한국 남해 서부해역은 많은 도서로 이루어진 다도해로서 수심이 50 m 내외로 얕은 천해역을 이루고 있고(Jeong, 2001), 참조기, 강달이, 고등어 등의 주요 어장으로 알려져 있으며(Cho, 1982; Cho, 1986; Yang and Cho, 1982), 정치성 어구인 낭장망을 사용하여 주로 멸치를 대상으로 많은 조업이 이루어지고 있다(Kim et al., 2012; Yoo et al., 2014). 또한, 이 해역은 하계에 저수온수가 출현하여 주변 외해역의 고수온수와의 사이에 전선이 형성되는 해양학적 특성을 가지고 있다(Jeong, 2001).
수온변화는 해양생물의 군집구조를 변화시키는 중요한 환경요인이다(Costa, 1990; Tittensor et al., 2010). Stefansdottir et al. (2009)은 아이슬란드 연안역에서 수온 증가에 따른 난류성 어종의 유입을 구체적으로 나타냈고, Sobocinski et al. (2013)은 미국의 체서피크만 주변 해조장에서 수온변화에 따른 어류의 군집구조 변화를 보고하였다. 따라서, 남해 서부해역에서 하계에 출현하는 저수온수의 세력 변화에 따라 서식하는 어류의 생물상에도 변화가 있을 것으로 추정되나, 이를 구체적으로 살펴본 연구 사례는 거의 없다. Yoo et al. (2014)은 완도 연안 낭장망 어획물을 대상으로 2001년과 2013년 간 군집구조를 비교한 결과, 2013년 여름철에 난류성 회유 우점종의 개체수 감소는 수온 상승 둔화에 따른 것으로 보고하였으나, 저수온수 세력 변화에 따른 어류 생물상 변화를 직접적으로 살펴본 것은 아니다.
해양에서 생물다양성(biodiversity)은 생태계의 구조와 기능에 중요한 역할을 수행한다(Johnson et al., 2011). 해양환경 변화에 따른 생물다양성 변화를 밝히는 것은 서식하는 생물의 군집 변화를 예측하고, 이에 따른 해양자원의 보존을 위한 생태계 기반 관리 전략을 수립하는 데 중요하다.
연안 가까이에 설치되는 낭장망에는 4 mm 내외의 세목망이끝자루(cod end) 부분에 부착되므로 멸치 외에 다양한 어종들이 혼획되며(Kim et al., 2012), 낭장망 어획물을 가지고 특정 해역에서 계절별 어류의 생물상 변화 등을 살펴본 연구가 다수 수행된 바 있다(Huh and Kwak, 1998a; Hwang, 1998; Kim et al., 2002; Han and Oh, 2007). 본 연구는 진도 연안 낭장망에 어획되는 어획물을 대상으로 남해 서부해역에서 하계에 출현하는 저수온수의 세력 변화와 이에 따른 연안에 서식하는 어류의 군집구조 변화를 밝히고자 하였다.
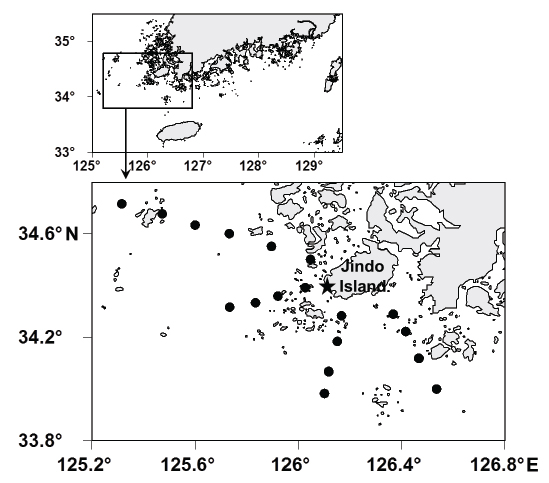
남해 서부해역에서 하계 수온변화를 살펴보기 위해, 2013-2015년간 매년 8월에 총 18개 정점에서 CTD (Conductivity-Temperature-Depth)에 의한 연직수온을 관측하였다(Fig. 1). 또한, 해양수산부에서 매년 발간하는 해양수산통계연보의 전라남도 낭장망 멸치 어획량 자료를 사용하여 남해 서부해역에서 하계 수온변화에 따른 낭장망 멸치 어획량 변화를 살펴보았다. 남해 서부해역에 위치한 진도군과 완도군의 낭장망 허가건수가 전라남도 전체 허가건수의 62.3%를 차지하고 있기 때문에 해양수산통계연보의 어획량 자료를 통해 여름철 남해 서부 해역에서의 낭장망 멸치의 어획량 변동 경향은 파악이 가능할 것으로 간주하였다.
