체르노빌 방사능 누출사고와 일본의 후쿠시마 원전사고 등 최근 큰 방사능 유출 사건이 일어났다. 이러한 이유로 원자력 발전소를 세우거나 방사능을 이용한 기술은 국민들에게 인식이 좋지 않다. 하지만 방사선 기술은 많은 산업분야에서 활발히 이용하고 있고 안전성도 입증되고 있다. 저선량 방사선 처리 기술개발은 생물자원의 생산성 향상, 식량 공급의 안정화, 식물의 환경재해 저항성 증가 등 생물활성의 긍정적 효과를 유도할 수 있는 hormesis 연구법으로 접근하고 있다(Lee et al., 2002). Luckey (1980), Miller and Miller (1987)에 의하면 방사선의 한 종류인 감마선에 의한 hormesis 영향으로는 발아와 출아 및 생장과 발육 촉진, 수량 구성요소가 증가된 것으로 보고 하고 있다. 특히, 식물인 배추, 고추, 참박의 경우 적정선량은 다르지만 초기생육과 수량이 증가 되었다고 보고되었다(Kim et al., 2000a; Lee et al., 1998). 또한, Kim et al. (2007)에 의하면 해양동물인 참전복(
감마선 조사에 의한 생리학적 변화에 관한 실험에 사용된 전복은 제주시 소재의 개인 양식장에서 구입하여 사용하였으며 둥근전복의 경우 평균 각장 8.86±0.06 cm, 각폭 6.01±0.11 cm, 전중량103.07±2.26 g의 개체를 사용하였다. 실험 전복은 감마선 조사 전 1주 동안 수온 23±0.3℃, 염분 32±0.2 psu의 수조에서 순치시켰으며 순치기간 중 먹이의 급이는 중단하였다.
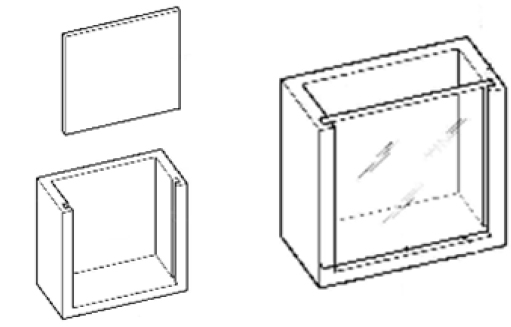
전복은 해양 무척추동물로서 이동성을 지니고 부착생활을 하며 장시간 공기 중에 노출시 폐사하기 때문에, 감마선 조사시 전복의 이동성을 제한하고 전복의 부착 및 탈착 시 스트레스를 최대한 줄이기 위한 전복의 감마선 돌연변이 유발용 케이스를 자체 제작하여 사용하였다. 케이스의 규격은 가로 10 cm, 세로 10 cm, 높이 5 cm의 속이 빈 아크릴 상자로 전복의 부착 및 탈착을 쉽게 하기 위하여 앞면부를 이탈식으로 제작하였다. 이를 위하여 아크릴 박스 양 옆에 홈을 만들어서 앞면판의 이탈을 용이하게 하였다(Fig. 1).
감마선 조사는 제주대학교 원자력과학기술연구소의 협조를 받아 저준위 조사시설(60CO)을 이용하여 실시하였다. 감마선 조사를 위하여 제작된 케이스의 앞면부에 전복의 모패가 부착될 수 있도록 하여 한 마리씩 부착시킨 후 전복의 이동을 제한하기 위하여 고무줄을 이용하여 고정 시켰다. 감마선 조사 실험구는 10, 15, 20 및 25 Gy로 설정하여 조사하였으며, 각 실험구별 암컷 25마리, 수컷 10마리의 전복을 사용하였고 대조구도 암컷 25마리, 수컷 10마리의 전복을 사용하였다. 조사선량은 fricke dosimeter를 이용하여 측정하였고 조사시간은 10 Gy는 18분, 15 Gy는 27분, 20 Gy는 36분, 25 Gy는 45분을 조사하였다. 대조구는 감마선조사 실험구에 비하여 일반적인 스트레스를 제공하기 위하여 17.5℃인 공기 중에 감마선 조사 실험구별 총 합계 시간인 126시간 동안 노출시켜 환경적 스트레스를 제공하였다.
