지구온난화로 인한 수온 상승과 초식자인 성게류의 증가로 인하여 세계적으로 600,000 km2의 면적을 차지하며 연간 4억 6천만 톤의 탄소를 흡수하는 대형해조류로 구성된 바다숲이 지속적으로 감소되고 있는 실정이다(Muraoka, 2004; Harley et al., 2012; Ordoñez et al., 2014). 대형해조류를 포함하는 해조류는 해초류, 식물성플랑크톤과 함께 해양생태계의 중요한 일차생산자로서 영양염 순환, 탄소순환, 어류 및 무척추동물을 포함한 다양한 해양생물의 은신처, 산란장 및 먹이장으로서 생태학적 기능을 수행한다(Lindstrom, 2009; Whitaker et al., 2010). 이러한 연유로 해조류는 연안생태계에서 먹이사슬의 상층에 있는 해양생물의 종다양성과 풍부도 및 어장형성에 영향을 미친다(Wan et al., 2009; Janiak and Whitlatch, 2012; Satheesh and Wesley, 2012). 또한, 해조류는 연안의 도시화와 산업화로 인해 해양에 투기되는 유기 및 무기 오염물을 제거하는 생물여과자(biofilter) 뿐만 아니라, 연안환경을 모니터링하는 생물지표자(biological indicator)로도 사용되고 있다(Worm et al., 2000; Wells et al., 2007; Scherner et al., 2013).
연안생태계의 일차생산자인 해조류의 군집은 인간활동에 기인한 다양한 환경스트레스(교란, 퇴적, 영양염 유입 등)로 인하여 생산성이 높은 수관부 형성(canopy-forming) 해조류에서 생산성이 낮은 덤불형성(turf-forming) 해조류로 단계적 변이(phase shift)가 일어나고(Kraufvelin et al., 2010), 종다양성 감소, 홍조류와 갈조류의 출현종수(출현종 비율) 감소, 그리고 생물량과 피도의 감소가 일어나며(Whitaker et al., 2010), 반대로 기회종인 녹조류의 생물량이 증가되는 등 점진적 변화가 발생한다(Arévalo et al., 2007; Pinedo et al., 2007; Wells et al., 2007). 인간 유래의 환경변화로 인한 해조상과 군집구조에 관한 연구는 전 세계적으로 활발하게 수행되고 있으며, 최근에는 지구온난화로 인한 수온상승과 해양산성화도 해조류의 생장, 분포 및 군집 구조에 영향을 미치는 것으로 확인되고 있다(Augyté and Shaughnessy, 2014). 따라서, 해조류의 종다양성과 군집구조에 대한 자료를 축적하는 것이 절실하게 필요한 실정이다(Orfañidis et al., 2001; Diaz-Pulido et al., 2011; Roleda et al., 2012).
우리나라 남해안 해안선은 굴곡이 심하고, 크고 작은 섬들로 구성되어 있을 뿐 만 아니라 해류, 수온, 염분, 수심, 영양염 및 탁도 등이 달라서 매우 복잡한 환경적 특성을 나타내며(Kang, 1966; Choi, 1992), 쿠로시오 난류의 영향으로 해조류의 식생이 풍부하고 생육하는 해조류의 종조성이 상이하여 보다 세밀하고 집중적인 연구가 필요한 해역이다(Sohn, 1987). 한국 남해안의 해조류에 대한 연구는 Kang (1966)의 “한국산 해조류의 지리적 분포”를 시작으로 해조상(Song et al., 1970; Kang and Lee, 1979; Oh et al., 2002; Choi, 2008), 생물량(Choi, 2008; Kim et al., 2008)과 수직분포(Hwang et al., 1997; Kwak and Huh, 2009)에 관한 많은 연구가 수행되었다. 특히, Choi (2008)는 2005년에 한려해상국립공원 내 24개 정점의 1–2회 조사에서 해조류 89 종을 동정하였고 수직분포를 기록하였으며, 7개 정점에서 우점종과 생물량을 보고하였다.
남해안 한려해상국립공원은 화려한 경관과 해수욕장이 잘 발달되어 관광객이 증가하면서 유·무기 오염물의 해양 투기량도 급증하고 있으며, 환경변화가 심한 지역이다(Choi, 2008; Ahn et al., 2013). 이러한 한려해상국립공원의 건강한 연안생태계 유지를 위하여 일차생산자의 보존 및 복원을 위해 기초자료 확보가 매우 중요한 시점이다. 따라서, 본 연구는 한려해상국립공원 내 서식하는 해조류의 생태학적 특성과 군집특성을 파악하기 위해 해조상, 우점종과 생물량의 계절적 변화양상을 파악하고 10년 전에 수행된 연구결과와 비교하여 환경변화와 해조류 군집구조 변화의 관계를 고찰하였다.