남해 서부 연안에 위치한 진도군 심상리 어장(Fig. 1)에서 조업 중인 낭장망 중 하나를 선정하여 수온조사와 같은 기간 동안 어획물을 매월 5 kg씩 무작위로 표본 채집하였고, 시료의 부패를 막기 위해 드라이아이스와 함께 밀봉하여 연구실까지 운반하였다. 채집한 어류의 동정은 Chyung (1977), Kim et al. (2005) 및 Kim et al. (2005)을 참고하였고, 종 수준에서 개체수 및 중량을 측정하였다.
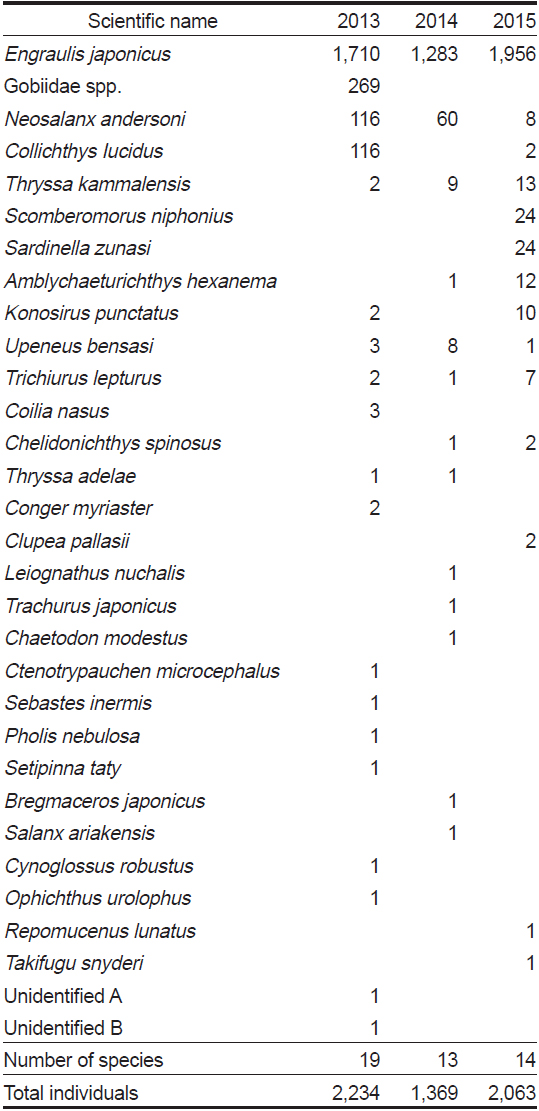
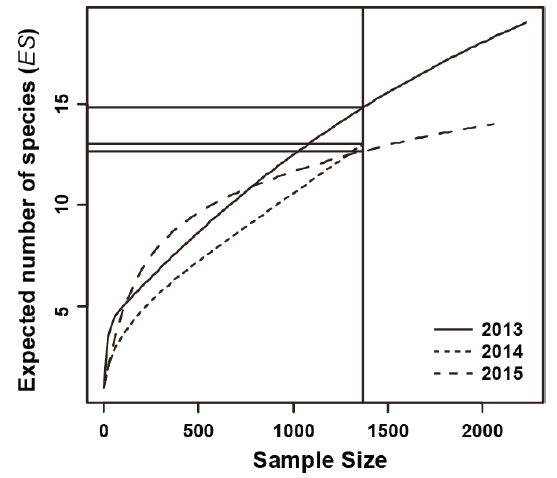
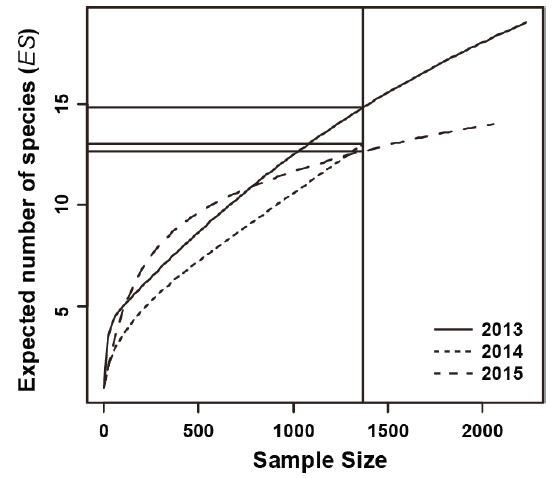
조사기간 동안 채집된 어류의 종조성을 보면(Table 1), 채집개체수가 1-2마리 뿐인 희박종(rare species)이 많이 출현하였다(Pabis et al., 2015; Rossi, 2011). 본 연구에서는 희박종의 출현 변화에 덜 민감하게 반응하고(Smith and Grassle, 1977), 출현개체수 크기에 독립적인 종 풍부도 지수로 알려진 Rarefaction curve에 의한 출현종수의 기대값(
