팔당호는 북한강, 남한강, 경안천이 합류하는 지점에 팔당댐이 축조되어 형성된 인공호로서 수도권의 주요 상수원이다(Kong, 2014). 팔당호로 유입되는 세 개의 하천은 특성이 상이하며, 넓은 유역으로부터 생활하수, 농업배수, 축산폐수 등의 유입이 지속되면서 부영양화가 진행되어, 규조류 및 남조류 등 조류의 과다증식이 국지적으로 발생하고 있다(Park and Jheong, 2003; Park et al., 2000). 특히 남조류에 의한 과다증식은 수돗물의 이취미(Bae et al., 2008)와 독소발생문제(Kim et al., 2010; Park et al., 2011)를 야기하여, 경관 및 친수활동 저해 등 상수원 관리에 어려움이 있다.
최근 팔당호에서 발생한 남조류의 과다증식은 주로 북한강에서 발생하였으며, 이 남조류는 팔당댐까지 영향을 미치는 것으로 나타났다(NIER, 2014). 2011년 11월의 경우 북한강에서 수온이 10℃ 이하의 저수온기 임에도 불구하고 남조류(
따라서 본 연구에서는 북한강, 남한강, 팔당호 등 팔당호 수역을 대상으로 퇴적층을 채집하여, 각 수온조건 변화에 따라 발생되는 조류군집 형성의 변화 및 그에 의한 이취미물질 발생여부를 파악하고자 하였다.
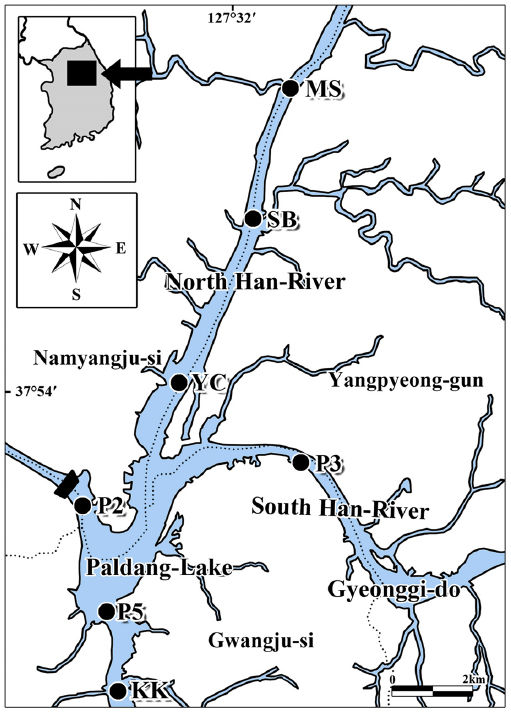
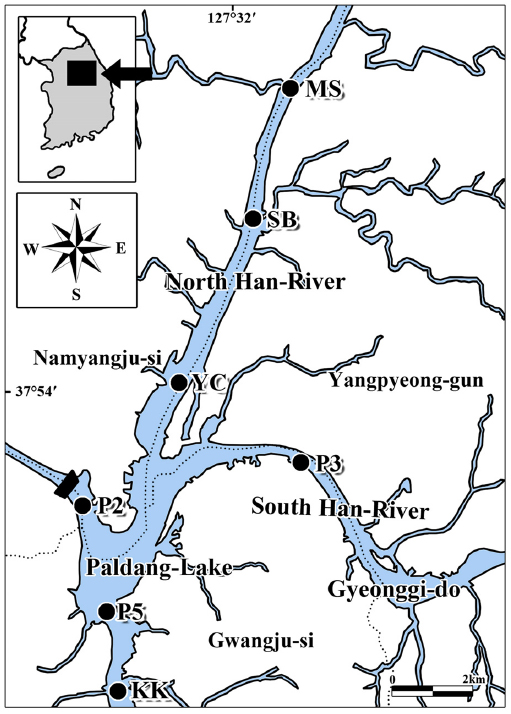
온도변화에 따른 퇴적층 배양과 식물플랑크톤 및 저서성남조류 군집발생 실험을 위해 북한강의 3개 정점(MS: 경기도 남양주시 화도읍 금남리 묵현천 합류부, SB: 삼봉리, YC: 조안면 진중리 양수철교), 남한강 1개 정점(P3: 경기도 양평군 양서면 신원리), 팔당호 및 경안천 합류부 3개 정점(P2: 경기도 남양주시 조안면 능내리 팔당댐, P5: 광주시 남종면 분원리, KK: 광주시 퇴촌면 광동리 광동교)을 대상으로 수행하였다(Fig. 1).