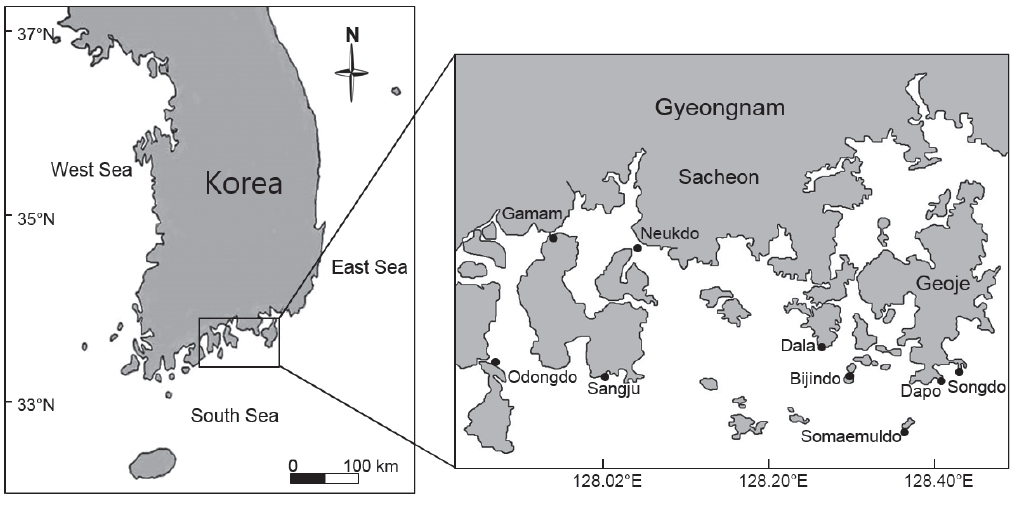
한려해상국립공원 내 9개 정점에서 2014년 3월(동계), 5월(춘계), 8월(하계), 10월(추계)에 계절별로 최 간조기에 해조류를 정량 및 정성 채집 하였다(Fig. 1). 정량조사는 조간대 상·중·하부 및 조하대 상부에 각 2-3개의 방형구(50 cm×50 cm)를 무작위적으로 놓고 방형구 내에 존재하는 해조류를 전량 채집하였으며, 해조상을 파악하기 위하여 다양한 기질과 조위에서 서식하는 모든 해조류를 정성 채집하였다. 채집된 해조류는 포르말린-해수 용액(5-10%)으로 현장에서 고정시켜 실험실로 운반한 후 현미경을 사용하여 분류 및 동정하였으며, 출현종 목록 및 국명은 Lee and Kang (2002)에 따랐다. 해조류의 기능형은 출현종의 외부형태, 내부구조, 광합성효율 등에 따라 엽상형(sheet form), 사상형(filamentous form), 성긴분기형(coarsely branched form), 다육질형(thick leathery form), 유절산호말형(jointed calcareous form), 각상형(crustose form)의 6개 그룹으로 구분하였다(Littler and Littler, 1984). 해조상의 특성을 파악하기 위하여 정성 채집된 해조류를 분류 및 동정한 후 출현종수로 갈조류에 대한 녹조류의 비(C/P), 갈조류에 대한 홍조류 비(R/P), 갈조류에 대한 녹조류 및 홍조류의 비로 (R+C)/P를 구하였다(Feldmann, 1937; Segawa, 1956; Cheney, 1977).
정량 채집된 해조류는 담수로 수회 세척하여 모래와 불순물을 제거하고, 종별로 구분하여 105℃로 설정된 건조기에서 24-48h 동안 건조한 후 건중량을 측정하였고 단위면적당 생물량(g dry wt/m2)으로 환산하였다. 해조류 종별 생물량 평균값과 단위 면적당 생물량 구성비(%)를 구하여 생물량 구성비가 30% 이상인 종은 우점종으로, 10-30%인 종은 준우점종으로 구분하였다(Kim et al., 1995; Kim et al., 1997). 또한, 정점별 출현종과 생물량 자료를 이용하여 풍부도지수(richness index, R), 다양도 지수(diversity index, H')와 균등도지수(evenness index, E)를 계산하였다(Margalef, 1958; Fowler and Cohen, 1990). 우점도지수(dominance index, DI)는 군집 내에서의 생물량 순서에 따라 제1, 2우점종을 선택하고 2종의 생물량 합에 대한 총 생물량의 비율로 산출하였다(McNaughton, 1967; Lee et al., 1983). 해조류의 출현종 유·무로 정점간 유사도를 분석하였으며, 그룹간 유의차는 SIMPROF (similarity profile) test로 검정하였다. 또한, 그룹간 유의차가 확인되면 SIMPER (similarity percentage) 분석을 실시하여 각 그룹간 비유사도(dissimilarity)에 기여하는 종을 확인하였다. 군집지수의 산출 및 도식화에는 PRIMER version 6 (Clarke and Gorley, 2006)을 사용하였다.
