In the deciduous temperate forests of North America, spring ephemerals represent a common component of the herb layer (Schemske et al. 1978, Motten 1986, Lubbers and Lechowicz 1989, Kudo and Maeda 1998). For the spring ephemeral plants, the period of active growth corresponds with high light period, water, and nutrient availability between snow melt and canopy closure in the understory of the deciduous forests in eastern North America (McKenna and Houle 2000). The life of spring ephemerals is characterized by early appearance soon after snowmelt, prompt flowering, and short growing season within the high-irradiance period before canopy closure (Lapointe 2001). However, most spring ephemerals are potentially long-lived herbaceous perennials and the total biomass increases with increasing light intensity (Struik 1965, Tamm 1972, McKenna and Houle 2000, Rothstein and Zak 2001, Whigham 2004, Horibata et al. 2007). They exhibit a typical ‘sun-plant’ photosynthetic property with high light-saturated rate of photosynthesis and light compensation points (Rothstein and Zak 2001). Previous studies of the spring ephemerals were focused on energy budget, flowering strategy, and adaptation to low air temperature. The flowering strategy is composed of time and duration of flowering. These parameters can be assessed on four ecological levels: a flower, a plant, a population, and a species. However, much data on the ephemeral plant are required on all levels to understand its strategy comprehensively. Flowering time is important to pollinator limitation, herbivore’s activity, and survival in abrupt low temperatures. If a plant flowers in the early spring time, more pollinators can visit and more seeds can be produced by long fruiting period. However, the low air temperatures in early spring slow down the metabolism, restrict cell division and elongation, and thus reduce the growth rate of the plant (Ting 1982). Moreover, cool and unsettled weather conditions in early spring may reduce pollinator availability and cause deterioration in seed-set due to the pollen restriction (Helenurm and Barrett 1987, Nishikawa and Kudo 1995). It was shown that hand pollination produced more seeds than natural one (Schemske et al. 1978). Fortunately,
