부레를 갖는 어류에 대한 음향산란강도의 약 90-95% 정도는 부레에 의해 생성되고, 그 나머지는 두부(head), 척추(등뼈, backbone), 어육(fresh) 등에 의해 생성된다(Foote, 1980). 그러나, 대서양 고등어(Atlantic mackerel)나 쥐노래미와 같이 부레를 갖지 않는 어류의 경우에 있어서는 두부, 척추, 어육 등과 같은 어류의 생체조직에 의해 음향산란신호가 생성된다(Nesse et al., 2009). 이 때문에 지금까지 어류에 의한 음향산란특성은 주로 부레의 형상, 크기, 체적 및 자세 등과 같은 형태학적 특징에 주목하여 해석되어 왔고, 이것을 토대로 대상 어족생물이나 플랑크톤 등의 자세와 크기, 또한, 어류의 부레 크기 등을 역으로 추정하려는 연구가 활발하게 진행되어 왔다(Stanton et al., 2003; Jaffe, 2006; Jaffe and Roberts, 2011). 만일, 어류의 두부 (head part)나 부레와 같은 내부조직에 대한 음향산란특성을 각각 독립적으로 분리하여 측정할 수 있다면, 이들 각 부분에 대한 산란 기여율을 토대로 더 유효한 어종식별정보를 추출할 수 있을 것이다. 어류의 두부조직은 두개골, 안구, 아가미 등과 같은 기관으로 구성되지만, 특히, 아가미는 해수 중에 분포하는 산소를 혈관을 통해 흡수하는 역할을 하는 점에서 매우 작은 기포(tiny air bubble)가 그 내부에 다량으로 존재할 가능성이 있다. 또한, 아가미 내부에 분포하는 미소한 기포 군에 의한 산란성분이 두개골이나 안구 등에 의한 산란성분과 결합되면 강한 echo 응답특성을 나타낼 것으로 판단된다. 두부에서 아가미를 제거한 경우와 존재하는 경우에 대한 반사강도를 측정, 비교한 연구(Nesse et al., 2009)에 따르면, 아가미가 있는 경우가 없는 경우보다 반사강도가 약 10 dB 정도 더 높았는데, 이것은 아가미에 다량의 공기가 존재한다는 사실을 암시한다. 이 때문에 부레를 갖는 활어를 대상으로 어종식별정보를 추출할 때, 아가미를 포함하는 두부조직은 부레와 함께 매우 중요한 음향산란요소가 된다.
본 연구에서는 이 점에 주목하여 chirp 어군탐지기를 이용하여 말쥐치, 자리돔, 불볼락의 활어에 대한 chirp 산란신호를 수록한 후, 이들 각 어류에 대한 X-ray 영상을 토대로 부레의 형태학적 차이에 기인하는 음향산란강도의 자세 및 주파수 의존성을 분석, 고찰하였다. 또한, 이로부터 추출된 반사강도에 대한 null의 위치 및 간격의 변동패턴을 SINC 모델에 의한 수치 시뮬레이션 결과와 비교, 분석하여 어체(또는 부레)의 등가적인 크기(equivalent scattering size)를 추정하였다. 이와 동시에 본 연구에 사용한 chirp 어군탐지기의 물표식별능력을 파악하기 위해 라텍스(latex) 재질로 만든 단일 및 이중 체임버(chamber)형 인공부레를 대상으로 펄스압축파형을 추출하여 공간 분해능을 추정하였다.
본 연구에서는 Lee et al. (2015)이 구축한 chirp 어군탐지기를 사용하여 말쥐치, 자리돔, 불볼락의 활어와 모형어체 및 인공부레에 대한 음향산란신호를 계측하였다. 실험은 투명 아크릴 수조(L×B×D, 1.8×1.2×1.2 m)에서 송·수신 음향변환기의 음축상 약 1.2 m 거리에 나이론 monofilament (Ø = 0.3 mm)를 사용하여 이들 활어 및 모형어체와 인공부레를 현수시킨 후, 그 깊이와 자세를 정밀하게 조정하면서 수행하였다. chirp echo 응답특성은 ±45° 자세각 범위(+: head up, −: head down)와 95-220 kHz의 주파수 범위에 대하여 +45°, 0°, −45°의 순서에 따라 약 1° 간격으로 정밀 측정하였다. 각 자세각에 대한 chirp 산란신호는 대역필터와 전치증폭기(VP2000, RESON, Denmark)를 통해 필터링 및 증폭하였고, 디지털 오실로스코프(DS1530, EZ, Korea) 및 스펙트럼 분석기(LSA-30, LIG Nex1, Korea)을 사용하여 일차적인 분석을 마친 후, USB 메모리에 수록하여 후일 목적에 따른 분석을 수행하였다.