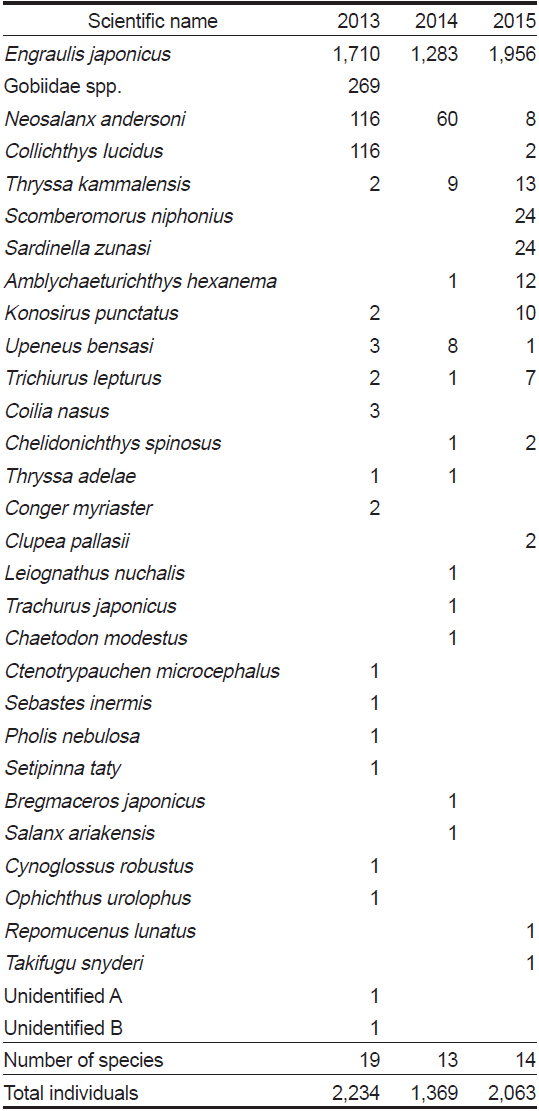
Species composition of fishes caught by gape net with wings in the coastal waters of Jindo Island each August between 2013?2015
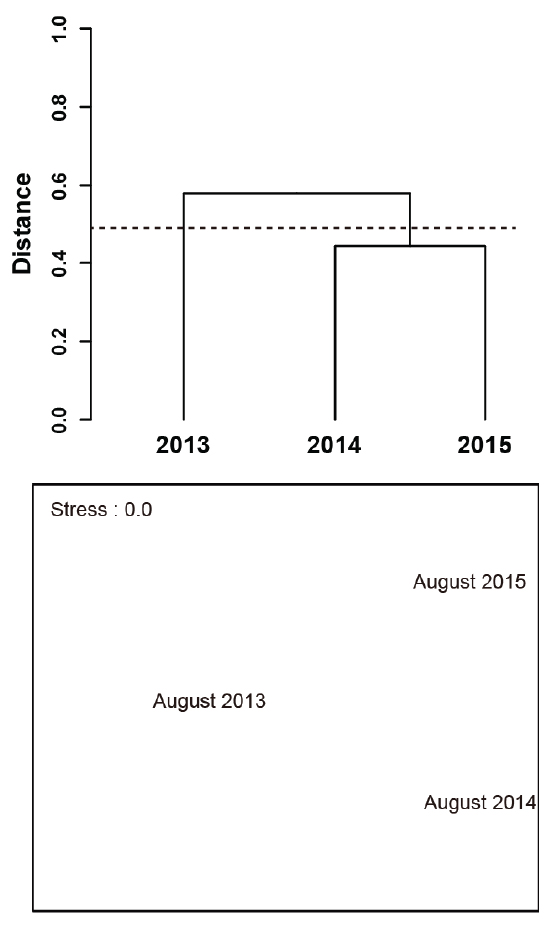
다음으로 연도별 군집 간 유사도를 파악하기 위해 비가중산술평균(UPGMA)에 의하여 군집화하는 계보적 집괴분석 (hierarchical cluster analysis)과 nMDS (non-metric multidimensional scaling)을 수행하였다. 거리 산출에 사용된 유사도 지수는 희박종이 많이 출현하였을 경우 유효한 것으로 알려진 Chao-Jaccard 유사도지수를 사용하였고(Doi and Okamura, 2011; Chao et al., 2006), 소수 출현 분류군에 의한 유사도지수의 과대평가를 낮추기 위해 로그 변환한 개체수 자료를 이용하였다. 