Flowering duration was responsive to pollinator activity, intensity of competition for pollinator acquisition, and climatic condition (Zimmerman 1988, Helenurm and Barrett 1987). In other words, long flowering duration is beneficial for pollination, but disadvantageous for feeding by herbivorous animal. Moreover, short fruiting periods result in the production of fewer seeds compared to longer fruiting periods. However, strategy for the flowering duration in spring ephemerals is diverse within the plant’s microhabitat in a population (Horibata et al. 2007) as a result of their very short life cycle of 40 to 60 days (Lapointe 2001).The spring ephemerals were rich in Korea (Cho 1998). One of the spring ephemerals,
The aim of this study was to examine the factors that influence flowering time in a natural
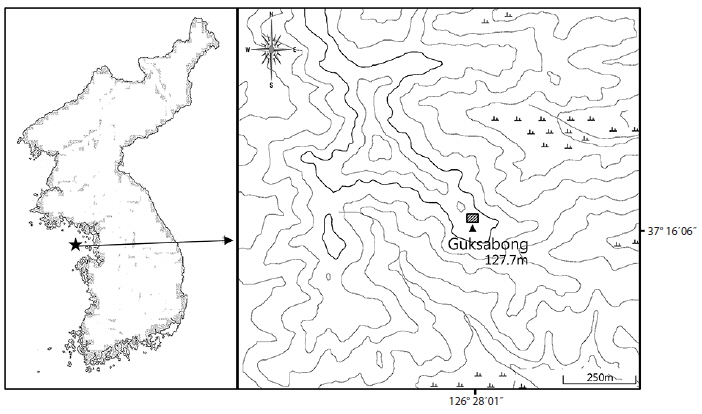
The location of the study area was at Nae-ri, Yeongheung- myen, Ongjin-gun, Incheon-city (37°16′14.8″ N, 126°27′49.9″ E) (Fig. 1). The altitude of this study site was 120.7 m above sea level (a. s. l.), with 15° slope and 30° in direction. Vegetation was composed of three layers: tree layer, shrub layer, and herb layer. The tree layer was composed of
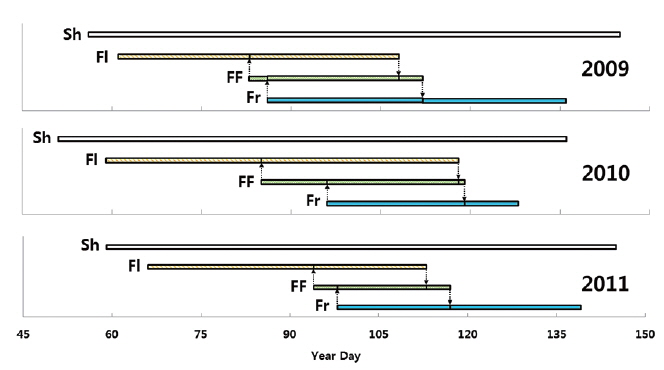
The permanent quadrat (50 m × 5 m) was located. By using numbered formax bars, 100 plants were individually marked on March 1, 2009. Formax rods were set in the soil near the each plant. Thirty five plants in the quadrat were additionally numbered on February 26, 2010. Reproductive organ of
[Table 1.] Phenological stages of sexual organ of Adonis multiflora

Phenological stages of sexual organ of Adonis multiflora
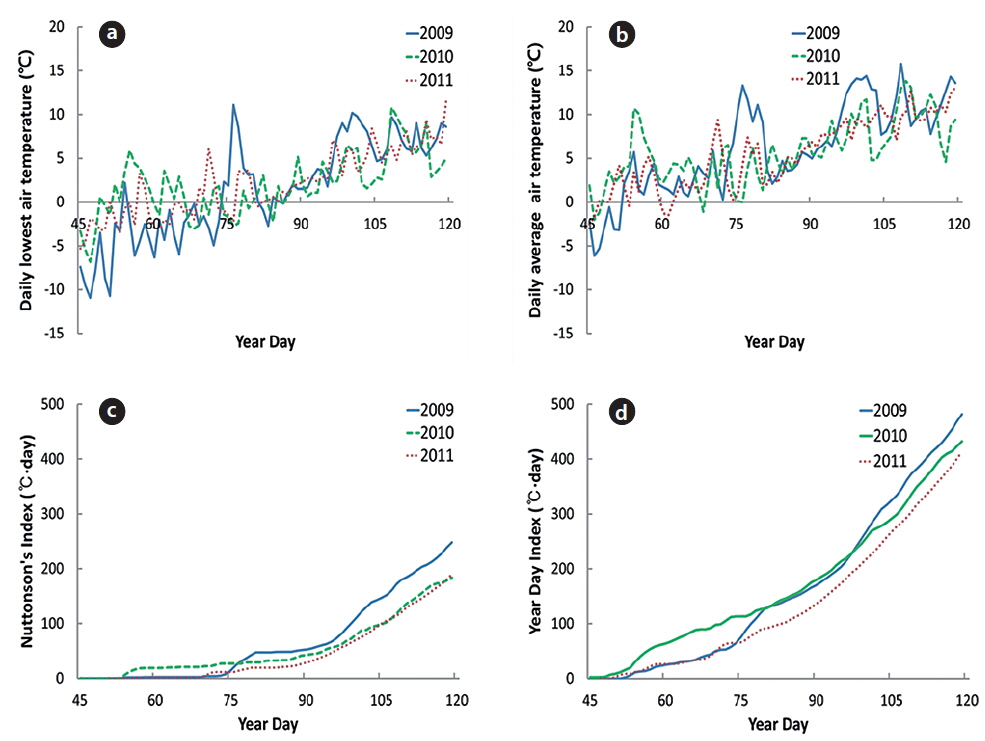