활어 및 모형어체와 인공부레에 대한 공간 분해능은 chirp 산란신호를 matched filtering, 즉, 펄스압축하여 추정하였다. 이때, 시간영역에 대한 matched filtering은 산란체로부터 얻어진 산란신호

에 의해 구하였는데(Lord, 2000), 여기서,
만일, 어체 내부에 음향 임피던스가 서로 뚜렷하게 다른 두 생체조직이 존재하고 있을 때, 이들 산란요소에 대한 음향중심의 유효간격을

에 의해 추정할 수 있다. 따라서, 이들 두 산란요소에 의한 펄스 압축파형을 식별하여 분해할 수 있는 최소한의 시간차
본 연구에서는 어류에 의한 echo 생성 메커니즘을 정량적으로 분석하기 위해 인공부레를 갖는 모형어체를 제작하고, 이것에 의한 음향산란 특성을 실제의 활어에 의한 그것과 비교, 분석하였다. 본 연구에서 대상으로 한 모형어체는 말쥐치로서, 말쥐치는 부레의 구조가 비교적 단순하고, 두부 중심과 부레 사이의 음향중심간격이 비교적 넓어 SINC 모델에 의한 음향산란특성의 해석이 용이하기 때문이다(Jaffe, 2006). 본 연구에서는 말쥐치의 척추골 및 다른 어체조직은 제외하고, 단지 부레 및 두부에만 초점을 두고 모형어체를 제작하였다. 말쥐치의 몸체(fish body)와 부레 모형은 연질의 우레탄 고무(VytalFlex-60, Smooth-On, USA)로 제작하였고, 또한 두부는 POM 구(sphere)를 산란요소로 대체하였다. 먼저, 몸체는 어육과 음향 임피던스가 유사한 연질의 우레탄 고무의 Liquid part A와 Liquid part B를 상온에서 1:1로 배합하여 미리 제작한 어체의 금형 틀에 부어 넣고 24시간 경화시켜 완성하였다. 모형부레 틀은 말쥐치의 X-ray 영상으로부터 얻은 부레 형상과 크기 데이터를 토대로 우레탄 고무로 제작하였다. 모형어체를 제작할 때, 모형부레 틀 표면에는 이형제를 도포하여 몸체 틀 내부에 삽입하였고, POM 구는 모형부레 틀 중심으로부터 약 65 mm 전방에 삽입하였다. 몸체 내부에 삽입되어 있는 모형부레 틀은 모형어체가 완성 된 후, 몸체 표면을 약 6 mm 정도 절개하여 제거하였고, 그 절개 부위는 다시 우레탄 고무로서 봉합하여 부레 내부에 공기가 충만한 상태가 유지되도록 하였다. 한편, chirp 어군탐지기의 공간 분해능을 추정하는데 사용한 인공부레의 사진은 Fig. 1과 같다. Fig. 1에서 (a)와 (b)는 각각 단일 체임버와 이중 체임버의 인공부레를 나타낸 것으로, 두 부레의 길이는 모두 67 mm로서 서로 같지만, (b)의 이중 체임버의 경우에는 두 체임버의 중심간격, 즉 음향중심간격이 35 mm가 되도록 제작하였다. 따라서, 본 연구에서는 이들 두 체임버로부터 산란되는 산란신호를 측정, 비교함으로써, chirp 어군탐지기의 공간 분해능을 추정하였다.
또한, 단일 체임버 부레는 어체 내부의 각 생체조직으로부터 산란신호가 생성될 때, 각 산란요소의 음향학적 성질이 균일한 경우, 이에 대한 반사강도의 자세각 및 주파수 의존성을 판단하기 위한 것이다. 반면, 이중 체임버 부레는 어체 내부의 각 생체조직 중에서 부레와 아가미를 포함하는 두부가 산란신호의 생성에 주된 역할을 담당할 때, 이들 두 중심적인 산란요소가 echo 생성에 어떻게 기여하는가를 분석, 고찰하기 위한 것이다.
>
chirp 펄스의 echo 응답패턴에서 반사강도의 null 위치 추적
어류의 음향반사강도는 어체의 체장, 음파의 입사방향에 대한 어체의 자세각(


에 의해 추정할 수 있다(Jaffe and Roberts, 2011). 여기서, sinc(x) = sin(πx)/πx이고, sinc 함수에 대한 null의 위치 n은
>
말쥐치 활어와 모형어체에 대한 광대역 음향산란특성의 비교
Lee et al. (2005)이 보고한 말쥐치의 반사강도에 대한 자세각 및 주파수 의존성에 따르면, peak와 null 간격은 입사각 0˚, 즉 수직입사방향을 중심으로 head-up과 head-down의 자세각 방향으로 좌우대칭적인 변동패턴을 나타내었다. 또한, 이들 패턴은 낮은 주파수에서는 반사강도의 peak와 null 간격이 넓고, 주파수가 증가함에 따라 그 간격이 점차 좁아져 가는 응답패턴을 나타내었다. 본 연구에서는 이와 같이 반사강도의 자세각 및 주파수 의존성 패턴에서 peak와 null 간격이 주기적으로, 또한, 대칭적으로 나타내는 원인을 정량적으로 규명하기 위해 모형 말쥐치에 대한 광대역 음향산란특성을 측정, 분석하였다. 실제의 말쥐치 활어와 본 연구에서 제작한 모형어체에 대한 X-ray 영상을 비교한 결과는 Fig. 2와 같다. 실험에 사용한 모형어체는 말쥐치의 부레 크기, 형상, 자세각과 두부에 대한 음향학적 기여도에 주목하여 제작되었고, 음향학적 기여도가 낮다고 판단되는 체형이나 다른 내부조직은 고려하지 않았다. Fig. 2의 모형어체의 자세각에 대한 산란신호의 시간 및 주파수응답특성을 나타낸 결과는 Fig. 3과 같다. 또한, 모형어체와 말쥐치 활어에 대한 산란 맵(map)과 matched filter 출력을 서로 비교, 분석한 결과는 각각 Fig. 4 및 Fig. 5와 같다.
