Among diverse freshwater ecosystems, riverine plains have distinctive landscape features and are characterized by high biodiversity and productivity and corresponding recreational and aesthetic values. Riverine plains are considered to be centers of biocomplexity, and more species of animals occur in riverine plains than in any other landscape type in most regions of the world, ranging from 12% of mammals to 60% of amphibians (Kelsey and West 1998). In tropical Asia, many mammals such as Malayan tapirs and Indian rhinos spend at least part of their life cycles in riverine wetlands (Dudgeon 2000). Furthermore, riverine plains support abundant vegetation that provides habitat for many non-mammalian species such as birds. For example, of all bird species breeding in northern Colorado, 82% occur in riparian vegetation, and about half of the species of the American Southwest depend upon riparian vegetation (Knopf and Samson 1994). Riparian areas also provide critical stopover areas for en-route migrants and therefore affect the breeding success of bird populations (Skagen et al. 1998). Diverse species of waterfowl (e.g., ducks, geese, and coots) use riverine habitats at certain stages in their life cycles, and riverine areas often support substantial numbers of wintering ducks, dabbling ducks, and diving ducks (Elkins and Lynch 1997, Krasnov et al. 2004). Patterns of habitat use by wintering waterfowl may provide insights into the habitat value of riverine areas. For example, if waterfowl are found to avoid using areas subject to indirect or nearby human disturbance, the value of that habitat for the conservation and management of wintering waterfowl populations may be reduced.
Riverine habitats are being increasingly impacted by both the direct loss and indirect effects of human activities within the habitat or the adjoining watershed (Mac et al. 1998, Kennish 2002). These effects are particularly acute in habitats that surround or are near urban centers. In order to adequately assess the costs and benefits of developing or protecting riverine lands, further information on the effects of human-induced alteration of these areas is required. Patterns of habitat use by wintering waterfowl may provide insights into the value of riverine habitats, particularly in heavily developed areas. To date, however, many habitat studies have mainly reported on habitats as refuges for animals avoiding predators and as protected conservation areas (Diehl 1992, Ward and Stanford 1993, Roux et al. 2008) and have not examined the effects of direct human disturbance on waterfowl. Riverine areas where human activity can frequently disturb waterfowl have been less studied, although they may be subject to greater pressures from human-induced disturbance. Even though a habitat itself may remain intact and protected, its landscape setting can be altered by development or human activity within its watershed.
The landscape setting of a habitat has been shown to strongly influence waterfowl distribution and species composition. Some studies have reported that waterfowl assemblages were clearly affected by habitat change resulting from changes in characteristics such as landscape heterogeneity (Roe and Georges 2007), the extent of urbanization or agriculture (Brouder and Hill 1995, Crooks et al. 2004), and edge density (Pasitschniak-Arts and Messier 1995). In this regard, the quantitative assessment of habitat can be used as an analytical tool to evaluate habitat characteristics for sustaining waterfowl populations. Habitat assessment of individual wetlands provides objective criteria for habitat protection and reinforces bioconservation strategies in establishing a natural sanctuary. Some researchers have suggested that certain structural or habitat characteristics are important in determining habitat use (Esselink et al. 2000, Adamik et al. 2003). For waterfowl, important habitat characteristics may include protection from exposure to harsh winds, cold, extent of the shoreline, the presence of adequate food, and size of habitat area (Christiansen and Low 1970, Sanderson 1980).
In this study, we examined waterfowl wintering in newly created artificial wetlands in a riverine area, the Nakdong River Basin, South Korea. We compared numbers of individuals and species of waterfowl in these created wetlands along a gradient of human-induced disturbance. We hypothesized that the excessive use of artificial materials when new wetlands are constructed negatively influences wintering waterfowl. Based on habitat assessment results at each created wetland, we discuss the ecological relevance of influences on riverine habitat use by wintering waterfowl.