또한, 비모수분산분석인 PERMANOVA (Permutational multivariate analysis of variance)을 통해 연도별 군집 간 유의한 차이를 검증하였다(Anderson, 2001; Harding et al., 2011). 위의 모든 분석에는 R 통계에서 제공하고 있는 vegan 패키지를 사용하였다(Oksanen et al., 2014).
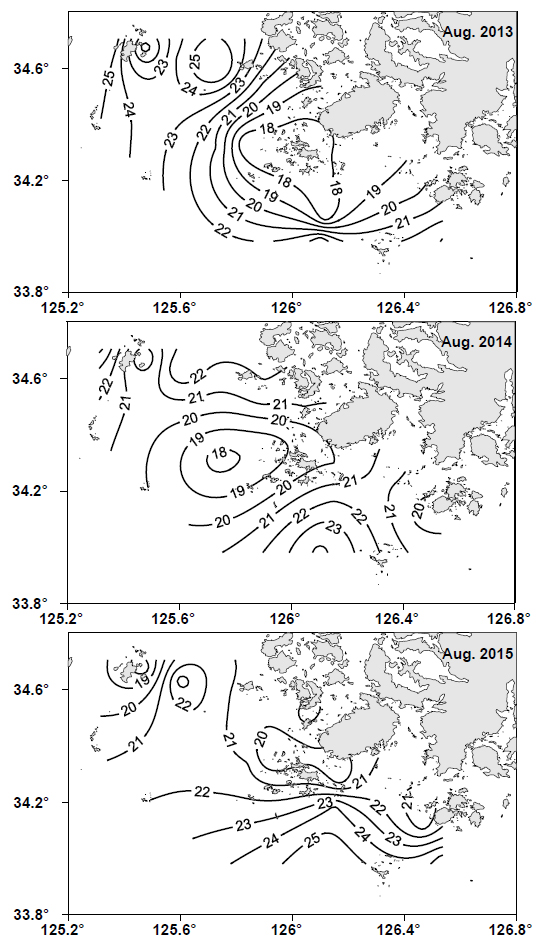
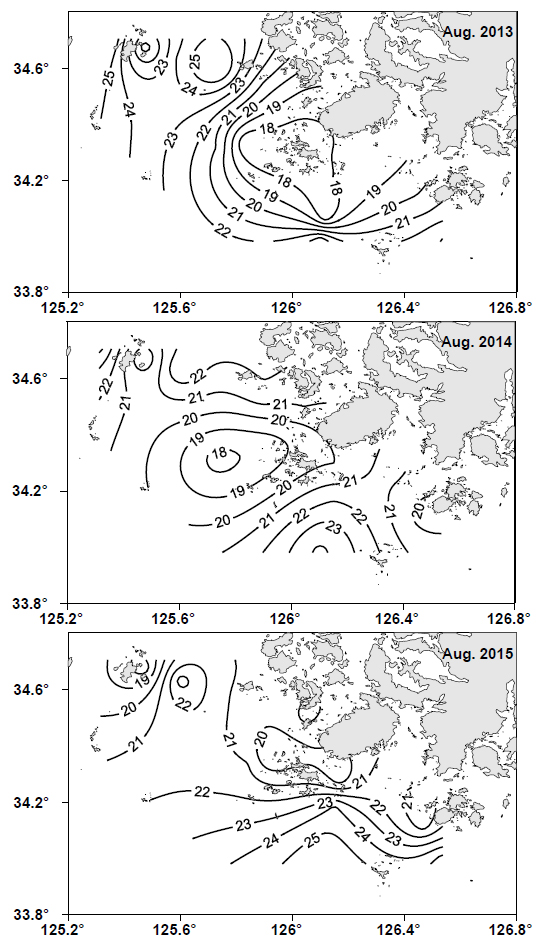
2013-2015년간 매년 8월에 남해 서부 해역에서 관측한 표면 수온의 수평분포도를 Fig. 2에 나타냈다. 2013년 8월에는 19℃ 이하의 저수온수가 조사해역의 중심부에 분포하였고, 이 저수온수는 주변과 비교적 강한 수온전선을 형성하면서 진도 남서쪽 연안을 뒤덮고 있었다. 2014년 8월에도 조사해역의 중심부에 19℃ 이하의 저수온수가 출현하였으나, 그 세력은 2013년에 비해 약하여 진도 연안까지는 미치지 못하였고, 진도 연안의 표면 수온대는 20-21℃ 이었다. 2015년 8월에는 19℃ 이하의 저수온수가 진도 북서쪽 외해의 가장자리 끝에 일부 관찰되었고, 진도 남쪽 외해로부터 24℃ 이상의 상대적으로 따뜻한 해수가 연안으로 유입하고 있었다. 