Fig. 2의 모형어체 제작에 사용한 폴리우레탄 고무의 음향 임피던스(Z = ρc, ρ:밀도, c: 음속)는 Z=1.79×106Rayls (ρ=1,150 kg/m3, c=1,560 m/s)로서(Mott et al., 2002), 이 값은 실제 어류의 몸체에 대한 음향 임피던스 Z=1.68×106Rayls (ρ=1,070 kg/m3, c=1,570 m/s)와 거의 유사한 값을 갖는다(Clay and Horne, 1994). 따라서, 20℃에 대한 물의 음향 임피던스 Z= 1.54×106 Rayls (ρ=1,030 kg/m3, c=1,490 m/s)에 대한 실제 어류와 모형어체에 대한 밀도 contrast는 각각 1.04와 1.12이었고, 음속 contrast는 모두 1.05로서 같았다. 이 때문에 모형어체의 음향 임피던스가 실제 활어보다 약 6.5% 정도 컸지만, 이 차이는 본 연구에서 목적하는 산란신호의 자세각 및 주파수 의존성을 평가하는데 있어 큰 영향을 미치지 않는다고 판단하였다. 또한, Fig. 2(a)에 나타낸 말쥐치 활어의 X-ray 영상(Lee et al., 2005)에서 척추골은 다른 어류와 달리 두부 뒤쪽에서 활 모양으로 휘어져 부레를 감싸고 있는데, 어류의 척추골을 포함하는 모든 뼈(bone)의 밀도와 음속 값을 각각 ρ=1,993 kg/m3 (Fassler et al., 2009)와 c=1,560 m/s (Wear et al., 2000)라 가정하면, 음향 임피던스는 Z= 2.96×106Rayls이 된다. 이 값에 대한 밀도 및 음속 contrast는 각각 1.95, 1.05로서, 특히, 음속 contrast는 몸체와 같으나 밀도 contrast는 약 87.5% 정도의 차이를 나타내어 척추를 포함하는 골조직과 몸체의 어육조직 사이의 경계면에서 음향산란이 발생할 것으로 판단된다. 그러나, 실제 이들 음향 임피던스의 차이가 echo 형성에 기여하는 점유율은 부레의 그것과 비교할 때, 극히 작은 수준이다. 즉, 일반적으로 부레의 음향 임피던스는 Z=427.8 Rayls (ρ=1,24 kg/m3, c=345 m/s)로서(Clay and Horne, 1994), 이 값은 어류의 뼈 조직에 대한 음향 임피던스의 1.45×10−4배 정도에 불과하다. 이 때문에 부레와 주변의 어육, 뼈 등과 같은 어체조직 사이에는 매우 큰 음향학적 불연속 경계면이 형성되고, 이들 경계면으로부터 매우 강한 산란신호가 형성된다. 따라서, chirp 펄스의 주파수 대역폭에 의해 결정되는 공간 분해능 영역내에서 부레와 뼈, 어육 등과 같은 내장조직이 함께 존재하는 경우, 부레를 제외한 내장조직에 의한 산란성분은 부레 경계면에서 생성되는 강한 산란성분에 의해 대부분 매스킹(masking) 된다. 또한, 이들에 의한 echo기여도는 부레에 의한 echo 응답신호에 약간의 변동을 초래하는 정도에 불과한 것으로 판단된다.
한편, 말쥐치의 두부는 다른 어류에 비해 몸체에 대한 체적 점유율이 매우 크고, 두개골의 조직이 매우 견고한 형태를 하고 있다. 또한, 두부의 주둥이 쪽을 향한 기울기가 부레의 꼬리 쪽에 대한 자세각과 거의 유사한 형상을 하고 있어 안구 바로 뒤쪽을 기준으로 볼 때, 거의 좌우 대칭적인 형태학적 패턴을 나타낸다. 이 때문에 이들 두 요소에 의한 산란성분은 서로 복잡하게 보강 및 상쇄 간섭을 일으킬 것으로 판단된다. 본 연구에서는 어류의 뼈와 음향 임피던스가 유사한 직경 12.8 mm의 폴리아세탈(polyacetal, POM-C) 구를 폴리우레탄 고무로 제작한 모형어체의 두부 쪽에 삽입하여 말쥐치의 두부 조직을 제작하였는데, 이 POM-C의 밀도와 음속은 각각 1,407 kg/m3, 2,486 m/s로서 음향 임피던스는 Z=3.481×106 Rayls이고, 이 재질은 인체의 두부 유사 팬텀(head-mimicking phantom)을 개발하는 데에도 널리 사용되고 있는 물질이다(Min et al., 2014).
Fig. 3에서 (a)는 모형어체에 의한 광대역 산란신호의 시간응답특성이고, (b)는 주파수 응답특성을 나타낸 결과이다. 이들 그림에서 시간응답의 진폭은 0∼1.2 V 범위의 상대레벨로서, 주파수 응답은 −15~+30 dB의 상대레벨로서 나타내었다. Fig. 3(a)의 echo 응답은 head-up 약 40° 부근부터 나타나기 시작하여 0° 부터 head-down의 자세로 접어들면서 강한 응답특성을 나타낸 후, 다시 자세각 15°를 벗어나면서부터는 점차 약해지는 변동특성을 나타내었다. 한편, Fig. 3(b)의 주파수 응답은 head-up 약 30° 부근까지의 자세각에서는 95-170 kHz의 저주파 영역에서 미약한 응답을 나타내었지만, 0°에 접근함에 따라 95-220 kHz의 모든 주파수 범위에서 점차 강해져 0°~15°의 자세각 범위에서 가장 강한 응답특성을 나타내었고, 그 이후에는 다시 약해지는 변동패턴을 나타내었다. 한편, 주파수 응답특성의 −15°~+20°의 범위에서는 170 kHz의 주파수를 전후하여 약간의 다른 응답패턴을 나타내었는데, 이 현상은 말쥐치 두부에 삽입한 POM-C 구의 산란신호가 부레에 의한 산란신호에 영향을 미쳐 초래된 현상으로 판단된다. 어류에 의한 음향산란은 서로 인접하고 있는 산란조직 사이의 음향 임피던스의 불연속성에 기인하여 발생하기 때문에 이들 어체 내부의 생체조직, 특히, 부레나 뼈와 같은 산란요소 사이, 또한, 해수와 몸체 사이의 밀도와 음속 contrast의 변화율(gradient)에 주목할 필요가 있다. 그러나, 모형어체 내부에 말쥐치 부레와 유사한 크기 및 형상의 인공부레가 존재하고, 그 전방 50 mm의 위치에 POM-C 구가 존재함에도 불구하고, 이와 같은 응답특성을 보인 것은 이들 두 요소에 의한 산란신호가 서로 간섭하고 있음을 의미한다. 따라서, 실제의 어류에 있어서는 내부조직 사이의 간섭이 매우 복잡하게 발생할 것으로 예견되기 때문에 산란요소 상호간의 상쇄 및 보강간섭을 일으키는 메커니즘을 정량적으로 이해해야만 어종 고유의 식별정보를 추출할 수 있을 것으로 판단된다.