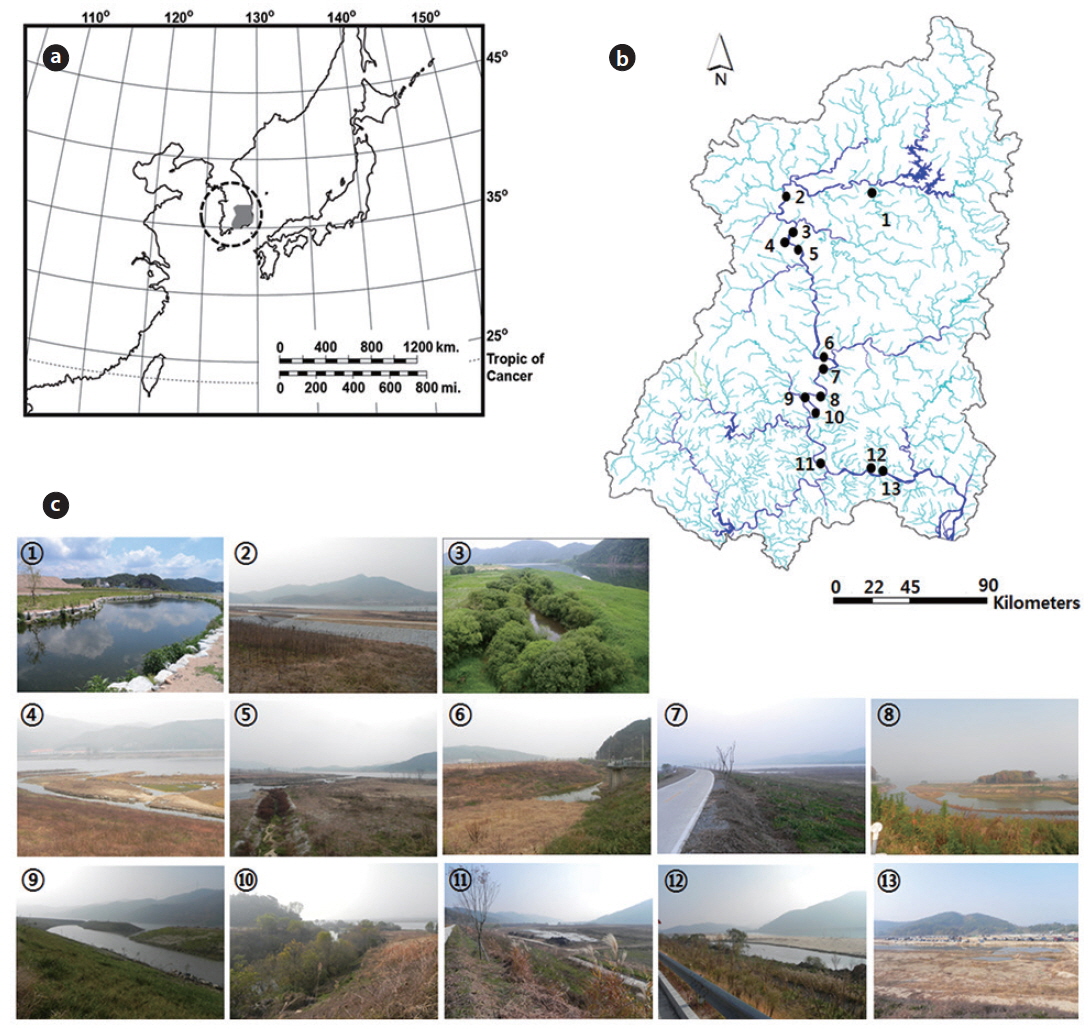
South Korea has a temperate climate with four distinct seasons. The annual average rainfall is approximately 1150 mm; freshwater ecosystems in this country have maximum rainfall in the summer, with more than 60% of annual rainfall occurring from June to early September (Jeong et al. 2007). The wetland study sites monitored in this study are located in the middle and lower reaches of the Nakdong River. The Nakdong River Basin was heavily modified by the Four Large River Projects (4LRP), which in 2011 built eight large weirs across the river to maintain a minimum channel depth and facilitate water supply. Furthermore, most riversides (riverine areas) have been changed by human-induced alterations (i.e., the construction of parks, parking lots, and other amenities). Historically, numerous riverine wetlands have dominated the river basin, but human impacts have led to modifications or losses in their morphology and habitat structure. Currently, most of the riverine wetlands along the Nakdong River were created or restored by humans.
