태평양 선망어선의 가다랑어의 단위노력당어획량은 이 해역의 가다랑어 분포로 보기에 합당하다고 생각된다.
1997년 이후 선망어업 어장의 위치변화는 극대화되었다. 우리는 이 변화의 원인에 대해 두 가지로 생각해 보았다. 첫 번째는 1997/1998년도의 엘니뇨는 아주 강력했으며 따라서 가다랑어의 분포도 동쪽으로 극대화되어 나타났다. 두 번째는 1997년 이후 선망어획자료의 수집이 전체의 50% 이하로 감소되었다는 점이다. 표본 크기의 변화로 변동은 극대화 되어 나타났지만 변동의 양상은 앞선 연도와 동일하게 나타났다.