한편, Fig. 4는 말쥐치 활어와 모형 말쥐치에 대한 산란진폭의 자세각 및 주파수 의존성을 나타낸 결과인데, (a)는 말쥐치 활어에 대한 산란진폭의 echogram이고, (b)는 모형 말쥐치에 대한 산란진폭의 echogram으로서 이들 그림에서 진폭레벨은 0~1 V의 상대진폭으로 나타내었고, 종축은 주파수(kHz)이고, 횡축은 말쥐치의 자세각(°)이다.
Fig. 2의 X-ray 영상을 볼 때, 모형 말쥐치는 실제 말쥐치의 부레 형상, 자세패턴, 크기 등과 거의 유사한 공기 체임버를 체내에 갖고 있고, 또한, 이 체임버는 두부에 위치하는 POM-C 구와 서로 음향학적으로 결합된 구조를 하고 있다. 따라서, 모형어체에 의한 echo 응답특성이 실제의 말쥐치에 의한 echo 응답특성과 유사하다면, 말쥐치로부터 음향산란신호는 주로 부레와 두부의 생체조직에 기인하는 것으로 판단할 수 있을 것이다. Fig. 4(a)의 말쥐치 활어에 대한 산란진폭의 자세각 및 주파수 의존성의 경우, 수직 입사각 0°를 중심으로 산란진폭의 peak, 또는 null이 좌우대칭으로 나타나면서 그 변동은 주파수가 증가함에 따라 peak 간격, 또는 null 간격이 점차 좁아져 가는 변동패턴을 나타내었다. Fig. 4(b)의 모형 말쥐치에 대한 산란진폭의 자세각 및 주파수 응답특성은 기본적으로 Fig. 4(a)의 말쥐치 활어에 대한 응답특성과 매우 유사한 변동패턴을 나타내고 있지만, 모형어체에 있어서는 95-130 kHz 주파수 범위에 있어서 peak 및 null 간격이 말쥐치 활어의 경우보다 더 좁은 경향을 나타내었고, 또한, 170 kHz 부근에서는 echo 응답의 변동패턴이 완만하게 역전되는 현상을 나타내었다. 또한, 모형어체에서는 head-down의 자세각 영역에서 산란진폭에 대한 peak와 null의 주기성 및 규칙성이 강하게 나타난 반면, 말쥐치 활어의 경우에 있어서는 head-up 자세각 영역에서 강하게 나타났다. 이와 같은 현상은 모형어체 내부에 척추골이 존재하지 않는다는 점에 주목해 볼 때, 이와 밀접한 관련이 있는 것으로 판단된다.
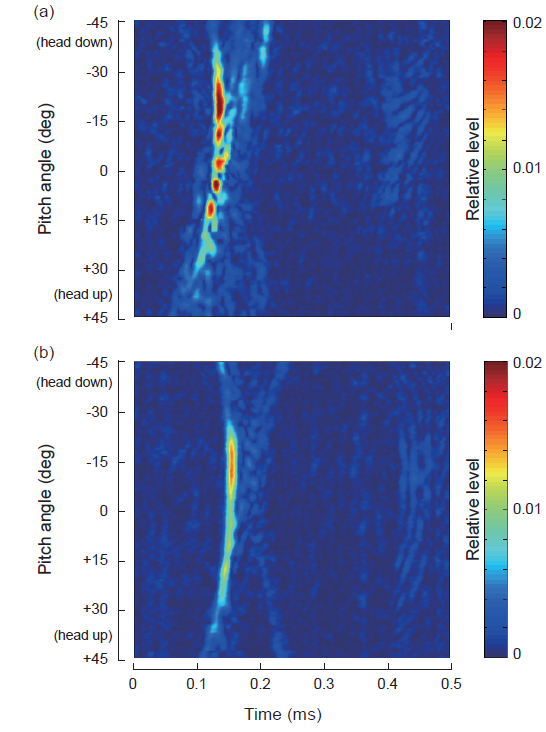
따라서, 본 연구에서는 어류로부터 산란신호가 생성될 때, 어체 내부에 존재하는 각 생체조직이 서로 어떻게 기여하는가를 정량적으로 고찰하기 위해 matched filtering을 통해 얻어진 말쥐치 활어와 모형어체에 대한 펄스압축파형을 서로 비교, 분석하였는데, 그 결과는 Fig. 5와 같다. Fig. 5의 (a)는 말쥐치 활어에 대한 펄스압축파형의 자세각 의존성이고, (b)는 모형 말쥐치에 대한 펄스압축파형의 자세각 의존성으로서, 이들 그림에서 진폭 레벨은 0∼0.02 V 범위의 상대진폭으로 나타내었고, 종축은 자세각(°)이고, 횡축은 시간(ms)이다. 실제 실험에 사용한 말쥐치의 X-ray 사진으로부터 얻어진 부레의 경사각은 22.3°이고, 모형어체 내부에 존재하는 모형부레의 경사각도 이와 같다. Fig. 5 (a)에서 알 수 있는 바와 같이 말쥐치 활어에 대한 펄스압축파형의 자세각 의존성은 부레의 경사각 부근인 약 21°를 중심으로 14°~30°의 자세각 범위에서 매우 강한 echo 응답을 나타나고 있는 점으로부터 이 산란성분은 주로 부레에 기인하는 것임을 알 수 있었다.