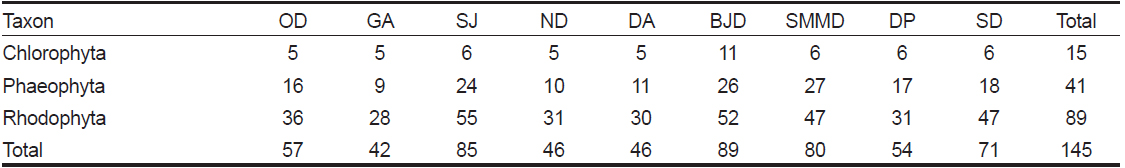
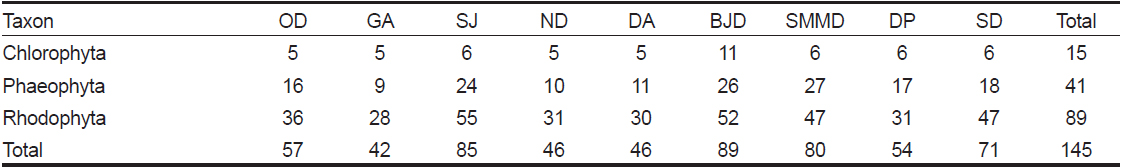
본 연구기간에 한려해상국립공원 내 9개 정점에서 출현한 해조류는 녹조류 15종, 갈조류 41종, 홍조류 89종이었으며, 홍조류는 전체 출현종인 145종의 61.38%를 차지하여 녹조류와 갈조류에 비해 출현빈도가 높았다. 계절별 출현종수는 동계에 100종으로 최대였고 춘계에 92종, 하계에 88종, 추계에 81종으로 가장 적었다(Table 1). 한려해상국립공원 9개 정점에서 연 중 관찰되는 해조류는 구멍갈파래(
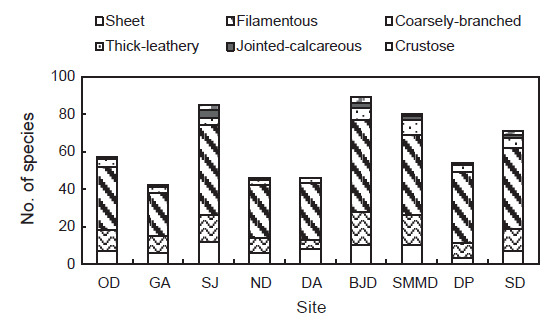
The number of macroalgal species observed at Hallyeohaesang National Park, South coast of Korea
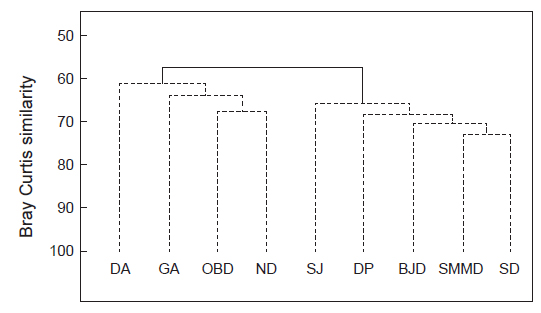
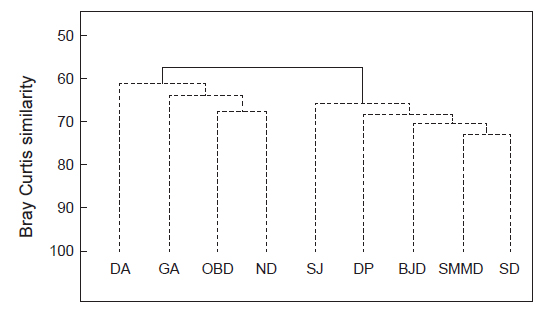
해조류 출현종 유·무를 근거로 하여 9개 정점의 유사도를 분석하여 보면, 그룹 A (달아, 감암, 오동도, 늑도)와 그룹B (상주, 다포, 비진도, 소매물도, 송도)로 구분되었으며, 그룹 A는 여수-남해 지역, 그룹B는 통영-거제 지역으로 지역적 차이를 보였다(Fig. 2). 그룹간 차이는 57.47%로 통계적 유의차를 보였다(SIMPROF test,
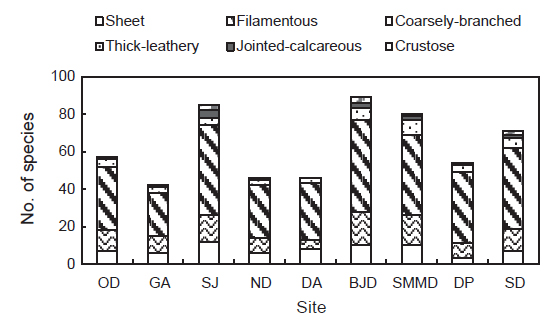
한려해상국립공원 9개 정점에서 출현한 해조류 145종에서 성긴분기형이 71종(48.97%), 사상형이 32종(22.07%), 엽상형이 18종(12.41%), 다육질형이 14종(9.66%), 각상형이 6종(4.14%), 유절산호말형이 4종(2.76%)으로 구분되었다. 출현종수를 근거로 한려해상국립공원 해역에서 가장 우점하는 해조류 기능형군은 48.97%를 차지하는 성긴분기형이었다. 또한, 정점별로 기능형 해조류의 출현종수는 다르게 나타났으며, 성긴분기형은 23-49종으로 감암에서 최소였고 비진도에서 최대였으며, 사상형은 5-18종으로 달아에서 최소였고 비진도에서 최대로 확인되었다. 