감마선을 조사한 전복의 모패와 대조구 모패를 일주일간 수온 23±0.3℃에서 1주일간 순치 사육하였으나 폐사하는 개체는 발생하지 않아 인공채란을 실시하였다. 인공채란 방법으로는 기온30℃에서 1시간 동안 간출 자극 후 자외선조사해수 자극을 실시하여 모패의 인공산란을 유도하였다. 감마선 조사 선량별 정자와 난자를 수정 시킬 때에는 타 실험구와의 혼합을 방지하기 위하여 선량별로 구간을 설정하여 수정시켰으며, 수정된 수정란은 100 μm 뮬러가제를 사용하여 회수하고 선량별 수정란 개체 수를 확인하였다.
수정률은 선량별로 회수된 수정란 약 20만 개체를 50 mL 유리비커에 옮겨 1 mL를 3반복 계수하여 전체 수정률을 구하였으며 부화율은 선량별로 수정란을 20 L 원형 플라스틱 용기에 수용한 후 부화한 유생과 부화하지 않은 유생을 각각 수거하여 1 L유리비커에 옮겨 1mL를 3반복 계수하여 전체 부화율을 측정하였다.
전복의 수정란은 4 ton FRP (fiber reinforced plastics) (가로 1 m×세로6 m×높이 70 cm) 15개 수조에 각 실험구 별로 3개의 수조에 수정란을 수용하였으며, 사육해수의 수온은 20±0.6℃이었다. 유각이 완전히 형성된 후 에어공급 및 유수식 시스템으로 유생을 사육하였으며, 채묘는 유생을 회수하여 미리 규조류가 배양된 파판에 실시하였다.
선량별로 채묘한 파판(가로 30 cm×세로 30 cm)에서 각각 3개의 파판을 무작위로 선택하여 파판에 부착한 전복을 카운팅하여 부착율을 측정하였으며 전복의 성장율은 부화 후 육안으로 확실히 식별이 가능한 90일 후부터 버니아캘리퍼스를 이용하여 각 실험구별 수조 파판에 부착하여 있는 전복 치패를 30마리를 무작위 선택하여 크기를 측정하였다.
감마선 조사 선량 25 Gy를 조사한 실험구에서 생산된 치패는 양성하는 과정에서 여름철 고수온기에 대량으로 폐사하여 25 Gy를 제외한 나머지 실험구에서 면역활성 변화를 측정하였다. 면역활성 변화는 선량별로 생산된 치패 중 크기가 비슷한 치패를 각각 30마리씩 무작위로 선별하였고 치패의 평균 각장은 1.98±0.08 cm, 평균 각폭 1.02±0.05 cm, 전중량 0.79±0.03 g인 개체를 사용하였으며, 면역활성의 증가 여부를 판단하기 위하여 본 연구진에서는 사전 연구된 30℃에서12 h 째 100%의 폐사를 일으켰던 폐사 수온인 30℃를 이용하여 급격한 수온 변화 0 및 12 h째 각각 5마리의 전복을 채집하여 lysozyme의 활성 변화를 평가하였다. 전복의 심장 옆 혈관으로부터 바늘규격 26 GX 1/2인 1 mL 주사기를 이용하여 혈액을 채취한 후 0.9% 생리식염수로 10배 희석하였다. 혈청 내 lysozyme 활성은turbimetric assay로 다음과 같은 방법에 따라 분석하였다.