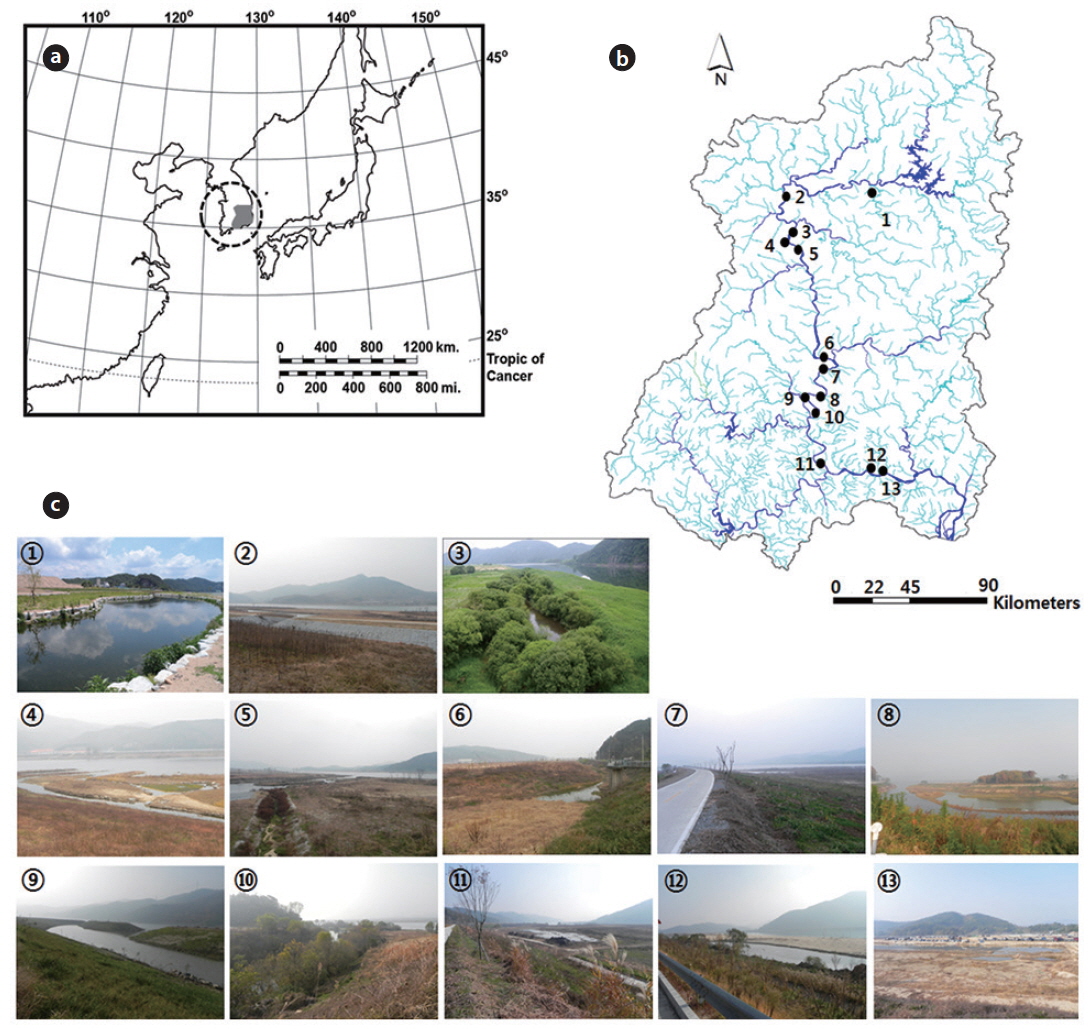
The river basin has a total of 57 created wetlands, of which this study examined 13. We used two criteria to select habitats for evaluating conditions that sustain waterfowl populations: (i) waterfowl should have been observed in the study site or surrounding area in previous studies; and (ii) the environmental variables of the study sites should be broadly similar. Consequently, we selected 13 wetlands in the river basin for analysis (Fig. 1). Six aquatic macrophyte species were commonly observed at the study sites:
We surveyed the wetlands during the winter (November 2012 to January 2013). Waterfowl were investigated using a 15-45 x 60 spotting scope or 10 × 50 binoculars. Each wetland was surveyed three times during the survey period (for a total of three samples per wetland over the course of the study) on randomly chosen days and at randomly chosen times of day. Each bird census was conducted using the point count method (Bibby et al. 2000). At each wetland, the census was conducted from a position that enabled observation of the entire surface of the site from the land. Waterfowl monitoring was performed in weather conditions calm enough for birds to be active and detected by sight and sound. The numbers and species of waterfowl present at the sites were recorded during each survey. Waterfowl were classified into six ecological numtrait groups according to species characteristics, including food sources (e.g., insects, fish, invertebrates, fruit, and seeds), foraging style (e.g., diving and hovering), habitat preference (e.g., mud flat, water surface, or tree), and morphological characteristics (e.g., taxonomic array and keys) (Weller 1999, Lee et al. 2000). Accordingly, the classification groups of ecological traits were grebes, egrets and herons, geese, dabbling ducks, diving ducks, and coots.