To measure the microclimate, automatic TRIX-8 thermometer (LogTag Recorders, Auckland, New Zealand) was set up at 4 m height from the soil surface on January 1 and collected on May 30 each year from 2009 to 2011. This thermometer was set up to measure air temperature by 0.1℃ in a two-hour interval. Daily mean air temperature, daily lowest air temperature, year day index (YDI), and Nuttonson’s index (Tn) were calculated from the field data. Tn was calculated by 4℃ instead of 5℃ (Yim et al. 1983, Min et al. 2007). YDI was the cumulative temperature over 0℃ and Tn over 5℃ in daily mean air temperature. Year day (YD) refers to the Julian calendar. The fact that was highly correlated with the YD meant nothing to do with daily mean air temperature. Correlation coefficients between phenological events and factors related to temperature were calculated by
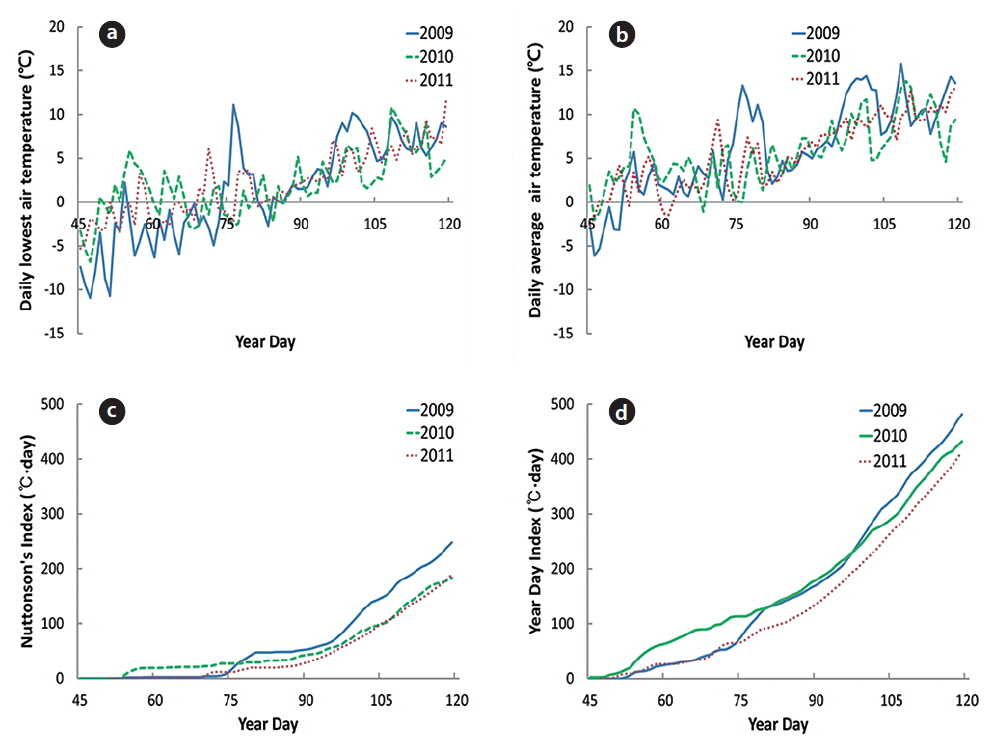
In 2009, the daily lowest air temperature (LT) was maintained below 0℃ until March 15 (YD 74), except for February 24 (YD 55); LT was also below 0℃ from March 24 (YD 83) to March 28 (YD 87). In 2010 and 2011, LT fluctuated above and below 0℃ from February 15 (YD 46) to March 28, and thereafter maintained above 0℃. The lowest air temperature was -6.3℃. Thus, plants experienced a sub-zero air temperature in March of each year (Fig. 2a).
In 2009, daily mean air temperature (MT) was maintained above 0℃ from February 22 (YD 53) and temporarily above 10℃ on March 17 (YD 76); thereafter, MT was maintained below 10℃ from March 22 (YD 81) to April 6 (YD 96) (Fig. 2b). In 2010, MT was maintained above 0℃ from February 18 (YD 49), except for March 10 (YD 69). In 2011, MT was maintained above 0℃ from February 17 (YD 48), except for March 3 (YD 62). The plant growth might have been retarded in mid-March in 2010 as following section.
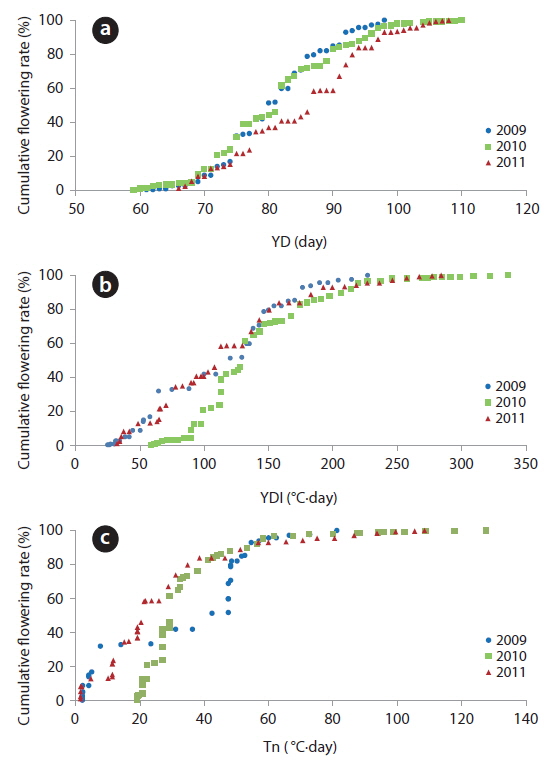
The values of year day index (YDI, ℃·day) were 50 on March 12 (YD 71), 100 on March 20 and 150 on March 29 (YD 88) in 2009. On February 27 (YD 50), March 14 (YD 73), and March 27 (YD 86) in 2010, YDI (℃·day) reached 50, 100, and 150, respectively; and the same pattern repeated on March 13, March 26, and April 3 in 2011. Thus, cumulative temperature-day of early-March was the highest in 2010. However, the date that YDI reached 150℃·day was similar for all three years.