모든 시험은 3회 반복으로 실시하였고, 각 실험에서 얻어진 자료에 대한 값의 유의차 유무는,
감마선 조사에 의한 모패는 순치기간 및 채란시기까지 20 Gy 및 25 Gy 실험구에서 다소 활동성이 떨어지는 모습이 관찰되었으나 모든 실험구에서 100%의 생존률을 나타내었고 채란시기에는 선량별로 종묘생산을 추진하여 산란한 난을 각각 분리하여 수정시켰다.
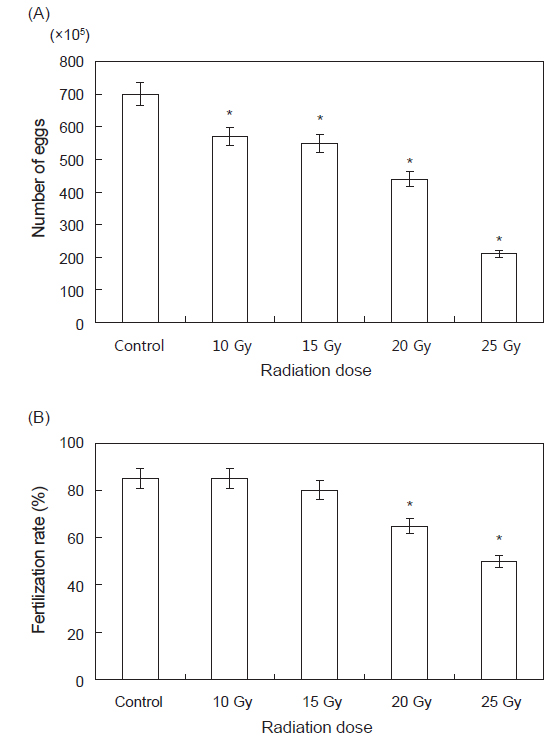
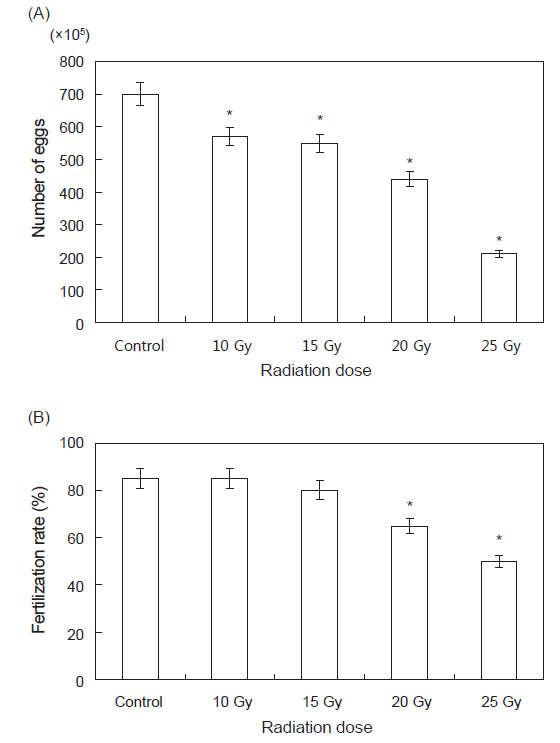
각 실험구별로 산란한 난의 수를 1 mL 피펫을 이용하여 광학현미경(Nikon PROFILE PROJECTOR V-12B)으로 측정한 결과는 Fig. 2 와 같다. Control 실험구에서는 7,000,000±120,000 개체, 10 Gy 실험구에서는 5,700,000±150,000개체, 15 Gy 실험구에서는 5,500,000±110,000 개체, 20 Gy 실험구에서는 4,400,000±100,000 개체, 25 Gy실험구에서는 2,100,000±80,000 개체가 확인되었다.