이들 해역을 제외한 나머지 해역에서의 표면수온은 대부분 20-23℃ 이었다.
본 연구에서는 위에 기술한 것처럼 표면수온이 가장 낮았던 2013년 8월을 기준으로 저수온수의 중심수온을 편의상 19℃ 이하로 보고, 상대적인 수온차이를 살펴보았다. 이를 통해 본 연구에서는 2013년 8월은 저수온수 세력이 강했던 기간이었고, 2014년 8월은 2013년에 비해 저수온수가 약하게 존재했던 기간이었고, 2015년 8월은 저수온수 세력이 매우 약했던 기간으로 판단하였다.
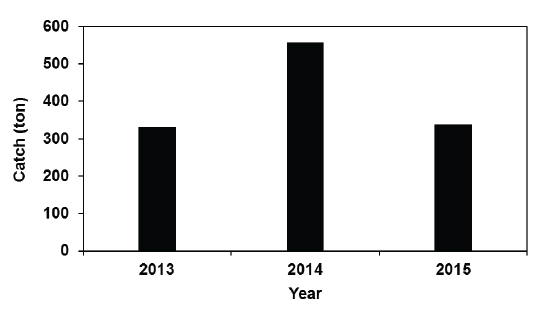
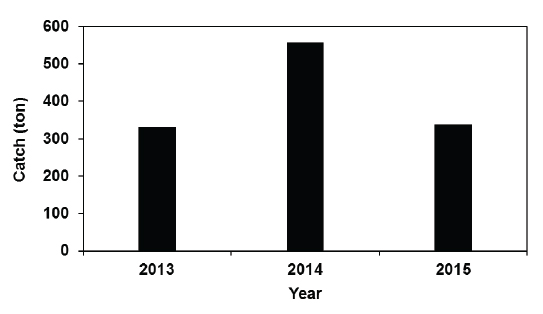
다음으로 2013-2015년간 여름철(7-8월) 전라남도 낭장망 멸치의 평균 어획량 변동을 보면, 2013년에는332톤, 2014년에는 558톤, 2015년에는 338톤이었다(Fig. 2). 즉, 남해 서부해역에서 하계에 저수온수 세력이 강했던 2013년에 낭장망 멸치 어획량은 가장 적었다.