그러나, Fig. 5(b)의 모형어체에 대한 자세각 의존성에 있어서는 약 −15°를 중심으로 −5°~ −25°의 자세각 범위에서 강한 echo 응답특성을 나타내었지만, 부레의 위치각 −22.3°부터는 약간 상이한 응답패턴을 나타내고 있다. 이것은 모형어체에 대한 부레의 체적, 구조 및 형상 등이 실제의 것과 다르기 때문에 발생한 것으로 판단되지만, 보다 정량적인 분석을 위해서는 다양한 구조의 부레를 대상으로 더 많은 실험적 분석을 수행할 필요가 있다고 판단된다.
>
부레의 형태학적 차이에 따른 반사강도의 peak 및 null 패턴의 변화
부레를 갖는 어류의 경우, 산란신호의 생성에 가장 큰 영향을 미치는 기관은 주위의 어체조직이나 매질과의 음향 임피던스의 차가 매우 커서 음향학적으로 불연속 경계면을 형성하는 부레이다. 그러나, 어류는 아가미의 혈관을 통해 각 생체조직에 산소를 공급하므로 아가미 내부에는 호흡시 매우 미소한 기포가 다량으로 발생하게 되는데, 이 때문에 두부조직 역시 매우 중요한 음향학적 산란요소가 된다. 이 때문에 chirp 산란진폭의 자세 및 주파수 의존성을 나타내는 반사강도 패턴을 먼저 작성하고, 또한, 부레와 두부조직이 척추골에 의해 음향학적으로 서로 결합되어 있다고 가정하여 SINC 모델에 의한 산란진폭의 null 패턴을 수치계산을 통해 추출하였다. 그 후, 수치계산의 null 패턴을 반사강도 패턴과 중첩시켜 어체의 등가적인 길이, 즉 등가적인 부레의 크기를 추정하였는데, 그 결과는 Fig. 6과 같다. Fig. 6에서 (a), (b), (c)는 각각 불볼락, 자리돔 및 말쥐치에 대한 반사강도의 자세각 및 주파수 의존성을 나타낸 결과이고, 종축은 자세각(°)이며, 횡축은 주파수(kHz)이다. 또한, 각 어종의 반사 강도의 레벨은 −70~ −30 dB 범위의 컬러 바 스케일로서 나타내었다. Fig. 6에서 실선은 각 어체를 등가적인 길이
따라서, Fig. 6의 echo 응답에 대한 규칙성 및 대칭성을 정량적으로 분석하면 부레의 등가적인 크기를 추정할 수 있다(Jaffe, 2006). 이 점에 주목하여 본 연구에서는 SINC 모델을 토대로 Fig. 6의 echogram에 나타나는 null 패턴을 가장 근사적으로 나타낼 수 있는 부레의 등가적인 크기를 추정하였다. SINC 모델은 기본적으로 어체의 산란성분은 부레를 중심으로 각 어체조직이 선형적으로 결합된 하나의 산란체에 의해 생성된다는 가정하에서 성립한다. 따라서, SINC 모델로부터 추정되는 부레의 등가적인 크기는 어체의 등가적인 체장에 상당하는 치수임에 주목할 필요가 있다. Fig. 6(a)의 불볼락(전장 258 mm, 부레 길이 58 mm)에 대한 반사강도의 자세각 및 주파수 의존성을 살펴볼 때, head-up 약 17.5°를 기준으로 반사강도의 peak 궤적은 주파수가 낮은 영역에서는 peak 진폭의 간격이 넓고, 주파수가 높은 영역으로 갈수록 그 간격이 점차 좁아지는 경향을 나타내었다. 또한, 이와 같이 대칭적이면서도 규칙적으로 나타나는 peak와 null 패턴은 head-down의 자세각 범위에 치우쳐 나타나고 있음을 알 수 있다. 이와 같은 현상은 불볼락의 부레의 형상이 이중 체임버형으로서, 전부와 후부의 부레 길이가 각각 47 mm (경사각 18.2°), 11 mm (경사각 35.9°)로서 서로 다르고, 그 경사각 역시 서로 다른 형태학적 패턴을 나타내고 있는 것과 밀접한 관계가 있는 것으로 판단된다. 반사강도의 변동패턴에 대한 null 위치의 궤적을 SINC 모델에 의해 도출한 궤적과 중첩시켜 고찰해 볼 때, head-down의 자세각 범위에서는 유사한 변동패턴을 나타내었으나, head-up의 자세각 범위에 있어서는 큰 차이를 나타내었다. 이 때문에 여기서는 head-down의 자세각 범위에 대한 null 궤적을 SINC 모델에 의한 null 패턴에 근사시켜 어체의 등가적인 길이를 추정하였는데, 그 길이는 72 mm이었다. 이 값은 불볼락의 전·후부의 총 부레길이 58 mm와 비교하여 약 24% 큰 값으로서, 이와 같은 차이는 불볼락의 경우, 부레 이외의 다른 어체 조직에 의해서도 비교적 강한 산란신호가 생성된다는 것을 암시하는 것이다. 특히, Fig. 6에서 반사강도에 대한 peak와 null 위치의 궤적변화는 비록 같은 자세각 범위라 할지라도 주파수가 높을수록 더 복잡한 패턴을 나타내었다.