그 중 정확한 결과를 도출하기 위해 각 실험구마다 약20만 개체를 수거하여 수정된 것과 수정되지 않은 것을 계수하여 방사선 조사에 따른 수정란과, 수정률을 각각 조사한 결과 control 실험구에서는 170,000±110 개체, 10 Gy 실험구에서는 170,000±150개체, 15 Gy실험구에서는 160,000±130개체, 20 Gy실험구에서는120,000±130개체, 25 Gy에서는 100,000±100개체의 수정란이 확인되었다. 수정률은control과 10 Gy에서 85%, 15 Gy에서는 80%, 20 Gy에서는 65%, 25 Gy에서는 50%의 수정률이 확인되었다. 평균 수온 20℃에서 수정부터 부화까지는 11±1시간, 유각 형성까지는 약 23±1시간, 채묘 단계까지는 70±1시간이 소요되었으며 각 실험구별로 유의적인 차이는 없었다(
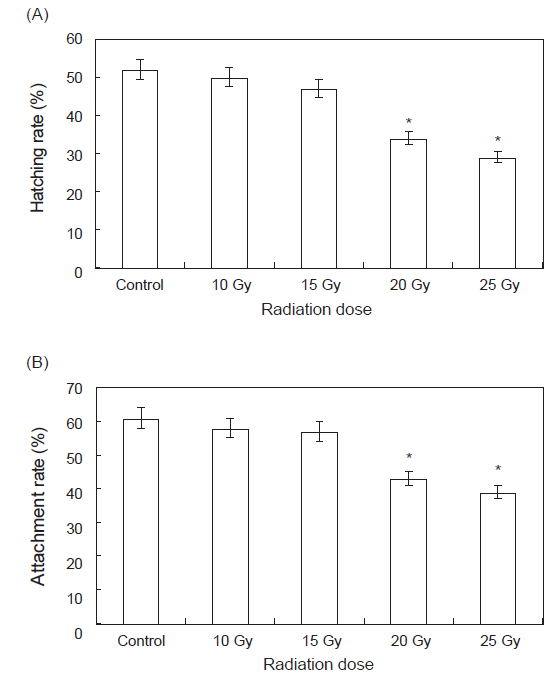
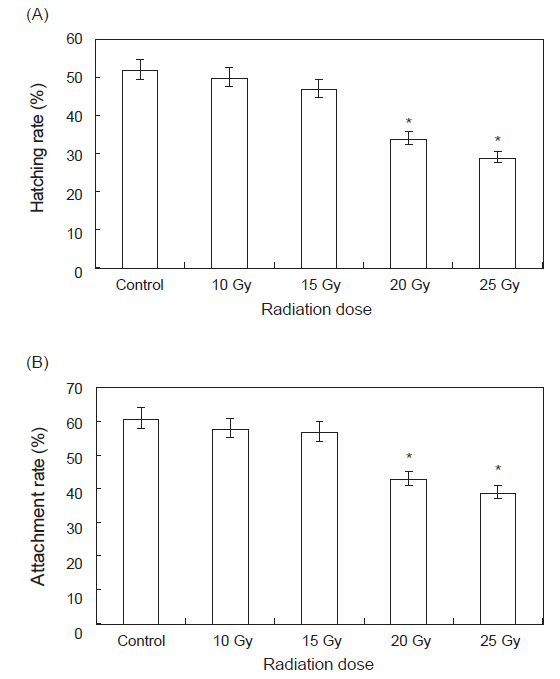
부화율 및 부착률을 측정하기 위하여 각 실험구 별로 세팅된 수조에 수정란을 수용, 사육하며 측정한 결과는 Fig. 3과 같다. 부화율의 경우 대조구, 10 및 15 Gy의 실험구에서는 50±5%의 부화율을 나타내었으나, 20 및 25 Gy 선량의 실험구에서는 30±5%의 부화율을 나타내었다. 부착률의 경우에는 대조구, 10 및 15 Gy의 실험구에서는 60±5%의 부착률을 나타내었으며, 20 및 25 Gy 선량의 실험구에서는 40±5%의 부착률을 나타내었다. 20 및 25 Gy 선량의 실험구에서 수정란의 수와 수정률, 부화율 및 부착률이 낮아 치패의 수가 타 실험구에 비해 적었다(
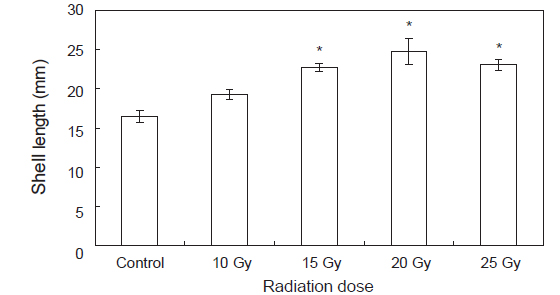