조사기간 동안 진도 연안 낭장망에서 총 31개 분류군에 총 5,666 개체의 어류가 채집되었다. 2013년 8월에는 19종 2,234 개체의 어류가 출현하였고, 2014년에는 13종, 1,369 개체의 어류가 출현하였고, 2015년에는 14종, 2,063 개체의 어류가 출현하였다(Table 1). 우점종 변화를 살펴보면, 멸치(
연도별 Rarefaction curve을 Fig. 4에 나타냈다. 1,369 개체(2014년 8월 총 출현개체수)가 동일하게 관찰되었다고 했을 때, 출현종수의 기대값(
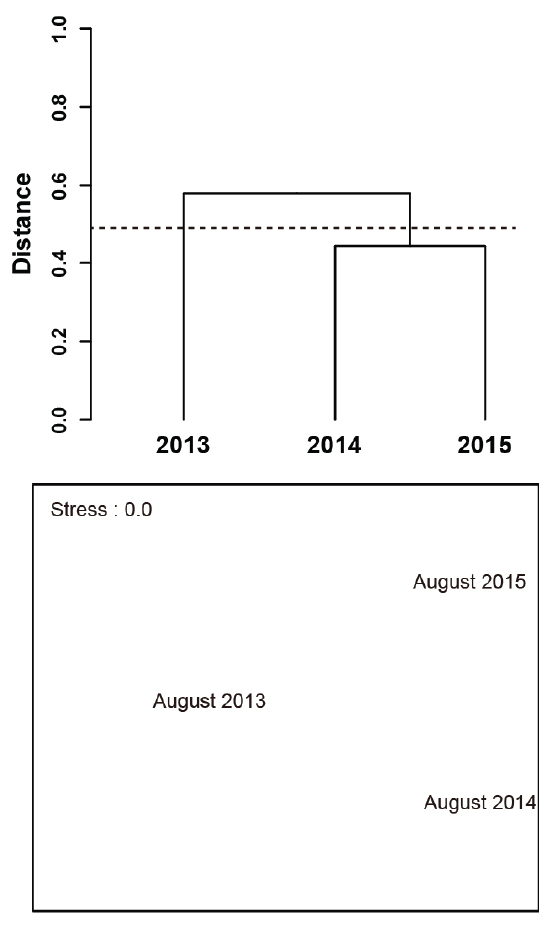
집괴분석을 이용하여 조사기간 동안 연도별 유사성을 살펴보면(Fig. 5), 유사도지수에 의한 거리 0.5 수준에서 저수온수 세력이 강했던 2013년 8월을 하나의 그룹으로, 저수온수 세력이 약했던 2014년 8월과 2015년 8월을 또 하나의 그룹으로 나눌 수 있었다. 그러나 nMDS에 의해 2차원 공간에서 연도별 유사성을 보면, 뚜렷한 군집으로 묶어지지는 않았다. 또한, PERMANOVA에 의해 집괴분석으로부터 나누어진 두 개의 그룹(2013년 8월, 2014년 8월 & 2014년 8월) 간 유의한 차이와 연도별 군집 간 유의한 차이를 확인한 결과, 둘 다 95% 신뢰수준에서 유의한 차이는 없었다.