엽상형은 3-12종으로 다포에서 최소, 상주에서 최대였고, 다육질형(3-8종)은 감암, 늑도, 달아에서 최소, 소매물도에서 최대로 나타났다. 각상형은 상주와 비진도에서 3종으로 최대, 송도 2종, 소매물도 1종을 제외한 다른 정점에서는 확인되지 않았다. 유절산호말형은 달아에서 채집되지 않아 최소를 보였으며 상주에서 4종으로 최대였다(Fig. 3).
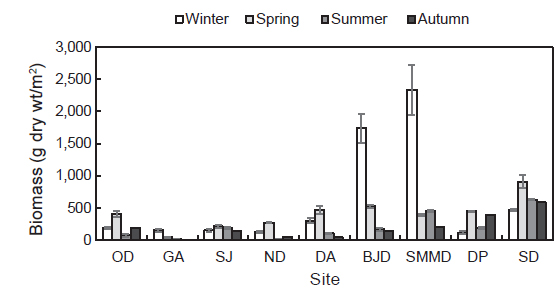
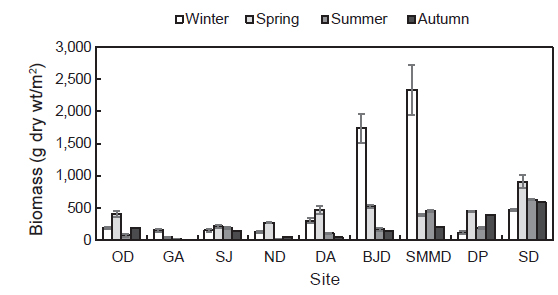
국립공원 내에 서식하는 해조류의 연평균 생물량(g dry wt/m2)은 358.00 g/m2 였고 소매물도에서 847.64 g dry wt/m2 로 최대였고 감암에서 56.51 g dry wt/m2로 최소였다. 계절별 생물량은 9개 정점에서 평균 196.04-621.08 g dry wt/m2로 춘계에 최소였고 동계에 최대로 확인되었다. 정점별 생물량은 비진도와 소매물도는 동계에 최대였고 이곳을 제외한 연구 정점에서는 춘계에 최대였다. 오동도에서 생물량은74.48-183.72 g dry wt/m2로 하계에 가장 낮았고 동계에 가장 높은 값을 보였다. 감암 연안의 해조류 생물량은 추계에 최소(4.70 g dry wt/m2)였고 동계에 최대(153.37 g dry wt/m2)였으며, 상주에서는 142.02-213.59 g dry wt/m2로 추계에 최소, 춘계에 최대였다. 늑도의 해조류 생물량은 하계와 춘계에 9.47과 273.59 g dry wt/m2로 최소와 최대 값을 기록하였다. 또한, 달아에서도 생물량은 43.59 g dry wt/m2으로 추계에 최소였고 춘계에 470.08 g dry wt/m2로 최대로 나타났다. 비진도와 소매물도에서 생물량은 추계에 148.99 g dry wt/m2과 200.59 g dry wt/m2로서 최소였고 동계에 1,740.90 g dry wt/m2과 2,340.26 g dry wt/m2 으로 최대를 보였으며, 다포와 송도에서는 동계에 116.88 g dry wt/m2과 474.19 g dry wt/m2로 최소였고 춘계에 452.88 g dry wt/m2과 910.08 g dry wt/m2 로서 최대로 확인되었다(Fig. 4).