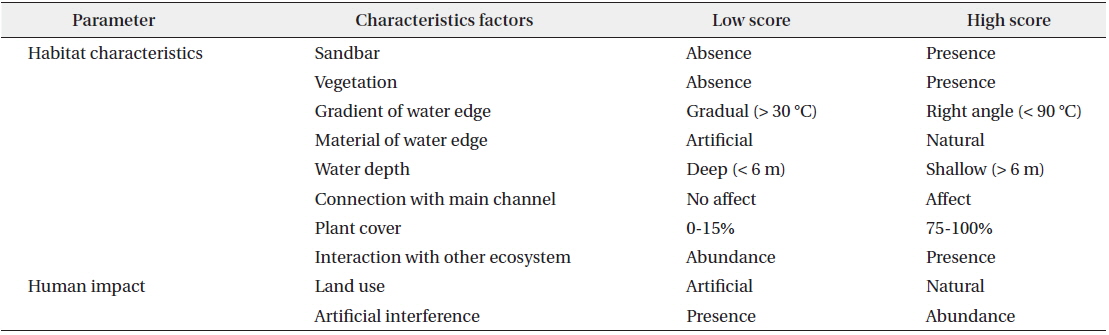
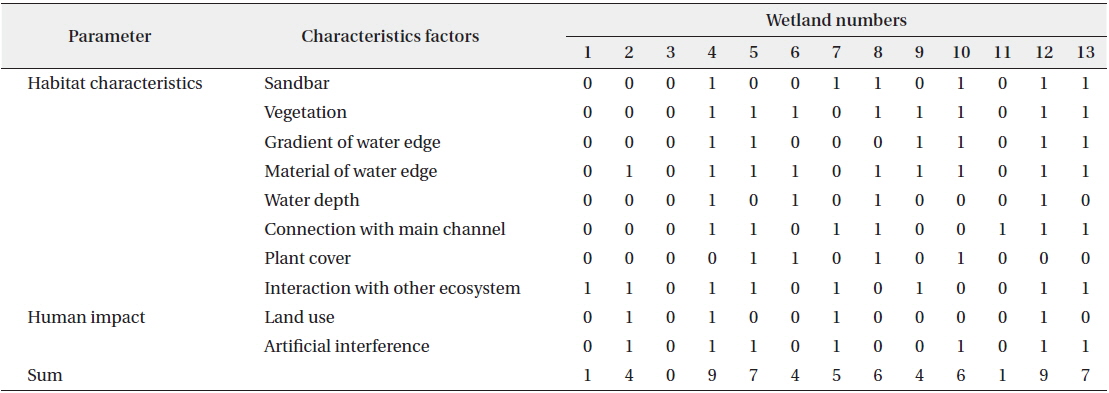
A total of 10 presence–absence type (dichotomous) survey parameters evaluating habitat conditions for sustaining waterfowl were used in field monitoring (Table 1). These parameters were subsequently combined to form two factors, habitat characteristics and human impact, for all 13 created wetlands in this study. Habitat suitability at each site was measured by scoring each of 10 parameters that we developed based on literature reviews on waterfowl habitat (Briggs and Maher 1985, Smith et al. 1989, McKinney et al. 2006), discussion with a number of freshwater ecologists, and an on-going Rapid Assessment Method (RAM) in Korea used to evaluate site information and wetland habitat conditions as described in a report of the Korea Ministry of Environment (2008). The 10 parameters for rating habitat suitability were sandbars, vegetation, gradient of the shoreline, material at the shoreline, water depth, connection with the main channel, plant cover, interaction with other ecosystems, land use, and artificial interference. In order to minimize bias in observations, a single scientist specializing in freshwater ecology scored the habitat suitability parameters at every study site. Monitoring was conducted at the center point of study sites where the overall characteristics of a wetland could be observed.
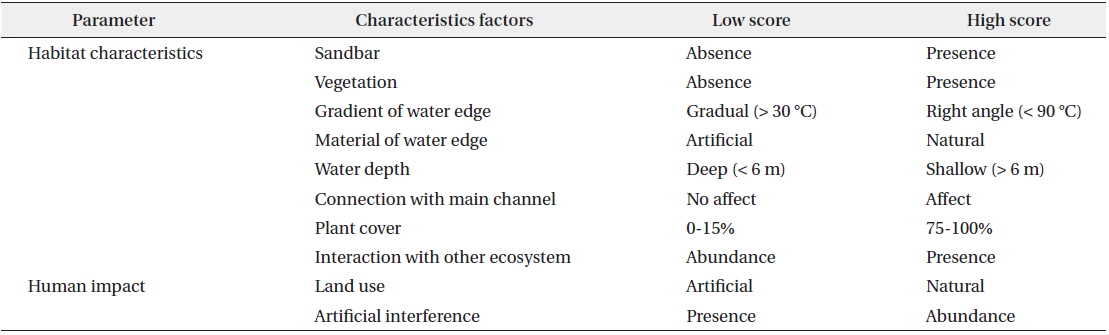
Habitat assessment index used in field monitoring for assessment waterfowl habitat condition
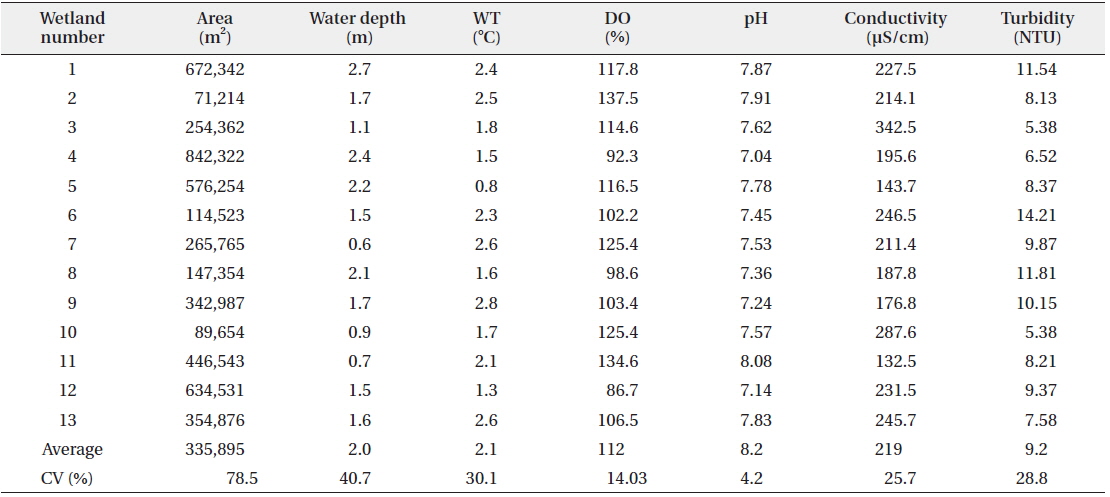
Measurements of water temperature, dissolved oxygen, pH, conductivity, and turbidity were performed at each of the wetlands included in the waterfowl survey. Water samples were collected at a depth of approximately 0.5 m. We used a dissolved oxygen (DO) meter (Model YSI 58; YSI Research Inc., Yellow Springs, OH, USA) to measure water temperature and dissolved oxygen pH and conductivity were measured using a pH meter (Orion Model 250A; Orion Research Inc., Boston, MA, USA) and conductivity meter (Model YSI 152; YSI, Yellow Springs, OH, USA), respectively. Turbidity was measured using a turbidimeter (Model 100B; HF Scientific Inc., Ft. Myers, FL, USA). The area of open water was measured using a Geographic Information Systems (GIS) program (ArcGIS 9.3; ESRI, Readlands, CA, USA) and a digital map (National Geographic Information Institute 2005; scale 1:25,000).