시료는 2014년 10월 6일에 grab sampler를 사용하여 5~10 cm 두께의 퇴적층을 채집하였다. 퇴적층의 배양은 멸균된 250 mL 삼각플라스크에 2±0.5 cm 높이로 퇴적층이 교란되지 않도록 100 mL를 주입하였고, 배양액은 팔당호 현장수를 식물플랑크톤 네트(mesh 5 µm)로 여과하여 100 mL을 주입하였다. 수온은 10, 15, 20, 25℃의 조건으로 40 μmol/m2·S의 광조건, 광주기 16(L):8(D)로 배양하였다. 퇴적층의 배양기간은 의암호와 공지천 퇴적층의 배양연구(Kim et al., 2014)와 퇴적층에서 남조류 발생연구(Hašler et al., 2004; Yamamoto and Nakahara, 2009)결과, 배양 10~15일이 식물플랑크톤 현존량이 증가하고, 저서성 남조류의 성장이 가장 활발한 것으로 연구되어, 본 연구에서는 14일간 배양하여 분석하였다.
각 채집지점의 이화학적 분석항목과 식물플랑크톤 발생현황은 물환경정보시스템(MOE, 2014)과 조류경보제 및 팔당호 조류 생리생태 연구(NIER, 2014)를 활용하였다. 퇴적층 배양에 따른 식물플랑크톤 군집분석은 표층수를 채집하여 Lugol’s solution으로 고정하였고, 저서성 조류는 퇴적표층 2×2 cm2 면적을 100 mL 증류수에 희석하여 포르말린으로 고정하였다. 식물플랑크톤과 저서성 조류의 분석은 Sedgwick-Rafter counting chamber를 사용하여 위상차 현미경(Nikon, Japan)하에서 100~1,000 배율로 개체수를 산출하였으며, 규조류는 Simonsen의 분류체계(Krammer and Lange-Bertalot, 1986, 1988, 1991a, 1991b)에 따라 정리하였고, 그 외 분류군은 Chung (1993), Hirose et al. (1977), John et al. (2002), Kormák and Anagnostids (2005)를 참고하였다.
휴면포자의 분석은 퇴적표층을 2 cm 깊이로 채집하여 습중량 1 g(w/w)을 증류수에 분산시켰다. 분산은 초음파기(WUC-A03, Korean)로 30초간 2회 처리하여 현탁액을 일정하게 분쇄하였다. 분산액은 90 µm, 60 µm Nylon재질의 체로 순차적으로 분리하였으며, 최종적으로 10 µm Nylon 체로 여과한 퇴적물 시료를 Panning methods (Matsuoka and Fukuyo, 2000)를 사용하여 휴면포자를 분리하였다. 분리된 휴면포자는 10 mL 부피로 농축하여 식물플랑크톤 검경과 동일하게 100~400배율로 현존량을 산출하였다.
이취미물질의 분석은 먹는물 수질감시항목 운영지침(MOE, 2011)에 따라 GC/MS(Bruker, 450GC/320MS)를 이용하여 headspace(HS)-solid phase microextraction (SPME)법으로 geosmin과 2-MIB(2-methyliso-borneol)를 분석하였다.