해조류의 연평균 생물량의 구성비를 근거로 한 우점종(30% 이상)과 준우점종(10-30%)을 살펴보면, 오동도에서는 우점종이 없었으며, 모자반류 3종과 작은구슬산호말이 준우점종이 없으며, 감암에서는 구멍갈파래(35.45%)가 우점하였다. 상주에서는 지충이가, 늑도에서는 구멍갈파래(35.17%)와 참모자반(30.44 %, S. fulvellum)이 우점종으로 나타났다. 달아와 비진도에서는 괭생이모자반이 전체 생물량의 49.30%와 58.76 %를 차지하였고 또한, 괭생이모자반은 소매물도에서도 51.30%를 그리고 동계에는 생물량의 74.26%를 보였다. 다포에서 우점종은 생물량의 47.08%를 차지하는 지충이였고 송도에서는 홍조류 작은구슬산호말(45.50 %)로 확인되었다(Table 2).

Dominant (≥ 30%) and subdominant (10-30%) species in terms of average biomass percentage (%) at nine study sties in Hallyeohasang National Park, South coast of Korea
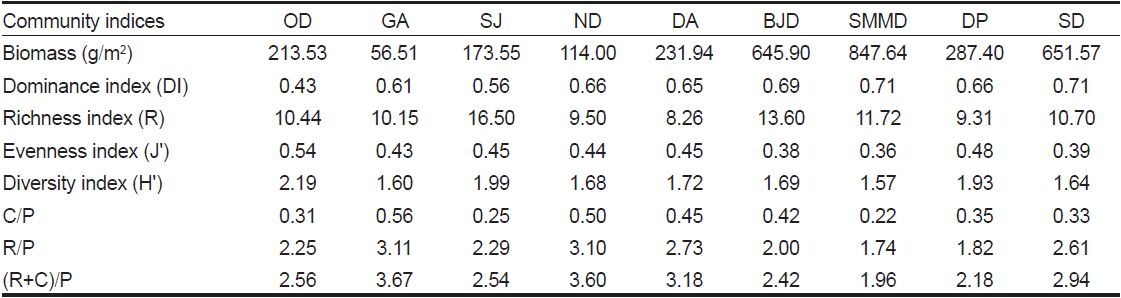
해조류 생물량과 출현종수로 계산된 군집지수를 살펴보면, 한려해상국립공원의 우점도지수(DI)는 0.63였고 오동도에서 0.43으로 최소였고 소매물도와 송도에서 0.71로 가장 높았다. 풍부도지수(R)는 평균 11.13 (정점별 8.26-16.50)으로 나타났으며 달아에서 최소였고 상주에서 최대였다. 출현종수와 생물량에 따라 변화하는 균등도지수(J')는 정점별 평균 0.43을 보였으며 소매물도에서 0.36으로 최소였고 오동도에서 0.54로 최대를 보였다. 출현종수와 종간의 풍부도 데이터(생물량)의 분포인 균등도지수에 의해 결정되는 다양도지수(H')는 균등도지수와 동일하게 소매물도에서 최소(1.57), 오동도(2.19)에서 최대값을 나타냈다. 해조상의 지역적 특성을 나타내는 C/P, R/P, (R+C)/P값은 각각 0.22-0.56, 1.74-3.11, 1.96-3.67의 범위로 소매물도에서 최소였고 감암에서 최대값을 나타냈다(Table 3).