The relationships among numbers of individuals and number of waterfowl species, habitat assessment scores, and environmental factors were tested with stepwise multiple regression. To understand the influence of the habitat assessment score on numbers of individuals and number of waterfowl species, we used regression analysis. We tested for linear, exponential, inverse, power, and logistic functions to determine the equation with the best curve fit. Among the regression results, the curve-fitting equation that returned the highest determination coefficient was selected to explain the observed relationships. All statistical analyses, including stepwise multiple regression and regression analysis, were conducted using the statistical package SPSS ver. 14 for Windows (SPSS Inc., Chicago, IL, USA).
The observed levels for environmental factors (area, water depth, water temperature, dissolved oxygen, pH, conductivity, and turbidity) at the study sites are listed in Table 2. Even though some wetlands showed exceptionally high or low relative values for these factors, the coefficients of variation (CV) were less than 100%. Among environmental factors, the area of the wetland had the highest CV, but the variation was only approximately 78.5%.
[Table 2.] Environmental characteristics measured at study sites
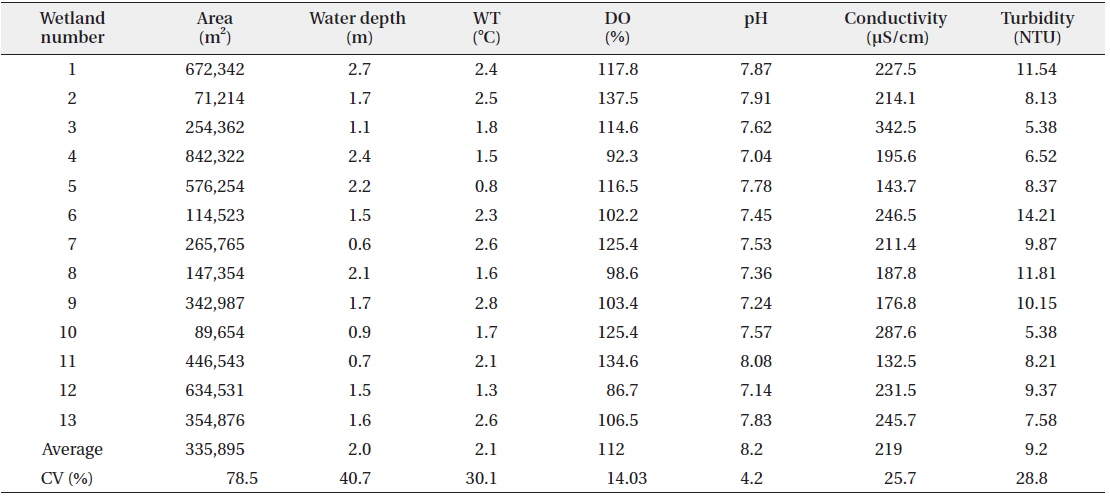
Environmental characteristics measured at study sites
Habitat assessment scores were clearly different among the study sites (Table 3). The highest scores were found at Sites 4 and 12, followed by Sites 5 and 13, while Sites 1, 3, and 11 had lower habitat assessment scores. Most of the created wetlands had natural material (e.g., soil and natural stone) and vegetation along the shoreline and were adjacent to other ecosystems (eight to nine sites). The study sites were characterized by relatively deep water, a low percentage of plant cover over the water’s surface, and artificial land use (i.e., parks, parking lots, and amenities) near the site (approximately within 5 m). These habitat characteristics strongly influenced the sustainability of waterfowl populations during winter.