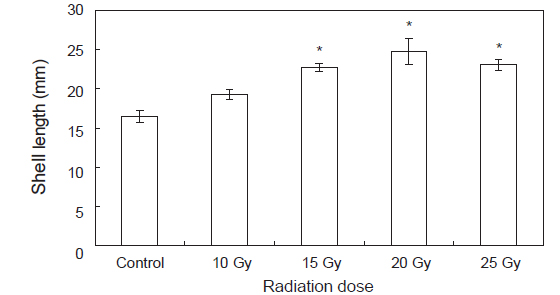
수정란이 부화한 후 유생관리를 통해 미리 규조류를 배양한 파판에 채묘하여 육안으로 확실히 식별이 가능한90일 후 실험구별 수조 파판에 부착하여 있는 전복 치패 30마리씩 무작위로 선택하여 측정한 크기의 결과는 Fig. 4와 같다. 파판은 총 65장으로 파판 1장당 10마리가 부착하여 평균 650±70개체가 부착하였다. 양식장의 사육수온은 평균 25.6℃, 염분은 평균32.6 psu 이었으며, 광주기는 일반 자연조건의 광주기로 10L:14D이었다. 대조구의 경우 17.1±0.6 mm성장하였고10 Gy의 실험구의 경우 19.9±0.3 mm 성장하였고 15 Gy의 경우23.2±0.8 mm성장하였고 20 Gy의 경우 25.7±0.4 mm 성장하였으며 25 Gy의 경우22.9±0.4 mm 성장하였다(
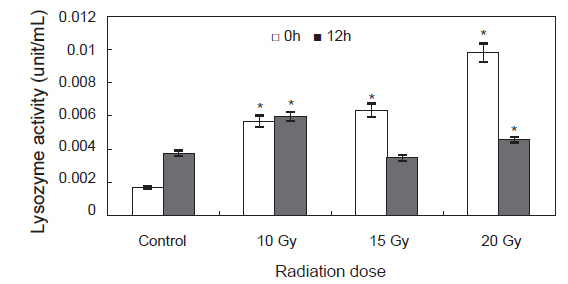
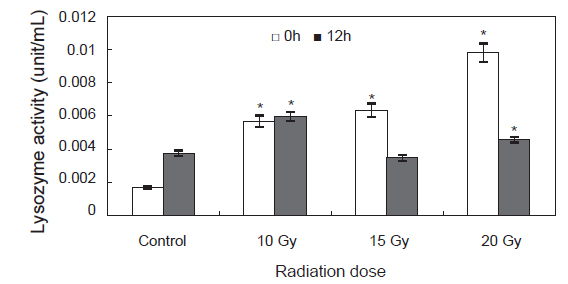
감마선 조사를 한 후 높은 성장률을 가져오는 원인을 규명하기 위하여 고수온인 30℃ 조건하에서 환경 내성을 조사하였다. 환경내성 인자로서 면역기능의 판단 지표로 사용 되는 lysozyme 활성 변화는 Fig. 5에 나타내었다. 0 h에 control에 비하여 다른 실험구에서 유의적인 증가를 나타내었고, 12 h에는 control에 비하여 10, 20 Gy실험구에서는 유의적인 증가를 나타내었고 15 Gy실험구에서는 유의적인 차이가 나타나지 않았다(
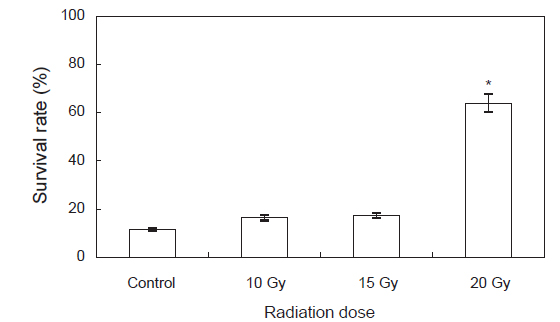