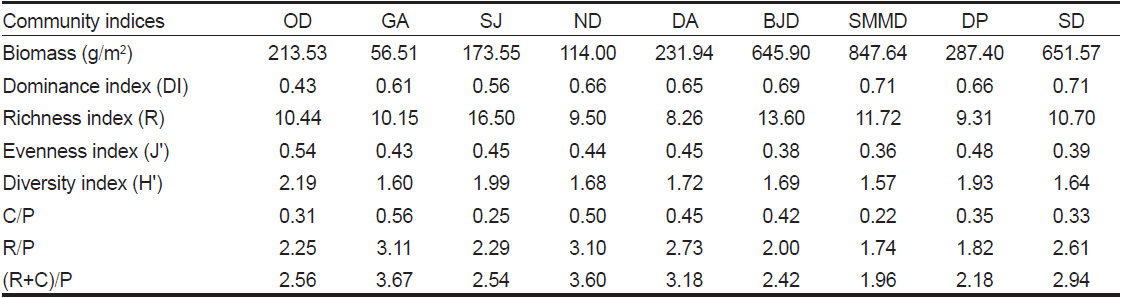
Average biomass (g dry wt/m2 ) and various community indices of seaweeds at the nine study sites in Hallyeohaesang National Park, South coast of Korea
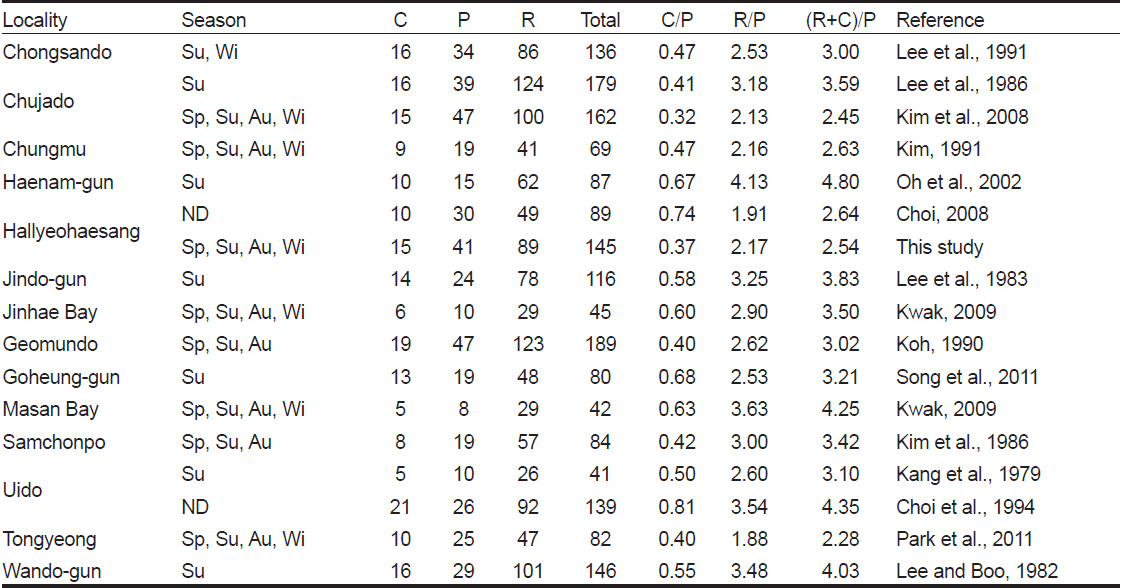
한려해상국립공원 내 9개 정점에서 출현한 해조류는 총 145종(녹조 15종, 갈조 41종, 홍조 89종) 이었으며, 정점별로 42-89종으로 확인되었다. 인근 해역의 해조류 출현종수는 충무에서 69종(Kim, 1991), 통영에서 82종(Park et al., 2011), 진해만에서 45종(Kwak, 2009), 거문도에서 189종(Koh, 1990), 마산만에서 42종(Kwak and Huh, 2009), 삼천포에서 84종(Kim et al., 1986)으로 본 해역의 해조류 출현종수는 인근 해역에 비해 높은 수준이었다. 또한, Choi (2008)의 2005년 조사에서 한려해상국립공원에 89종의 해조류가 서식함을 보고하였는데, 본 연구에서는 56종이 더 많은 145종이 기록되었다. 이외에도 해조류 출현종수는 본 연구정점의 서쪽(서남해안)에 위치한 해남에서 87종(Oh et al., 2002), 진도에서 116종(Lee et al., 1983), 고흥군 무인도서에서 80종(Song et al., 2011), 신안군 우이도에서 41-139종(Kang et al., 1979; Choi et al., 1994), 완도군에서 하계에 146종(Lee and Boo, 1982)이 보고되었으며, 완도군 청산도의 3지역(당리, 진산리, 동촌리)의 조간대와 조하대에서 2계절(하계와 동계)에 출현한 해조류는 136종으로 기록되었고(Lee et al., 1991), 제주도 추자도(예초와 후포)에서는 162종이 서식하는 것으로 확인되었다(Kim et al., 2008). 따라서, 한려해상국립공원 해역에 서식하는 해조류 출현종수는 서남해 해역에 비해 높게 나타났고 이는 상대적으로 청정해역인 연구해역이 해조류 서식지로 적합한 환경이라고 할 수 있겠으나, 해조상은 환경변화, 채집방법 및 채집시기에 따라 달라지기 때문에 한려해상국립공원 해역의 해조류 종다양성에 대한 논의는 좀더 자료가 축적된 후에 논의되어야 하며 효율적인 해조자원의 관리 및 변화를 모니터링하기 위한 연구 노력이 요구된다.