한편, Fig. 6(b)의 자리돔(전장 163 mm, 부레 길이 53 mm)에 대한 반사강도의 자세각 및 주파수 의존성을 살펴볼 때, 반사강도는 head-down 약 3.5°를 기준으로 peak 궤적이 고주파수를 향해 대칭적으로 수렴해 가는 변동패턴을 나타내었다. 즉, 반사강도의 peak와 null 패턴은 head-down의 자세각 범위에 약간 치우쳐 나타나는 경향이 있었다. Fig. 6(b)에 나타낸 반사강도의 echogram에서 null 위치의 궤적을 SINC 모델에 의해 도출한 궤적과 중첩시켜 추정한 어체의 등가적인 길이는 52 mm이었다. 이 값은 자리돔의 부레 길이 53 mm와 비교하여 거의 일치하는 결과로서, 이것은 자리돔으로부터의 산란신호는 거의 전적으로 부레에 의해 생성된다는 것을 암시한다.
Fig. 6(c)의 말쥐치(전장 210 mm, 부레 길이 39 mm)의 경우에 있어서는 반사강도의 자세각 및 주파수 의존성에 대한 null 위치의 측정된 패턴이 SINC 모델에 의한 계산된 패턴과 매우 유사한 경향을 나타내었다. 또한, SINC 모델에 의해 추정한 어체의 등가적인 길이는 77 mm이었다. X-ray 영상으로부터 측정된 말쥐치 부레의 길이는 39 mm, 어체와 부레의 길이비와 체적비는 각각 0.18, 0.021로서, 어체의 등가적인 길이의 추정치는 부레의 실제 길이보다 매우 큰 값을 나타내었다. 일반적으로 말쥐치는 두부의 두개골이 매우 견고하고, 다른 어체 보다 완만하게 기울어져 매우 양호한 음향 산란체로서의 기능을 발휘하는데, 말쥐치의 두부 길이(주둥이 끝에서 안구 뒤쪽까지의 길이)는 68 mm이었고, 주둥이 끝에서 부레 끝단까지의 길이는 약 124 mm이었다. 또한, 두부와 부레 중심 사이의 간격은 약 71 mm로서, 이 길이는 SNC 모델에 의한 어체의 등가적인 길이의 추정치 77 mm와 유사한 값을 나타내었다. 이와 같은 결과로부터 말쥐치의 두부와 부레는 음향산란신호를 생성하는 주된 음향 산란체로서의 역할 및 기능을 발휘하고 있음을 알 수 있었다. Fig. 7(c)의 echogram에서 자세각 0°를 기준으로 head-down, 또는 head-up 방향으로 자세각이 변화할 때, 자세각이 증가함에 따라 반사강도의 peak 진폭이 점차 약해지는 경향을 나타내었다. 이것은 어체 조직에 기인하는 음향산란의 감쇠(damping)현상이 점차 증대되어 진동변위가 감소하였기 때문인 것으로 판단된다. 이와 같은 peak와 null의 간격 및 변동패턴이 산란체의 내부조직의 재질과 관련된 것인가, 또는 산란체 자체의 형상과 관련된 것인가에 대해서는 보다 정량적 분석이 필요하다. 또한, 어류는 척추골을 비롯한 수많은 뼈 조직이 어체 내부에 존재하고, 주위 매질과 이들 사이에는 음속과 밀도 contrast에 차이가 있을 뿐만 아니라 척추 내부에는 미소하지만 cavity가 존재하기 때문에 이들 요소에 대한 영향에 대해서도 정량적인 분석이 필요하다.
>
단일 및 이중 체임버 인공부레의 광대역 음향산란특성과 공간분해능 추정
어류 부레는 어종에 따라 형상, 크기, 분포위치 및 자세각 등과 같은 형태학적 특징이 서로 매우 다르고, 압력의 변화에 반응하여 음향산란특성이 매우 불규칙하고 복잡하게 변동한다. 그 중에서 특히, 부세나 고등어와 같이 부레가 척추골을 따라 길이 방향으로 길게 전개되어 있는 경우에는 두부나 척추골, 또한 어체 근육 등으로부터 생성되는 미약한 산란성분이 부레로부터 생성되는 강한 산란성분에 의해 매스킹되어 거의 식별할 수 없다(Lee et al., 2015). 더욱이, 이들 어종에 있어서는 음파가 부레에 수직 입사하는 자세각 부근의 좁은 범위에서는 매우 강한 echo 응답이 집중적으로 나타나지만, 그 자세각 범위를 벗어나면 echo 응답이 급속히 약화되는 패턴을 나타낸다. 본 연구에서는 이와 같은 현상이 나타나는 echo 생성 매커니즘을 파악하기 위해 두 종류의 모형 부레, 즉 단일 체임버형과 이중 체임버형의 부레를 제작하여 echo 생성패턴을 분석, 고찰하는데, 그 결과는 Fig. 7, Fig. 8 및 Fig. 9와 같다. 여기서 이중 체임버의 인공부레는 Fig. 1(b)에서 알 수 있는 바와 같이 앞쪽의 부레 체임버(길이 43 mm)가 뒤쪽의 부레 체임버(길이 24 mm) 보다 19 mm 긴 형상을 하고 있음에 주목할 필요가 있다.