돌연변이 유도를 통한 육종연구는 농업을 중심으로 많은 연구가 되어져 왔으며 그 효과 및 경제적 가치가 매우 높은 것으로 평가되고 있다(Kim et al., 2012; Kim et al., 2013). 본 연구에서는 저선량 감마선을 둥근전복 모패에 조사하여 육종학적 가치에 대한 평가를 위하여 방사선 조사에 의해 생산된 차세대 둥근 전복 치패의 생리적인 변화와 사육을 통하여 성장을 관찰하였다. 일반적으로 생물체에 감마선 조사를 실시할 경우, 세포내 물 분자의 수산화기 과다생산에 의한 세포의 산화적 스트레스 발생이 유발되는 것으로 보고하고 있다(Dubner et al., 1995). 본 연구 결과에서는, 감마선 조사 후 고농도의 감마선 조사 구간인 20 Gy 및 25 Gy 실험구에서는 모패의 폐사는 일어나지 않았으나 활동성이 저하되는 현상이 나타났다. 따라서 본 연구에서 설정한 감마선의 선량은 모패에 대한 스트레스 유발은 일부 나타날 수 있으나 번식에는 영향을 미치지 않아 돌연변이 유도에 적합한 선량으로 판단된다.
감마선 조사된 모패로부터 생산된 수정란의 부화율과 부착률은 대조구, 10 및 15 Gy의 실험구에서는 부화율 50±5%, 부착률 60±5%를 나타내었으며, 20 및 25 Gy 선량의 실험구에서는 부화율 30±5%, 부착률 40±5%를 나타내었다. 20 및 25 Gy 선량의 실험구에서는 수정란의 수와 수정률, 부화율 및 부착률이 낮아 치패의 수가 타 실험구에 비해 적어지는 결과를 나타내었다. Kim et al. (1996) 따르면 감마선 조사는 어린 개체일수록 DNA 및 RNA 손상을 강하게 유발시켜 생리적 장애를 유발하는 것으로 보고하고 있다. 본 연구결과에서 나타난 고선량에서의 부화율과 부착률의 감소 또한 모패의 체내에 존재한 난자에 대한 유전자 손상 유발에 의해 농도 의존적으로 나타난 결과로 추정된다.