[Table 3.] Habitat assessment scores of field monitoring in study sites
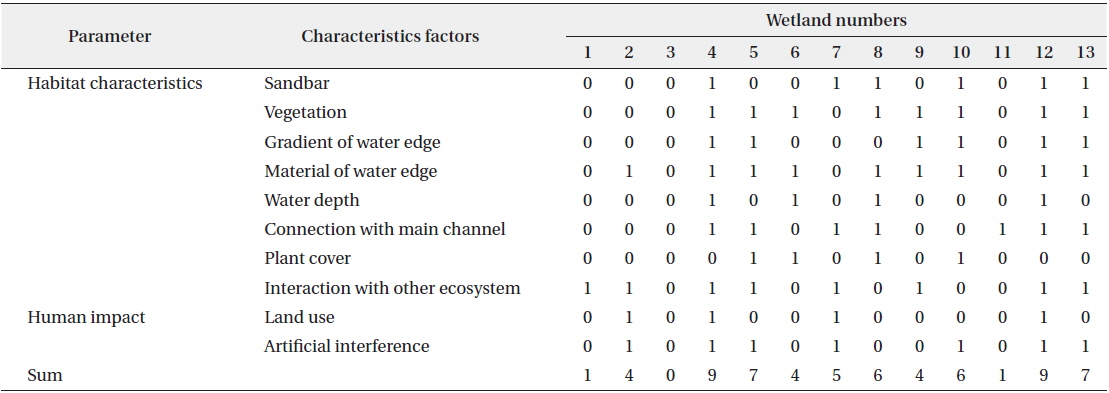
Habitat assessment scores of field monitoring in study sites
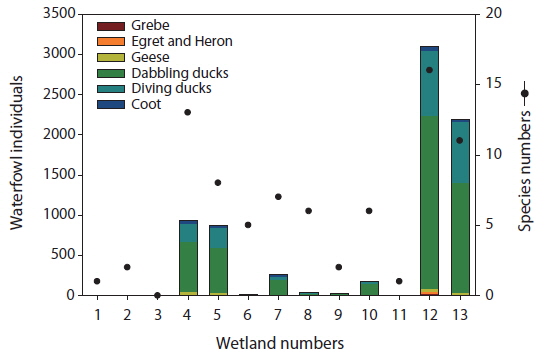
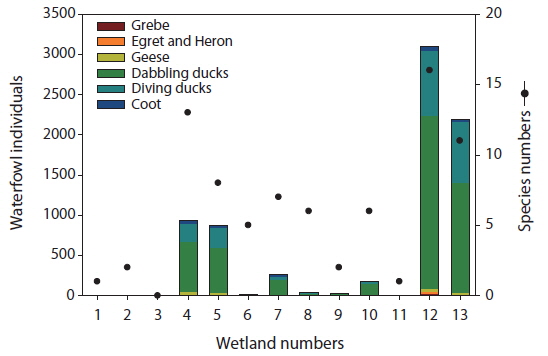
The numbers of individuals and species of waterfowl also differed among the study sites (Fig. 2). We identified a total of 22 species among six groups of waterfowl (grebes, egrets and herons, geese, dabbling ducks, diving ducks, and coots) in the 13 created wetlands. Among the study sites, Sites 4, 5, 12, and 13 supported over 500 individual waterfowl. The greatest number of individuals (3,104) was observed at Site 12, followed by Site 13 (2,197). The number of species of waterfowl in general corresponded to the number of individuals. However, Sites 6 and 8 had relatively many species despite their low number of individuals. Furthermore, Site 4 had fewer individuals than Site 13 but more species. Among waterfowl groups, dabbling ducks were the most abundant in all study sites, followed by diving ducks. In particularly, the sites with over 500 individuals also had more diving ducks than other sites as well as geese. We did not count the number of individual waterfowl at Site 3.