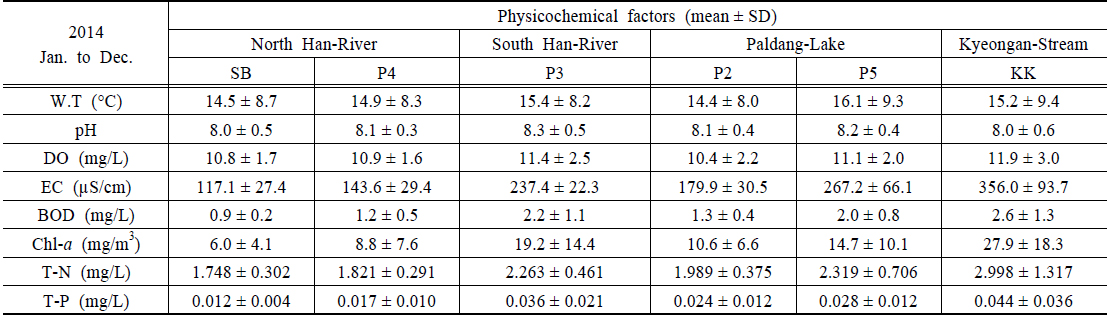
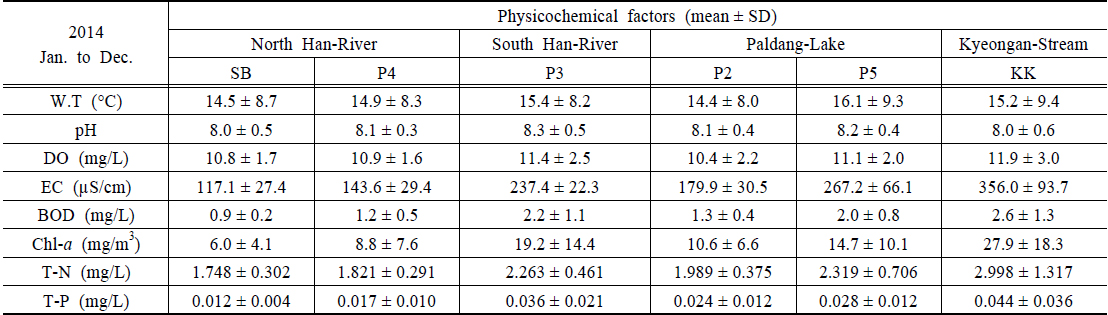
조사지점의 이화학적 수질은 앞에서 제시한 수질측정망자료(MOE, 2014)를 활용하였으며, 북한강의 경우 삼봉(SB)지점과 양수철교(YC)지점에 인접한 P4(한강물환경연구소 앞)지점의 자료를 활용하였다(Table 1). 연평균 수온은 14.5±8.7~16.1±9.3℃의 범위로 P5지점에서 약간 높았으며, 2014년 10월 6일 퇴적층 채집 시 20.2~20.7℃의 범위로 지점간 차이가 거의 없었다. pH는 8.0±0.5~8.3±0.5의 범위로 지점 간 큰 차이를 보이지 않았으나, 채집 시 경안천 지점(KK)이 9.1로 다소 높았다. 용존산소(DO)는 10.4±2.2~11.9±3.0 mg/L의 범위로 남한강(P3)과 KK지점이 높았고, 전기전도도(EC)는 117.1±27.4~350.0±93.7 µS/cm의 범위로 북한강 수역이 대체로 낮으며, KK가 가장 높게 나타났다. BOD는 0.9±0.2~2.6±1.3 mg/L의 범위로 경안천에서 가장 높았으며, 채집 시 모든 지점에서 2 mg/L 이하로 양호한 수질을 나타냈다(NIER, 2014). 엽록소
Mean values of physicochemical factor values in North Han-River (SB, P4) South Han-River (P3), Paldang-Lake (P2, P5) and Kyeongan-Stream (KK), 2014
팔당호에서 채수 및 여과하여 배양에 사용된 현장수는 총인이 0.004 mg/L 수준으로 낮았으나, 퇴적층시료에 주입한 이후에 0.042~0.044 mg/L의 범위로 증가 하였다. 배양 시작 시 총인의 농도는 부영양 상태(Forsberg and Ryding, 1980)로 식물플랑크톤 및 저서성 조류의 성장에 인이 제한요인으로 작용하지 않을 것으로 판단된다.