해조류는 생태학적 특성에 따라 다년생의 느린 생장을 나타내는 해조류인 다육질형, 성긴분기형, 유절산호말형, 각상형 해조류와 교란이나 환경오염 해역에서 빠른생장 을 보이는 엽상형과 사상형 해조류로 구분되고 기능형군 구성비율은 해조류 서식 환경상태와 밀접한 관련을 보인다고 알려져 있다(Arévalo et al., 2007; Pinedo et al., 2007; Wells et al., 2007). 고흥군 4개 무인도서에서 출현한 해조류 53종을 기능형별로 구분하면, 성긴분기형 50.94%, 엽상형과 사상형이 각 16.98%, 유절산호말형과 각상형이 각 5.66%, 다육질형이 3.77%로 나타났으며 (Song et al., 2011), 본 연구에서는 출현종 145종에서 성긴분기형이 58.95%, 사상형이 17.72%, 엽상형이 12.11%, 다육질형이 7.02%, 유절산호말형이 2.63%와 각상형이 1.58%의 순서로 나타났다. 이러한 연구 결과는 남해안에 서식하는 해조류의 기능형 군별 구성비가 성긴분기형, 사상형, 엽상형, 다육질형, 각상형, 유절산호말형의 순서로서 Sohn (1987)의 기록과 유사하였고 교란이나 환경오염 해역에서 번무하는 사상형과 엽상형은 약 30%로서 환경상태는 양호한 편이다.
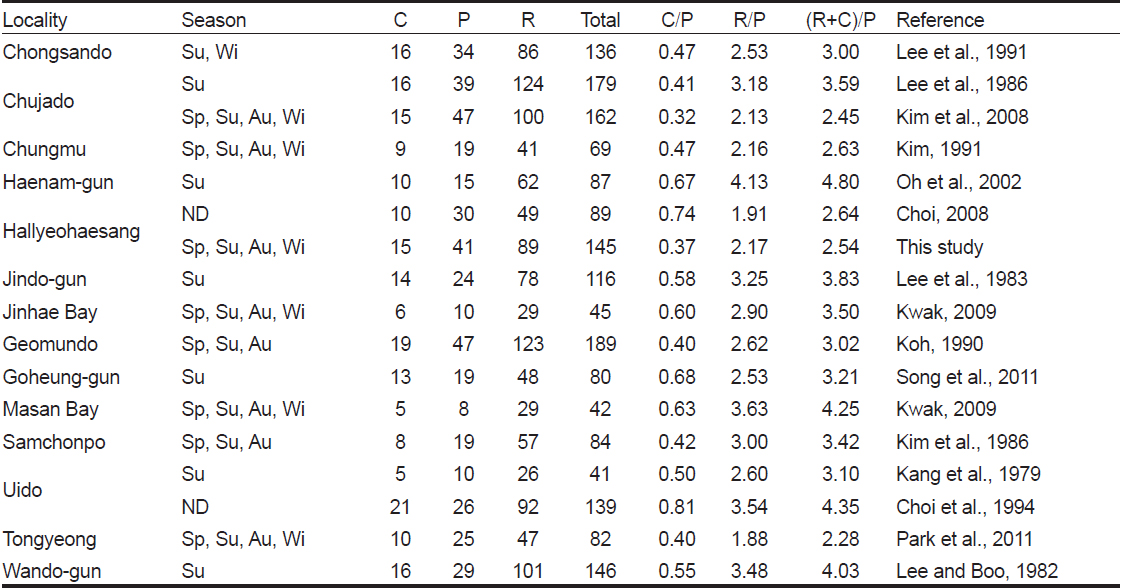
Floristic comparisons of marine seaweeds for several localities in South coast of Korea (Sp, Spring; Su, Summer; Au, Autumn; Wi, Winter)
본 연구에서 한려해상국립공원 내 해조류의 연평균 생물량은 358.00 g dry wt/m2 으로 감암에서 최소(56.51 g dry wt/m2)였고 소매물도에서 847.64 g dry wt/m2 으로 최대였다. 해조류 생물량은 전남 여수시 거문도 인근의 4개 정점(대삼부도, 서도, 동도, 고도)의 3계절(춘계, 하계, 추계) 조사에서 계절별로 83.32-169.16 g dry wt/m2 (평균 121.48 g dry wt/m2)였으며(Koh, 1990), 고흥군 4개의 무인도서(내매물도, 칼도, 아래돔배섬, 진지외도)의 하계 생물량은 21.39-76.22 g dry wt/m2 (평균 50.85 g dry wt/m2)을 보였고, 통영에서 4개 정점(덕포, 가조도, 칠천도, 예포)의 4계절 평균 생물량은 622.61 g wet wt/m2을 보였다(Park et al., 2011). 이외에도, 경남 욕지도 인근 해역에서 계절별로 2년간 수행된 조간대 조사에서 해조류 생물량은 235.30 g wet wt/m2으로(Choi et al., 2008), 금번에 조사된 한려해상국립공원 해역의 해조류 생물량은 다른 해역에 비해 상대적으로 높은 것으로 확인되었다. 