Nuttonson’s index (Tn, ℃·day) reached 30 on March 19 (YD 78), 50 on March 30 (YD 89), and 100 on April 10 (YD 100) in 2009. The dates that Tn (℃·day) reached 30, 50, and 100 in 2010 were March 24 (YD 55), April 5 (YD 95), and April 16 (YD 106), and the same pattern repeated on April 2 (YD 92), April 7 (YD 97), and April 17 (YD 107), in 2011. Thus, Tn in March increased faster in 2009 than in 2010 or 2011.
>
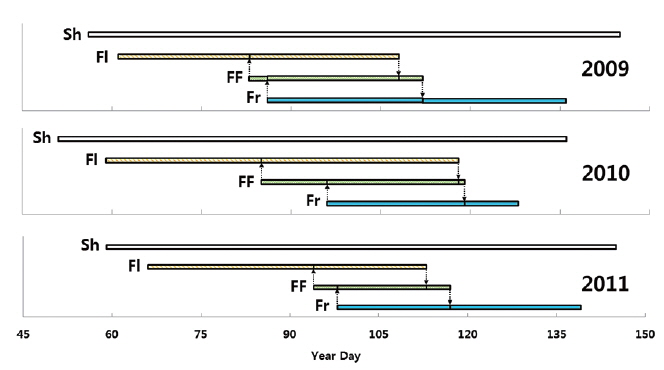
Life cycle of adonis multiflora during growth season
A shoot of
>
Flowering and daily mean air temperature
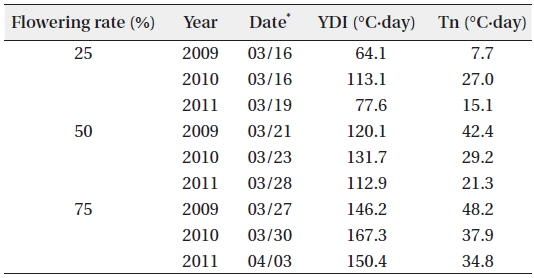
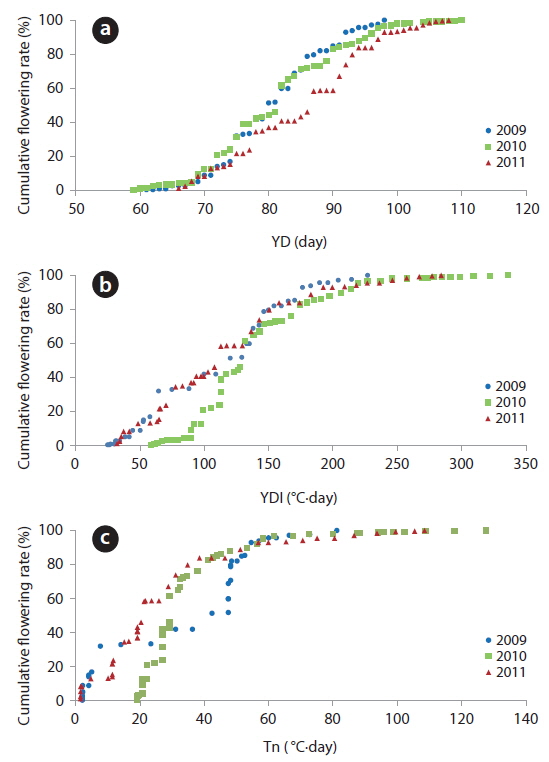
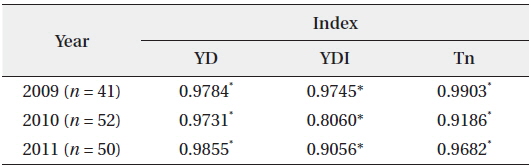
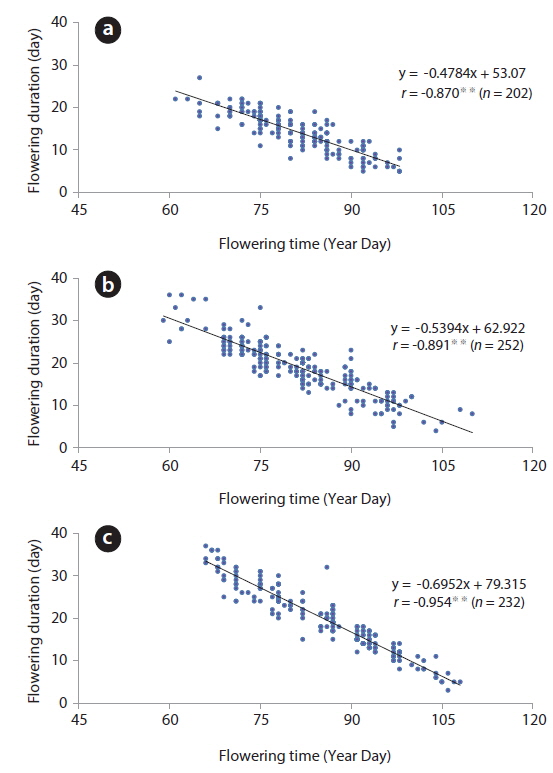
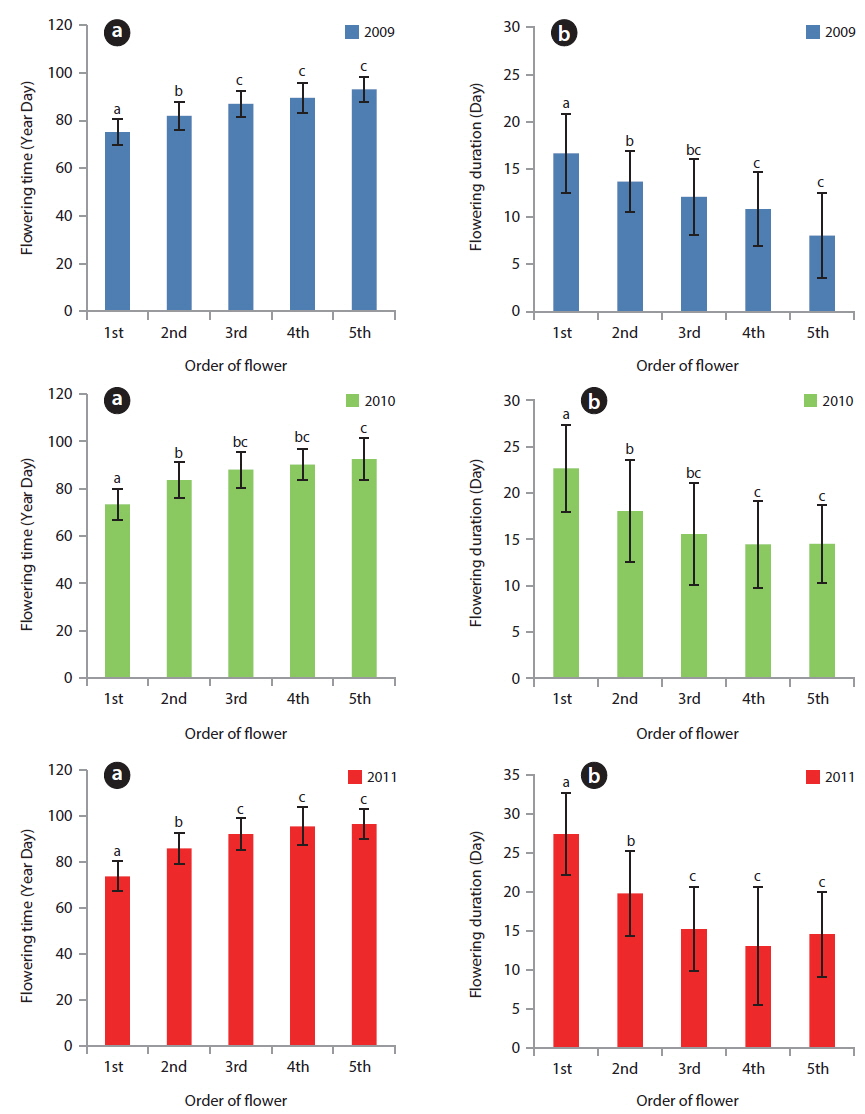
The YD at which cumulative flowering rate (CFR) reached 25% was 75 in 2009, 75 in 2010, and 79 in 2011; therefore, the difference between the three years was maximum