3.2. 휴면포자(akinetes)의 분포와 Anabaena의 발생량
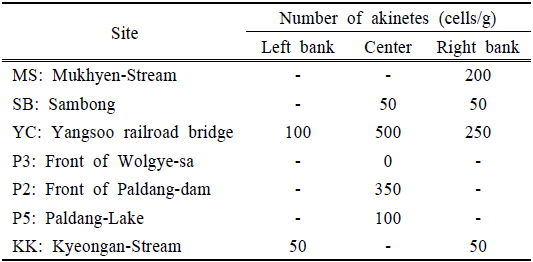
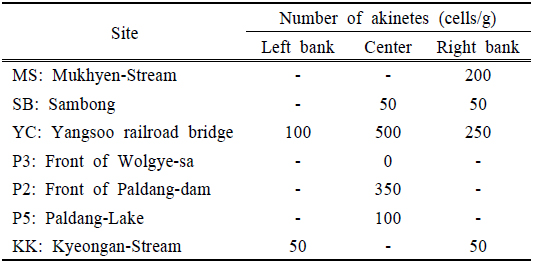
북한강, 남한강 및 팔당호 지역의 휴면포자 분포 조사 결과 0~500 cells/g (w/w)의 범위로 양수철교(YC) 중앙에서 현존량이 가장 높은 것으로 나타났다. 북한강에서는 유입천인 묵현천 합류부(MS)에서 200 cells/g (w/w)으로 하류의 삼봉(SB) 지점보다 높게 나타났다. SB는 2014년 7월~9월에 유해 남조류인
팔당호 댐앞(P2) 지점은 휴면포자 현존량이 350 cells/g(w/w)수준으로 검출되었다. P2지점에서 출현한
Distribution of akinetes in North Han-River (MS, SB, TC) South Han-River (P3), Paldang-Lake (P2, P5) and Kyeongan-Stream (KK), 2014
휴면포자의 발아율(gemination rate)은 대상 분류군이나 조건에 따라 다르지만 배양 1~3일 동안 10~15℃에서 3~12%, 20~25℃조건에서 17~48%로 알려져 있다(Baker and Bellifemine, 2000; Fay, 1988; van Dok and Hart, 1997). 공지천과 의암호의 연구에서는 퇴적층 배양 3일 후 휴면포자의 현존량이 10% 수준으로 감소하였으며,