선행연구에서 해조류 생물량은 습중량 혹은 건중량으로 기록되어 직접적인 비교는 어렵지만, 건중량(습중량의 1/5)으로 환산하면(Tanigychi, 1998), 한려해상국립공원 내 해조류 생물량은 다른 해역에 비해 높았으며, 추자도(예초, 후포)의 연평균 생물량(425.63 g wet wt/m2)과도 견주어 볼 만 하였다(Kim et al., 2008). 또한, 2005년에 Choi (2008)가 수행한 한려해상국립공원 7개 정점에서 해조류 생물량은 322.60-4,794.00 g wet wt/ m2 (평균 1,727.21 g wet wt/m2)로서 습중량을 건중량으로 환산하여 보면 생물량은 본 연구 결과와 매우 유사하였다.
어떤 해역의 해조상의 특성을 분석하는데 있어서 홍조류와 갈조류의 비인 R/P값을 이용하여 보면 한대와 극지역에서 ≤1.0 이하, 온대에서 1.0-4.0, 열대해역은 ≥4.0 이상을 보였으며(Feldmann, 1937), 녹조류와 갈조류의 비(C/P) 값은 한대해역에서 아열대해역에 걸쳐 0.4-1.5범위의 값을 나타낸다고 하였다(Segawa, 1956). Cheney (1977)는 홍조류와 녹조류의 합과 갈조류의 비인 (R+C)/P값에 따라, 온대성 내지 한대성(< 3), 혼합성(3-6), 열대성(> 6) 해조상의 특징을 나타낸다고 하였다. 본 연구에서는 C/P값이 0.22–0.56, R/P값은 1.74-3.11, Cheney (1977)의 비율인 (R+C)/P값은 1.96-3.67로서 혼합된 해조상을 보였다. 경남 통영의 해조상 연구에서 C/P 값은 0.33-0.55로 한대해역을, R/P값은 1.52-3.27으로 혼합된 해조상을, (R+C)/P값은 1.90-3.82로서 한대에서 혼합성 해조상까지 다양한 형태를 보였다(Park et al., 2011). Choi (2008)의 연구결과, 한려해상국립공원의 24개 정점에서 C/P값은 0.74, R/P값은 1.91, (R+C)/P 값은 2.64로서 C/P값을 제외한 R/P값과 (R+C)/P 값은 본 연구와 차이를 보였는데, 이러한 차이는 연구 정점의 위치와 수 및 채집시기 등의 차이 때문으로 사료된다.
한려해상국립공원 내의 해양서식지 보전 및 가치 증대를 위해서는 생태계의 기초생산자인 해조류의 종조성 및 군집구조에 대한 연구와 관리가 매우 시급한 실정이다. 현재, 남해안은 임해공단 조성 및 항만시설 확충 등의 연안개발로 많은 해역에서 해조류 군락이 황폐화되고 있으며, 종다양성이 감소하고 있다(Lee and Kim, 1977; Koh, 1990; Choi, 1992; Choi, 2008; Ahn et al., 2013). 본 연구 결과는 한려해상국립공원 내 해조류 생물량과 해조류 군집은 남해 연안에서 수행된 선행 연구와 유사하게 나타남으로써(Choi et al., 2008), 아직까지 국립공원 연안의 해조류 군집은 양호한 상태로 평가되지만, 향후 기후변화와 환경오염은 해조류의 종조성과 생물량에 많은 영향을 미칠 것으로 생각된다. 따라서, 우리나라의 대표적인 관광지이고 자연생태계의 보고로 알려진 한려해상국립공원 내에 있는 수많은 도서 및 연안에 서식하는 해조류의 서식처 보호와 보전관리 방안이 마련되어야 할 것이다.