한편, Lee et al. (1998)은 저선량의 감마선을 고추 작물의 종자에 조사하여 각 선량별로 나타나는 hormesis 효과를 조사한 결과 대조구에 비해 저선량에 감마선 조사구에서 발아율이 증가되는 것을 관찰하였고, 생육 양상 등에 영향을 미치는 적정선량을 1 Gy에서 20 Gy 사이임을 보고 하였다. Kim et al. (2000b, 2000a, 1999a)도 종자에 저선량 감마선을 조사하여 자란 옥수수, 참박, 배추등의 생육이나 수량이 증가하는 것을 관찰하였으며, Koepp and Kramer (1981)는 저선량 감마선이 조사된 옥수수의 발아율이나 생육의 증가는 종자의 종피를 투과한 저선량 감마선이 지베렐린이나 앱시스산 등의 식물 호르몬의 양적평형에 영향을 미쳐 발아율 및 유식물의 생장을 촉진시키고 결국에는 광합성 기구 및 순 광합성양 환경 스트레스에 대한 저항성을 증가 시킨다고 보고하였다. 본 연구진에서 연구된 참전복의 성장에 대한 감마선의 영향에 대한 hormesis 효과도 30 Gy 미만의 선량에서 발생하는 것으로 보고 하였다(Kim et al., 2007). 또한 본 연구에서도 둥근전복에 저선량 감마선 조사시 비록 부화율과 부착률은 낮게 나타났지만 성장에서는 유의적으로 높게 나타났으며 이는 육상식물과 유사하게 감마선 조사가 호르몬계의 성장 및 생육과 관련된 인자에 영향을 미치기 때문이라 여겨진다. Kim et al. (2011)에 의하면 참돔과 감성돔의 교잡에 의해 생산된 F1을 이용하여 성장속도 및 수온변화와 저염분 환경에서의 내성을 측정한 결과 성장이 빠른 개체들은 환경변화에 대한 내성이 강하다고 보고하였다. 무척추 동물의 경우 면역활성의 지표로 lysozyme이 자주 사용되어 지고 있으며(Wang et al., 2012), 살균 및 항 염증성 성질을 지니고 있어 옵소닌 및 항 기생충 작용, 항 바이러스 작용 및 오염물질의 작용 등에 관여함으로서 생체의 방어기작에 다양한 역할을 담당하는 것으로 알려져 있다(Mohandas, 1985). 이번 실험에서 고수온 스트레스에 대한 lysozyme 활성 변화는 0 h에 control에 비하여 감마선 조사 실험구에서 높게 나타났으며, 특히 20 Gy실험구에서 유의적으로 높게 나타났다. 12 h에는 15 Gy 실험구를 제외한 10 Gy 실험구, 20 Gy실험구에서control에 비해 유의적으로 높게 나타났다. 이러한 결과를 보았을 때 고수온 스트레스에 의해 감마선을 조사하지 않은 control 실험구에서는 lysozyme활성이 낮은 반면, 감마선을 조사한 개체에서는 control 실험구에 비해 lysozyme활성이 높게 나타나 외부의 스트레스에 대한 생체내 방어체계가 작동하여 유해한 활성산소를 제거 함으로서 보다 높은 생존률과 빠른 성장률을 나타낸 것으로 여겨진다.
한편Jwa et al. (2009)에서는 저염분 자극에 의한 둥근전복의 lysozyme의 활성을 측정한 결과 본 연구와 마찬가지로 스트레스를 가한 직 후 control에 비하여 다른 실험구에서 lysozyme 활성이 유의적으로 증가하였다가 감소하는 경향을 나타낸 것으로 보고하였다. 이러한 결과는 수온 및 염분스트레스를 가한 직후 초기 면역활성의 증가로 생존율을 향상시키는 것으로 판단되며 내성이 강한 개체들은 초기 면역활성이 증가하였다가 그 후에는 빠르게 안정화되는 것으로 사료된다. 따라서 본 연구에서 무척추 해양생물인 둥근전복을 이용한 감마선 조사가 생리학적으로 미치는 영향을 조사한 결과 기존의 육상식물들에서 발표된 결과들과 같이 둥근전복 역시 저선량 감마선 조사에서 생존률, 성장률 및 면역활성 반응이 높게 나타나는 것으로 확인되었다. 이는 감마선조사 모패에서 hormesis의 효과가 자손에게도 유전적인 영향을 나타낸 것으로 추정된다. 이러한 결과는 해양동물에 있어서도 감마선을 이용한 돌연변이 유도를 통한 새로운 육종기법으로의 도입 가능성을 제시할 수 있을 것으로 기대된다. 하지만 생리학적 지표는 다양한 환경변화, 개체 및 종에 따라서도 매우 민감하게 반응하므로 추후 이에 대한 세부적인 실험을 통하여 그 지표설정을 위한 연구를 실시하고자 한다.