3.3. 퇴적층의 온도별 배양에 따른 식물플랑크톤 군집 발생
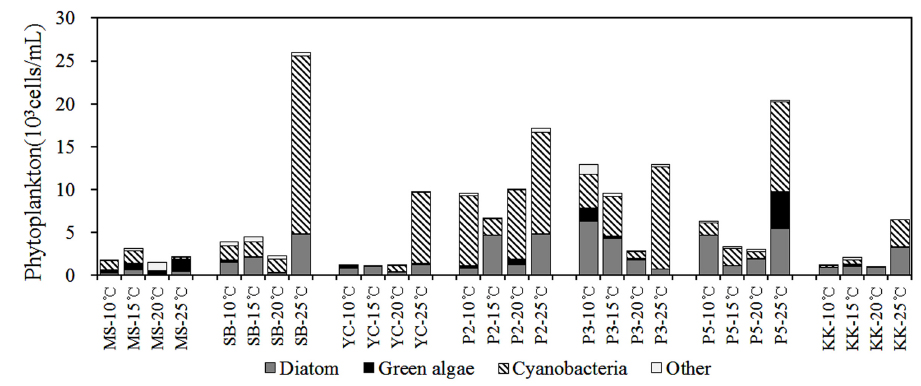
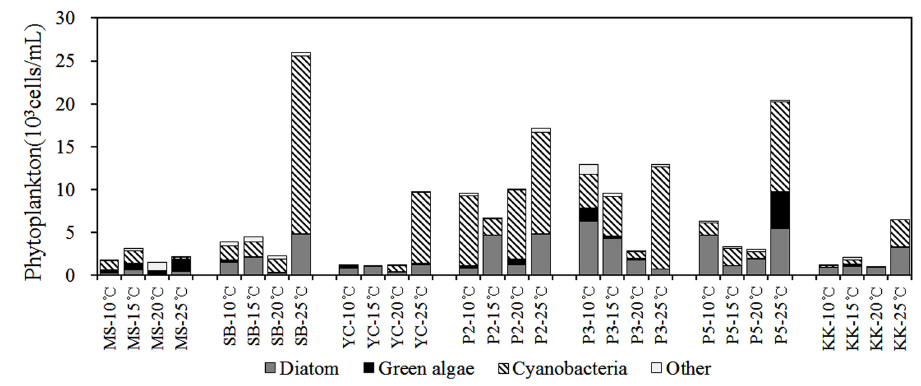
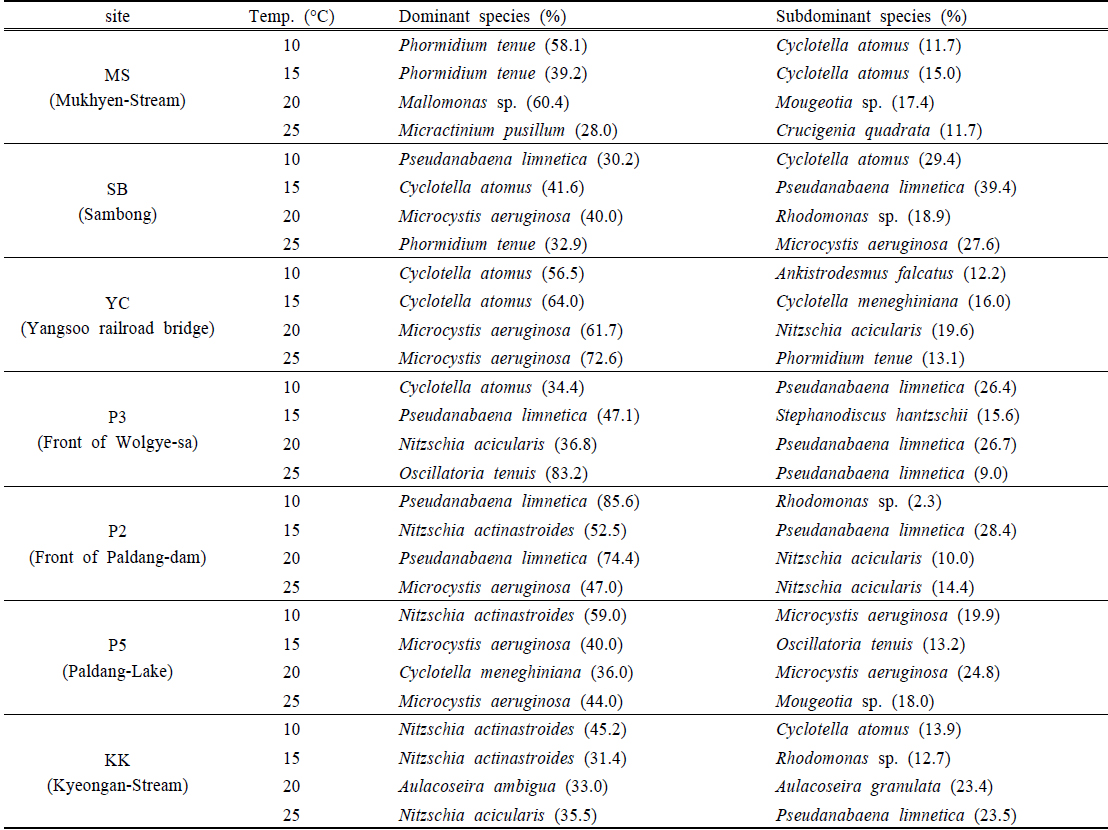
북한강, 남한강 및 팔당호 퇴적층의 온도별 배양에 따른 식물플랑크톤 군집은 전체 68분류군이 출현하였으며, 규조류가 29분류군으로 가장 다양하였으며, 녹조류 19분류군, 남조류 10분류군, 갈색편모조류 등 기타조류가 6분류군 출현하였다. 조사 지점에 따라 각 지점별 6~20분류군이 출현하였으며, 양수철교(YC) 15℃ 배양시료에서 가장 적은 분류군이 출현하였고, 팔당댐(P2) 20℃ 배양시료에서 가장 다양한 분류군이 출현하였다. 공지천 및 의암호의 퇴적층 배양연구(Kim et al., 2014)에서 31일간 배양한 결과 40~45분류군, Hašler et al. (2004)의 퇴적층 배양 연구에서도 40일간 49분류군이 출현한 것과 비교 할 때 출현 분류군이 다양하였다. 이것은 7개 조사지점과 4개 배양조건 총 28개의 각기 다른 배양조건으로 각각의 출현 분류군은 공지천 및 의암호 퇴적층 배양연구(Kim et al., 2014)에서 배양 14일 까지 14~18분류군 출현한 것으로 동일 기간 내 출현한 분류군수는 본 연구결과와 유사한 것으로 나타났다.