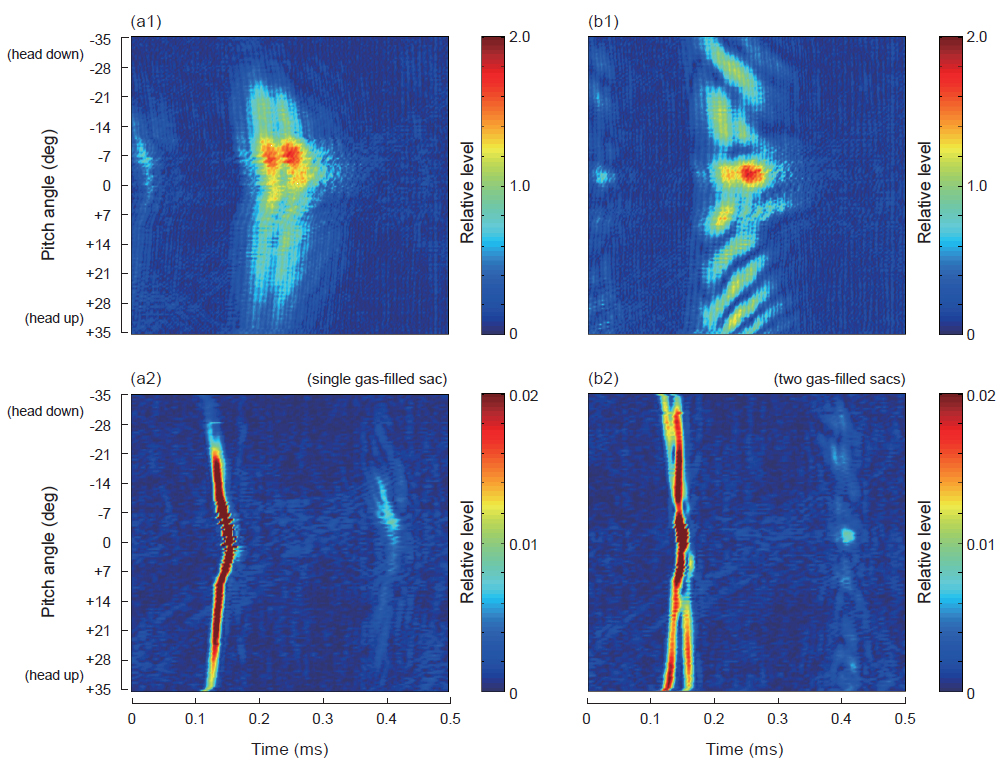
Fig. 7에서 (a1)과 (b1)는 각각 단일 및 이중 체임버를 갖는 인공부레에 대한 산란신호의 시간응답특성을 나타낸 결과이고, (a2)와 (b2)는 이들 산란신호를 matched filtering하여 얻은 압축펄스파형의 echogram이다. Fig. 7에서 산란신호의 시간응답은 0~2 V 범위의 상대진폭으로, 또한, 압축펄스파형의 echogram은 0~0.02 V 범위의 상대진폭으로 각각 나타내었다. Fig. 7의 (a1)에서 단일 체임버를 갖는 인공부레의 경우, 시간 echo 응답은 두 개의 분리된 산란신호가 일정한 간격을 유지하면서 head-up 30°의 자세각부터 head-down 약 30° 부근까지 연속적으로 출현하였다. 또한, 가장 강한 echo 응답은 자세각 0°가 아닌 −5.5° 부근에서 출현하고 있는데, 이것은 인공 부레가 완전히 좌우 대칭적인 방추형 구조가 아닌 점과 인공 부레를 수조에 현수시킬 때, 설치상의 오차가 발생했기 때문인 것으로 판단된다. 또한, (b1)의 이중 체임버를 갖는 인공부레의 경우, 시간 echo 응답은 head-up 35°부터 head-down −35°까지 연속적으로 출현하고 있지만, 그 응답특성은 단일 체임버의 경우와 완전히 다른 변동패턴을 나타내었다. 즉, 가장 강한 echo 응답은 자세각 −2.5° 부근에서 출현하였고, 이 자세각을 기준으로 echo peak 진폭은 각각 head-up과 head-down 방향에서 빗살무뉘 모양으로 휘어져 전개되는 변동특성을 나타내었다.
한편, Fig. 7(a2)의 단일 체임버를 갖는 인공부레에 대한 압축펄스파형의 echogram을 살펴 볼 때, ±35°의 자세각 범위에 걸쳐 한 개의 산란신호만이 매우 선명하게 출현하였고, 자세각 0° 부근에서는 인공부레로부터의 echo 펄스신호의 왕복 시간이 가장 크게 되어 echo 응답패턴이 활 모양으로 휘어진 형상을 나타내었다. 그러나, (b2)의 이중 체임버를 갖는 인공부레의 경우에는 ±35°의 자세각 범위에 걸쳐 2개의 펄스응답패턴이 매우 선명하게 분리되어 나타났는데, 이들 2개의 응답패턴형상은 단일 체임버의 경우와 거의 유사하였다. Fig. 7에서 (b2)에서 두 압축펄스 응답파형은 자세각 0°에서 서로 교차한 후, ±35°의 head-down, 또는 head-up 방향으로 이동해 감에 따라 응답파형의 간격이 점차 넓어져 가는 변동특성을 나타내었다. 이 현상은 Fig. 1(b)에서 앞쪽 체임버에 대한 길이 및 체적이 뒤쪽 체임버의 그것보다 크기 때문에 head-up 상황에서는 chirp 펄스가 큰 체임버에 먼저 부딪쳐 산란신호가 생성되고, 그 다음으로 작은 체임버로부터의 산란신호가 순차적으로 생성되기 때문이다. 반면, 부레의 자세가 head-down 상황으로 전환되면 head-up 상황과는 정반대로 뒤쪽의 작은 체임버에 의한 산란신호가 먼저 생성되고, 앞쪽의 큰 체임버에 의한 산란신호는 나중에 생성되는 echo 생성 매커니즘을 나타낸다. 이 때문에 Fig. 7의 (b2)에서 앞쪽의 큰 체임버에 대한 압축파형의 진폭이 뒤쪽의 작은 체임버에 대한 진폭보다 강하게 출현하였다.