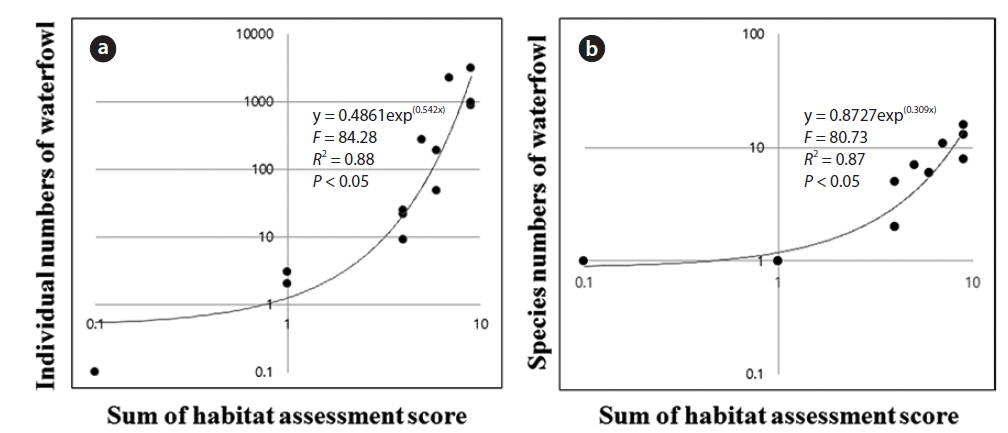
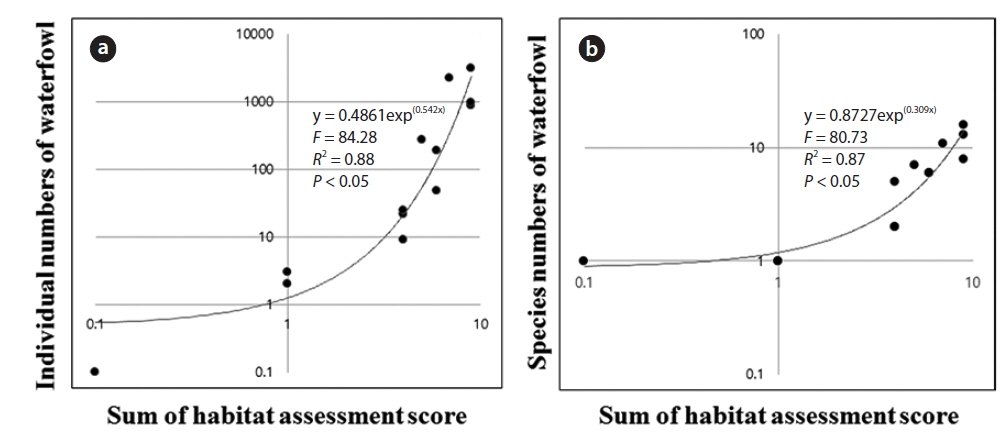
The results of the stepwise multiple regression allowed us to examine the relationship of environmental factors and habitat assessment scores to the distribution and assemblage of waterfowl (Table 4). The number of species had a strong relationship with the sum of the habitat assessment scores (

Summary of stepwise multiple regression aimed to predict density of species number and individual of waterfowl (response variable) with respect to habitat characteristics (environmental factors and habitat assessment score; explanatory variables) in study sites
Regression analysis (Fig. 3) clearly showed a relationship between the sum of habitat assessment scores and the number of individuals. An exponential function generated the highest determination coefficient. The number of individuals was positively correlated with the sum of the habitat assessment scores (Fig. 3a). The number of species also had a significant relationship with the sum of the habitat assessment scores (Fig. 3b).
In this study, the numbers of individuals and species of waterfowl were greater in created wetlands with high habitat assessment scores than low habitat assessment. A positive relationship between waterfowl numbers and the habitat assessment score was demonstrated through regression analysis. However, the absence of certain habitat characteristics led to low assemblages of waterfowl during winter. These differences may, to some extent, be attributed to human-induced disturbance of the riverine landscape setting of Nakdong River Basin; however, there may also be differences due to unrelated seasonal and yearly changes in the number of individuals. Most of the created wetlands were not intended to function as habitats for diverse animals, including waterfowl, but instead were constructed for anthropocentric reasons (Richardson 1994). Moreover, the surrounding land use is important for waterfowl habitat, but currently the areas surrounding the wetlands have artificial land uses or habitats with low heterogeneity. Natural habitats with woods and plants tend to be better protected from prevailing winter winds and from wave action because of the buffering effect of neighboring land, and waterfowl may seek better-protected sites to minimize the energy costs of thermoregulation (Bennett and Bolen 1978, Jorde et al. 1984). Among the study sites, only four created wetlands had a natural surrounding landscape (Sites 1, 8, 11, and 12), and most created wetlands had artificial surrounding land uses. In particular, Sites 8 and 11 supported relatively many individuals. However, even if the surrounding landscape was artificial, some created wetlands that had no human interference still had many individuals and species (e.g., Sites 6, 7, and 10). Therefore, the utilization of riverine areas by humans disrupts the distribution of waterfowl during winter.