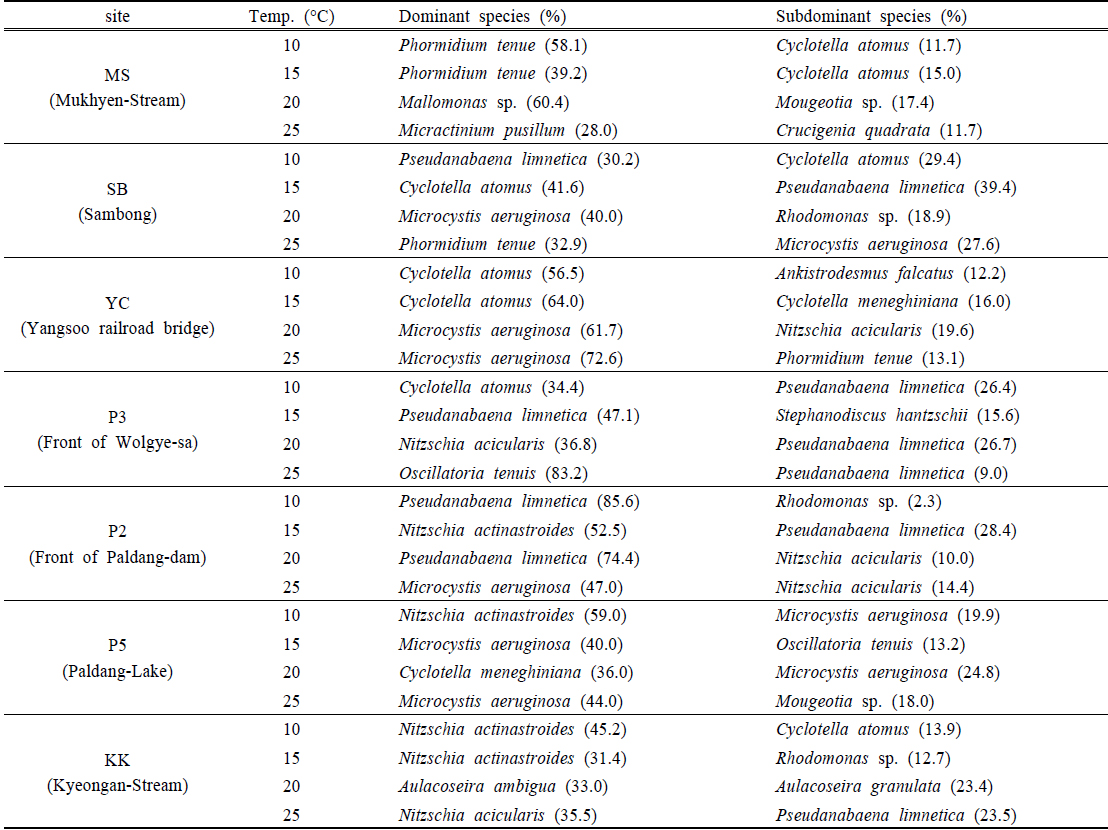
전 지점의 주요 출현종은
Dominant and subdominant species of phytoplankton for sediment incubation by various water temperatures
YC지점 퇴적층은 10~15℃의 배양조건에서
P2지점의 퇴적층은 10~20℃의 배양조건에서
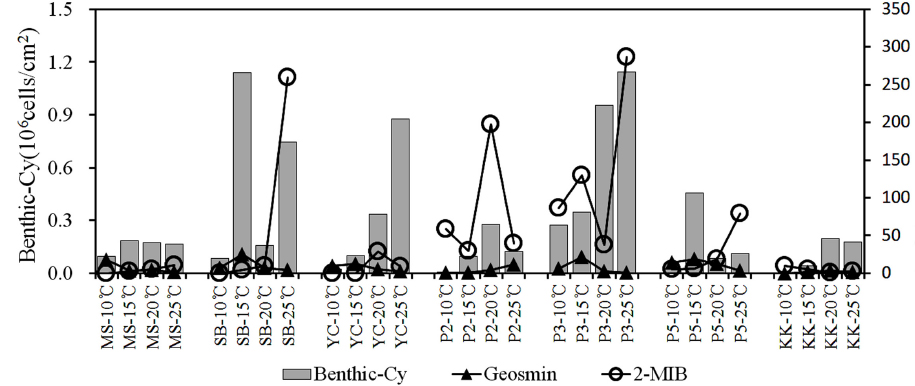
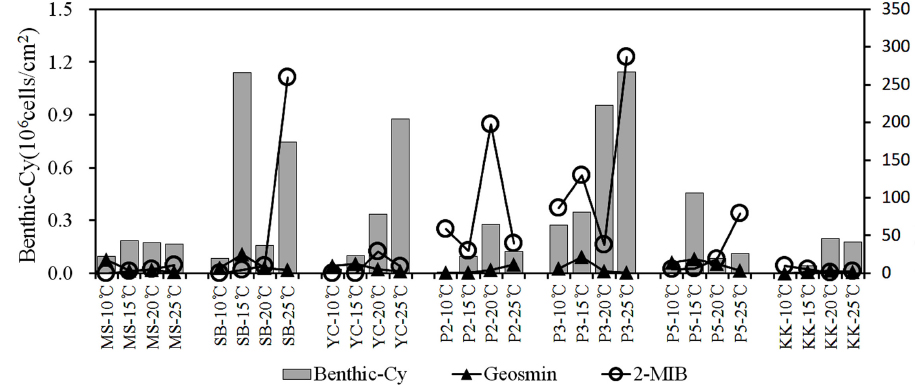
3.4. 저서성 남조류의 발생과 이취미 물질(geosmin, 2-MIB)
일반적으로 이취미물질은 조류의 성장기와 안정기에는 체외로 거의 배출되지 않지만 사멸기에는 체내 축적된 물질이 체외로 배출되는 것으로 알려져 있다(Bae et al., 2008; Jüttner and Watson, 2007). 저서성 남조류는 남조매트의 형성과 부상시기에 성장과 사멸을 반복하면서 이취미 물질의 농도가 증가하는 것으로 연구되었으며(Kim et al., 2014), 이 때문에 남조매트의 형성은 이취미 물질의 발생여부를 확인 할 수 있는 지표로 활용될 수 있다.
각 실험조건에서 퇴적표층의 변화는 10~15℃의 조건에서 배양 7일까지는 큰 변화가 없었으며, 배양 14일에 15℃ 조건의 삼봉(SB), 남한강(P3), 팔당호(P5)지점에서 남조류 사상체가 일부 관찰되었다. 20℃ 조건에서는 배양 7일에 양수철교(YC), 팔당댐(P2), P3지점에서 부분적으로 남조매트가 관찰되었으며, 배양 14일에 P3 지점이 남조매트가 뚜렷하게 형성되었다. 25℃는 YC, P2, P3지점에서 남조매트가 관찰되었으며, P3지점에서 남조류 사상체가 일부 표층으로 부유된 것을 확인하였으나, 실험종료 시까지 매트의 부상은 관찰되지 않았다.