of 3 days (Table 2). YDI (℃·day) and Tn (℃·day) at which CFR reached 25% were from 64.1 to 113.1 and from 7.7 to 27.0, respectively, and the difference between the three years was larger than that of YD. However, YD, YDI, and Tn at which CFR reached 50% during the three years were 80 to 87 of YD, 112.9 to 131.7 of YDI (℃·day), and 21.3 to 42.4 of Tn (℃·day). Thus, the differences between the three years increased in YD and decreased in YDI, and all three years were similar in Tn. YD, YDI, and Tn at which CFR reached 75% during the three years were 86-93 of YD, 146.2 to 167.3 of YDI (℃·day), and 34.8 to 48.2 of Tn (℃·day). Thus, the difference between the three years decreased in Tn only. On the other hand, CFR rates were closely related with YD, YDI, and Tn (Fig. 4) and their correlation coefficients were mostly over 0.9 and significant at 1% level (Table 3). However, their regression curves were sigmoid rather than linear, meaning that anyone of the three factors couldn’t be the most important for
[Table 2.] Date, YDI, and Tn at the flowering rate of 25%, 50%, and 75%
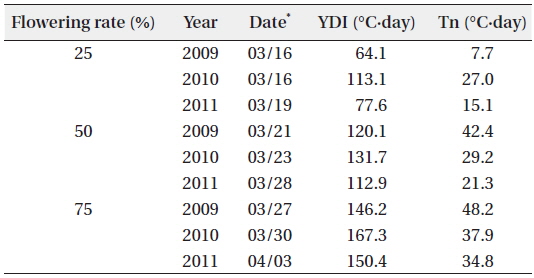
Date, YDI, and Tn at the flowering rate of 25%, 50%, and 75%
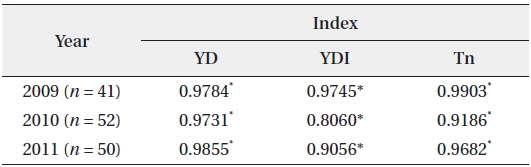
Correlation coefficients between cumulative flowering rate and YD, YDI, and Tn (n = dates from the first flowering time to the last one)
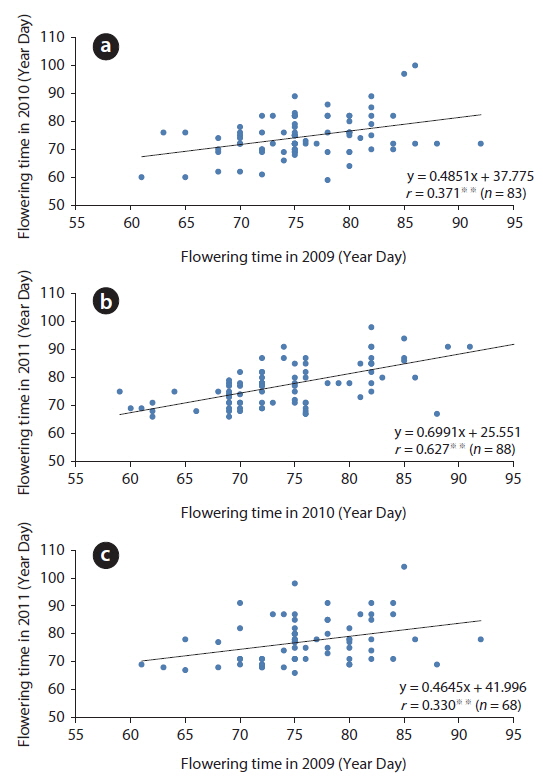
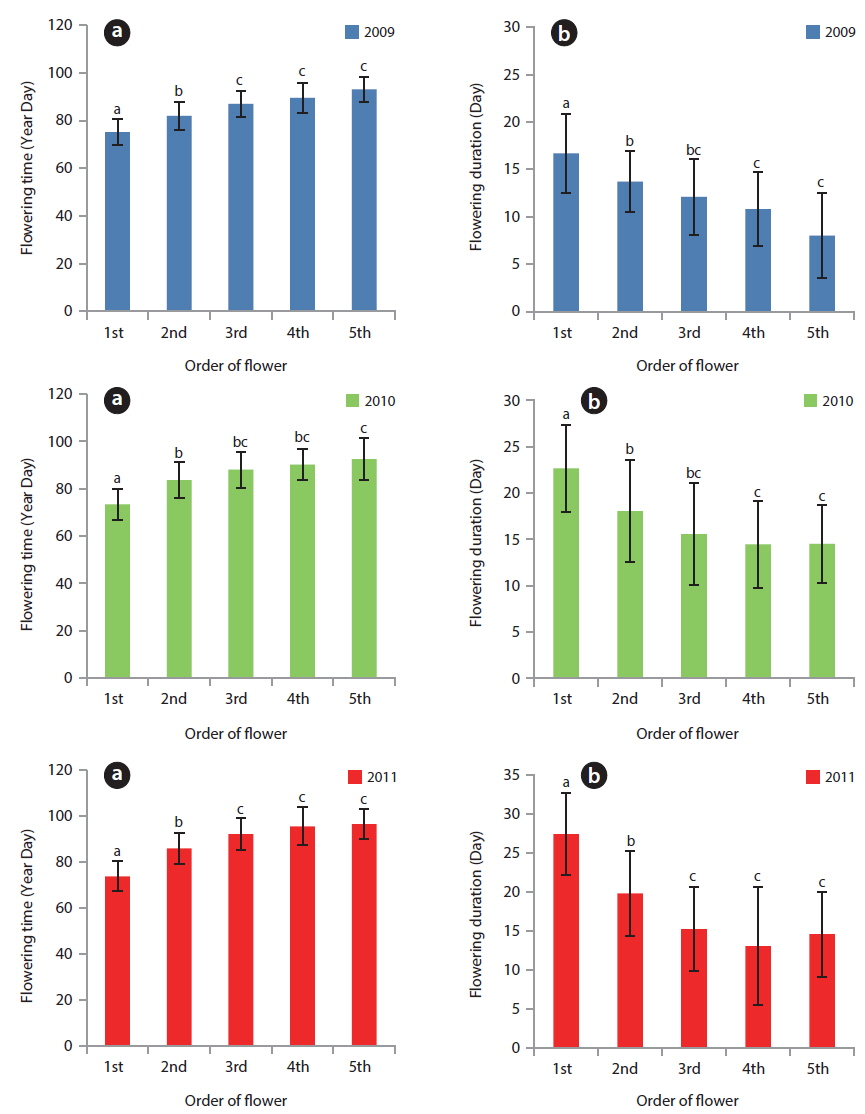
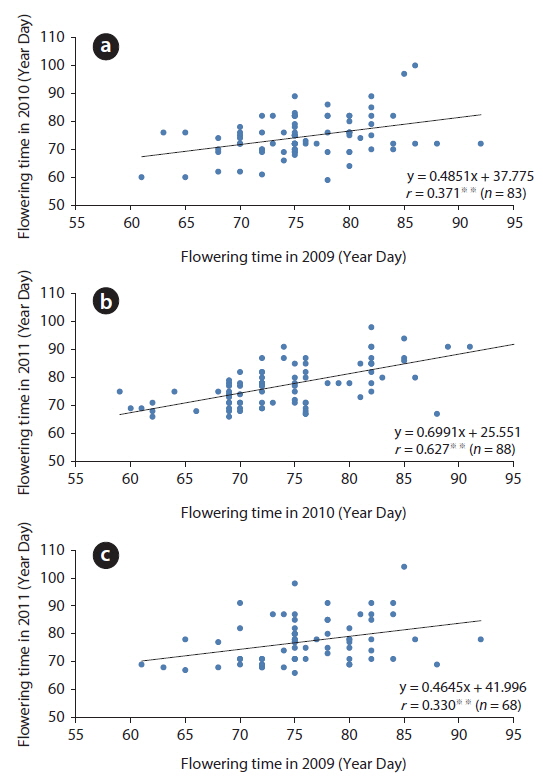
Based on the appearance of the first flower, the relative flowering time of each plant in a population was roughly constant for three years (Fig. 5). The correlation coefficients between two years were 0.371 (in 2009-2010,
>
Flowering time and other properties
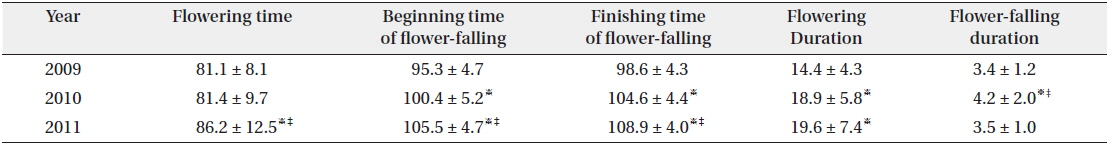
Flowering start times (mean ± SD) were 81.1 ± 8.1 YD in 2009, 81.4 ± 9.7 YD in 2010, and 86.2 ± 12.5 YD in 2011 (Table 4). Flowering duration was 14.4 ± 5.8 days in 2010 and 19.6 ± 7.4 days in 2011. Flower-falling duration was 3.4 ± 1.2 days in 2009, 4.2 ± 2.0 days in 2010, and 3.5 ± 1.0 days in 2011. Thus, flowering start time in 2010 was similar to that in 2009. However, flowering duration in 2010 was similar to that in 2011. Consequently, flowering started in early season and was maintained for a relatively long time in 2010. Thus, there was no relationship between the time and the duration of flowering at the population level. Flower-falling durations were similar in the three years. Moreover, the flowering duration was noticeably longer than the flower-falling duration in the three years. It is likely that the long flowering durations in 2010 and 2011 resulted from low air temperatures of mid-March. At the same time, Tn in 2010 and 2011 was lower than that in 2009.