Fig. 4 shows a panoramic view of some created wetlands (Sites 4 and 13). Most of the created wetlands constructed in riverine areas of the Nakdong River Basin by 4LRP had a low degree of naturalness and/or low structural heterogeneity of the surrounding landscape, resulting in areas that were not suitable for supporting a large assemblage of waterfowl. A low degree of naturalness of the landscape surrounding wetlands inhibits mutual connection with other habitats and leads to habitat fragmentation. Because of the importance of nesting success to waterfowl population dynamics (Sæther and Bakke 2000), many empirical studies have attempted to address the effects of habitat fragmentation on nest survival (Robinson et al. 1995, Donovan et al. 1997). The fragmentation of waterfowl wintering and breeding habitats has been extensive and continues to occur at a rapid rate in many countries, with negative demographic implications (Askins 2000, Boulinier et al. 2001). Habitat fragmentation typically reduces total habitat area and the size and proximity of individual habitat patches and can increase the amount of edge habitat. A reduction in the area of suitable habitat can result in waterfowl population declines simply by reducing the amount of adequate space for territories, nest sites, and other critical resources (Rolstad 1991).
The internal characteristics of created wetlands and their surrounding landscape are important to sustaining waterfowl populations. In this study in particular, not only the presence of sandbars and vegetation but also a low shoreline gradient and the use of natural materials along the shoreline led to a high abundance of waterfowl during winter. Waterfowl are ecologically specialized with respect to food and habitat use, with distinct species assemblages on different wetland types (Kantrud and Stewart 1977, Kauppinen 1995). Kauppinen (1995) reported that variations in waterfowl community structure were explained by the quantity of emergent vegetation and the size and depth of wetlands. Moreover, research conducted by Losito and Baldassarre (1995) found a positive correlation between emergent vegetation in wetland basins and the number of waterfowl. Waterfowl did not prefer wetlands with low structural heterogeneity around the shoreline and in the surrounding landscape (Bowman and Harris 1980). In particular, the shoreline areas of some created wetlands were specially constructed for easy human access, and this human-oriented construction limited the distribution of wintering waterfowl. Furthermore, interaction with other ecosystems was also an important factor in sustaining waterfowl during winter. Murphy et al. (1984) found that ponds in eastern Alaska that were hydrologically connected to a river ecosystem had more waterfowl and higher levels of most nutrients and ions than isolated wetlands. The input of nutrients in wetlands was also shown to affect the diversity and the total number of waterfowl (Hahn et al. 2008).
Nevertheless, artificially created wetlands in the study area were used by waterbirds for overwintering. As with other man-made wetlands (Maeda 2001, Tourenq et al. 2001, Ma et al. 2004, Paracuellos and Telleria 2004, Santoul et al. 2004), these wetlands were exploited throughout the year by several waterfowl species and provided them with a refuge, food, and/or breeding sites. In general, artificial wetlands have less abundant populations than natural wetlands (e.g., Tourenq et al. 2001, Ma et al. 2004), but these man-made wetlands may be important in some situations and for certain species (Froneman et al. 2001, Elphick and Oring 2003). For example, irrigation ponds support larger populations of black-winged stilts, little grebes, and common shelducks than natural wetlands in the area (Martí and Del Moral 2003).
Dabbling ducks, which feed on invertebrates in the summer, can feed on foliage in the winter. These ducks were more sensitive to habitat changes. In addition, passerines mirrored the changes in land-use patterns for surrounding the wetlands. In this study, the presence or absence of littoral vegetation was also influential in determining changes in species richness. Larger wetlands, which have a greater variety in habitat and cover, support more diverse and larger waterfowl communities. Restored wetlands are typically size-restricted because of constraints on available land area (Paracuellos 2008). To improve the resilience of restored wetlands, it is therefore necessary to increase the connectivity between created wetlands and adjacent rivers, which creates greater diversity. Increasing this connectivity concordantly increases the types of habitat patches available for the waterfowl community to colonize and use. This in turn supports greater species diversity within the restored wetlands.
In this study, we found that the numbers of species and individuals of waterfowl were clearly influenced by the habitat characteristics of created wetlands. In particular, not only the presence of habitat characteristics such as macrophytes and sandbars, but also the surrounding land-use patterns were recognized as important factors for sustaining waterfowl assemblages in created wetlands. These findings imply that increasing the naturalness of surrounding land and the wetland itself would increase species diversity and abundance. Many studies have shown that increasingly complex land-use patterns have positive impacts on avian diversity because of the relationships between diversity and complexity (McKinney et al. 2011, Harrison and Whitehouse 2012). The different habitat requirements of the waterfowl community have led to different waterfowl assemblages among created wetlands. Consequently, we suggest that the anthropocentric construction of created wetlands decreases the diversity and abundance of waterfowl, which may lead to less biodiversity in the food web structure of wetlands. In this study, habitat assessment was used as an analytical tool to predict the assemblage of waterfowl in restored and created wetlands. Further, complex habitats, such as wetlands and some terrestrial habitats, support both aquatic and terrestrial species because mixed habitats feature a larger array of food sources than more limited habitats do.