각 지점에서 출현한 저서성 남조류는 0~1.15×106 cells/cm2의 범위로, 경안천(KK)지점에서(10℃) 저서성 남조류가 출현하지 않았으며, P3지점에서(25℃) 가장 높은 현존량을 나타냈다. 묵현천(MS)과 KK지점은 모든 온도조건에서 0.2×106 cells/cm2 이하로 낮았으며, SB의 15℃에서 1.14×106 cells/cm2 , P5의 15℃에서 0.45×106 cells/cm2 로 출현하는 등 저서성 남조류의 현존량은 온도조건보다 조사지점의 퇴적층 발달과 관련이 있을 것으로 판단된다. 출현종은 저온시기에 주로
수계에서 발생되는 이취미물질은 남조류의 성장과 밀접한 관련이 있으며, 부유성 남조류 뿐만 아니라 저서성남조류의 증식에 의해서도 영향을 받는다고 보고되고 있다(Jüttner and Watson, 2007). 본 연구의 결과 팔당호 수역에서는 이취미 원인종인
북한강, 남한강 및 팔당호 지역의 퇴적층의 온도별 배양에 따른 조류군집 형성과 이취미 발생특성을 분석한 결과 다음과 같은 결론을 얻었다.
1) 퇴적층에서
2) 식물플랑크톤 군집은 배양기간 동안 전체 68분류군이 출현하였으며, 규조류가 29분류군으로 가장 다양하였고, 주요 출현종은
3) 식물플랑크톤 현존량은 0~20.7×103 cells/mL로 유해 남조류인
4) 퇴적표층에서는
5) 이취미 물질은 geosmin이 최대 25.5 ng/L, 2-MIB가 286.8 ng/L로 모든 온도조건에서 이취미가 검출되었으며, 이취미원인종으로 출현한