[Table 4.] Average duration (days) or time (YD) of flowering and flower-falling
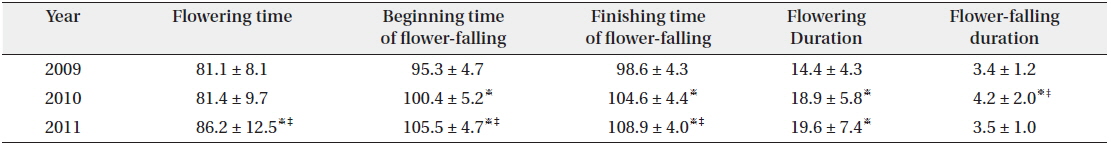
Average duration (days) or time (YD) of flowering and flower-falling
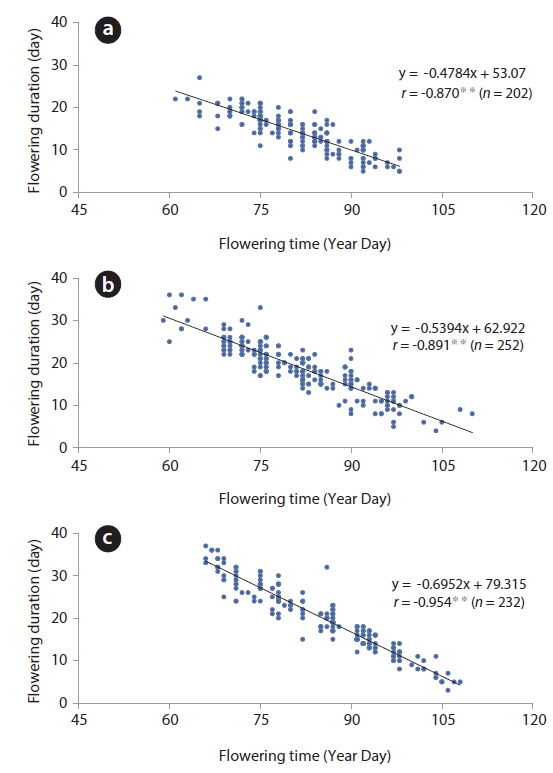
On the other hand, correlation coefficients between time and duration of flowering were below -0.8 and significant at 1% level (Fig. 6). Based on this result, the flowering duration was reversely related to flowering time at a flower level, meaning that the blossoms in early March could be maintained for long time periods. However, when air temperature was low (sub-zero) in mid-March, the flowers were maintained in blossom for a long time. Thus, as the flowering time and duration were considered, the latter does not appear to be related to the former but simply influenced by air temperature. Generally, the timing of flowering in plants is temperature-sensitive (Kudo and Hirao 2006). The life cycle of spring ephemerals inhabiting the deciduous forests is most sensitive to spring climatic conditions in which air temperature and snowmelt timing are major determinants of flowering initiation (Diekmann 1996, Lapointe 2001, Min et al. 2007). To this date, many studies investigated the relationship between the flowering time and air temperature. However, very few looked at the interrelation between flowering duration and air temperature. Hypothetically, low temperatures slow down the metabolism, restrict cell division and elongation, and thus reduce the growth rate of a plant (Ting 1982). Based on this, flower’s metabolism might be slowed down by low air temperature.
In this study,
In summary, my results indicated that the earlier the flower of