본 연구는 하계에 진도 연안 낭장망 어장에 분포하는 저수온수의 세력 변화와 이에 따른 어류의 종조성 변화 등의 군집구조 변화를 구체적으로 살펴보았다. 저수온수 세력이 강했던(표면수온 19℃ 이하) 2013년 하계의 종 풍부도는 저수온수가 약했던(표면수온 20℃ 이상) 2014년 8월과 2015년 8월보다 약간 높았다. 특히, 2013년 하계에는 망둑어과(Gobiidae spp.), 황강달이(
2015년 8월의 경우, 삼치(
하지만, 조사기간 동안 연도별 군집 간 유의한 차이를 확인하기 위해 실시한 PERMANOVA 결과, 연도별 군집 간에는 유의한 차가 없었다. 이는 비록 위에 언급한 것처럼 하계에 진도 연안 낭장망 어장에 출현하는 저수온수의 세력 변화에 따른 몇몇 어종의 출현 변화를 확인하였으나, 이것이 연도별 군집구조 간에 유의한 차이를 일으킬 정도는 아니라는 것을 의미한다. 즉, 남해 서부 해역에 출현하는 저수온수는 그 분포범위가 60-90 km이고(Jeong et al., 2009), 7월 말-8월 초경에 가장 발달한다(Yun et al., 2005). 이와 같이 짧은 기간 동안에 진도 연안에 출현하는 소규모(> 100 km 이하) 저수온수가 진도 연안에 서식하는 어류의 군집구조 전체를 크게 변화시키기에는 한계가 있을 것이다.
본 연구에서는 남해 서부해역에서 하계에 출현하는 저수온수의 세력 변화에 따른 낭장망 멸치 어획량 변동도 살펴보았는데, 저수온수 세력이 강했던 2013년에 비해 저수온수 세력이 약했던 2014-2015년에 낭장망 멸치 어획량이 많아지는 경향을 보였다(Fig. 2). Kim et al. (2002)도 1999-2001년간 완도군 수협의 낭장망 멸치 위판량 자료를 사용하여 완도 연안에서 하계에 상대적으로 수온이 낮았던 2000년에 어획량이 가장 적었음을 보였고, 이 연구에서 언급한 것처럼 수온과 멸치 어획량 간에는 상관관계가 있는 것으로 본 연구의 결과에서도 판단된다.
한편, 낭장망은 조류에 의해 휩쓸려 들어오는 어류를 수동적으로 어획하는 어구이므로 유영능력이 큰 어류들의 채집에는 한계가 있고(Huh and Kwak, 1998b), 낭장망에 어획된 어획물이 남해 서부해역 전체를 대표하고 있다고 보기 힘들다. 또한, 본 연구에서 조사된 종조성 자료를 보면(Table 1), 채집 개체수가 1-2마리 뿐인 희박종이 다수 출현하고 있고, 이들 출현이 저수온수의 세력 변화에 의한 것인지 판단하기 힘들었다. 한 해역에서 서식하는 생물상을 정확히 파악하기 위해서는 여러 종류의 채집도구를 동시에 사용하는 것이 바람직하다(Huh and Kwak, 1998b). 따라서, 남해 서부해역에서 저수온수의 세력 변화가 서식 어류의 군집구조 변화에 미치는 영향을 보다 정확히 파악하기 위해서는 향후 수동어구 외에 저인망과 같은 능동어구를 이용한 생물 채집이 필요할 것으로 판단되나, 여러 어구들을 동시에 사용하는 데에는 경비가 많이 소요된다는 해결해야할 문제점이 있다.
또한, Jeong (2001)은 남해 서부해역에서 하계에 출현하는 저수온수는 저층의 저수온수가 강한 조석혼합에 의해 상하 연직 혼합함으로써 나타난다고 설명하였으나, 저수온수 출현에 주기적인 경년 변화가 있는지, 하계에 황해 중앙골을 따라 남하하는 황해저층냉수와의 관계(Jang et al., 2011), 저수온수의 명확한 기준 등 불명확한 점이 많다. 수온변화가 생물의 군집구조 변화를 유발시키는 중요한 환경요인이므로(Tittensor et al.,2010), 남해 서부해역에서 생물의 군집구조 변화를 예측하기 위해서는 저수온수의 출현에 대한 장기적인 모니터링과 함께 보다 명확한 물리적 기작은 구명되어져야 할 것이다.