한편, Fig. 7에서 펄스압축기법에 의한 펄스응답패턴 (a2) 및 (b2)를 일반 어군탐지기의 echo 응답패턴 (a1) 및 (b1)와 비교해 볼 때, SNR과 echo 응답신호의 공간 분해능이 대폭 개선된 것을 알 수 있다. Fig. 7(b2)의 이중 체임버를 갖는 모형부레에 대한 펄스압축파형의 변동패턴을 토대로 본 연구에서 실험적으로 구축한 chirp 어군탐지기의 공간식별 분해능을 분석, 고찰한 결과는 Fig. 8과 같다. Fig. 8에서 종축은 Fig. 7의 (b2)에 나타낸 펄스압축파형의 echogram에서 각 자세각에 대한 두 압축파형의 시간차(
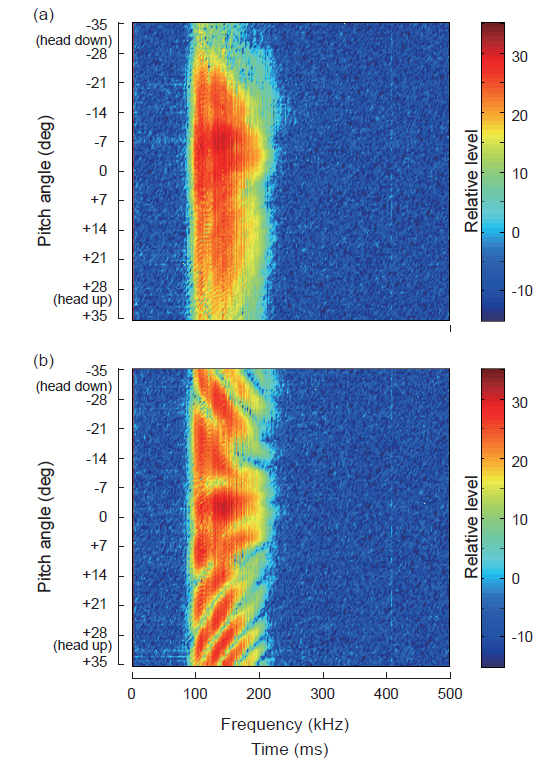
Fig. 9에서 (a)는 각각 단일 체임버를 갖는 인공부레에 대한 산란신호의 주파수 스펙트럼이고, (b)는 이중 체임버를 갖는 인공부레에 대한 산란신호의 주파수 스펙트럼이다.
Fig. 9에서 종축은 자세각(°)이고, 횡축은 주파수(kHz)이다. 또한, 산란신호의 주파수 스펙트럼 레벨은 −15~35dB 범위의 상대진폭으로 나타내었다. Fig. 9(a)의 단일 체임버에 대한 주파수 스펙트럼의 변동특성은 Fig. 7(a1)의 시간응답특성과 유사하게 0°~15°의 자세각 범위에서는 95-220 kHz의 전 주파수 범위에서 강한 응답특성을 나타내었지만, 이 자세각 범위를 벗어나면 응답특성이 점진적으로 약화되는 경향을 나타내었다. 한편, Fig. 9(b)의 이중 체임버를 갖는 인공부레의 경우에는 Fig. 7(b1)에 나타낸 시간 echo 응답특성과 매우 유사하게 head-up 35°부터 head-down −35°까지 연속적으로 대칭적인 응답패턴을 나타내었는데, 그 응답특성은 단일 체임버의 경우와 완전히 다른 변동패턴을 나타내었다. 즉, 가장 강한 echo 응답은 자세각 −2.5° 부근에서 출현하였고, 이 자세각을 기준으로 스펙트럼 peak 진폭은 각각 head-up과 head-down 방향으로 빗살무뉘 모양으로 휘어져 전개되는 변동특성을 나타내었다.
Fig. 1의 단일 체임버와 이중 체임버를 갖는 인공부레에 대한 반사강도의 자세각 및 주파수 의존성을 나타낸 결과는 Fig. 10과 같다. Fig. 10에서 종축은 자세각(°)이고, 횡축은 주파수(kHz)이다. 또한, 이들 인공 부레에 대한 반사강도 레벨은 −70~−30 dB 범위의 컬러 스케일로서 나타내었다. Fig. 10(b)에서 실선은 인공부레를 등가적인 길이
Fig. 10(b)의 이중 체임버를 갖는 인공부레(길이 67 mm, 이중 체임버의 중심간격 35 mm)에 대한 반사강도 패턴에서 null 위치는 SINC 모델에 의한 계산된 패턴과 매우 유사한 변동특성을 나타내었고, 또한, SINC 모델에 의한 인공부레의 등가적인 길이의 추정치는 35 mm이었다. Fig. 1(b)에서 이중 체임버를 갖는 인공 부레의 경우, 두 체임버 사이의 중심간격은 35 mm로서, 이 값은 SNC 모델에 의해 추정한 인공부레의 등가적인 길이 35 mm와 완전히 일치한다. 이와 같은 결과로부터 Fig. 10(b)에 나타낸 반사강도의 자세각 및 주파수 의존성 패턴은 Fig. 1(b)의 두 체임버로부터 산란된 산란신호가 서로 보강 및 상쇄간섭을 일으켜 생성된 것임을 알 수 있었다.