지수식 및 순환여과식 뱀장어양식장에서는 어류의 빠른 성장과 생산량 증대를 위해 고수온과 고밀도 사육조건에서 과량의 사료가 공급되어 사육수중에는 질소성 대사산물을 포함한 자가오염원의 부하량이 비교적 높다. 담수어류의 아질산 중독은 수중의 암모니아를 아질산(NO2-)과 질산(NO3-)으로 산화하는 생물여과시스템의 용량 부족, 효율저하 및 기능부전 등에 따른 수계의 질화작용 불균형에 의해 나타나며(Kroupova et al., 2005), 질화작용의 불균형 발생 시에 사육수의 아질산 농도는 1 mM~3.57 mM에 이르기도 한다(Kamstra et al., 1996; Kroupova et al., 2005). 질화세균에 의한 질화효율은 수온, pH, 용존산소, 질화세균의 수 및 수중 유해물질(항생물질 등)의 존재 여부에 영향을 받는다(Russo and Thurston, 1991; Jensen, 2003).
양식장 사육수중에 유기물이 축적되면 양식어류에게 만성적인 스트레스를 유발할 수 있는 암모니아, 아질산 및 황화수소와 같은 미생물의 대사산물이 많이 생성된다(Das et al., 2004). 이러한 환경조건에서 초래되는 어체 스트레스는 궁극적으로 어류의 성장저하, 면역억제, 질병발생 및 폐사를 유발한다(Snieszko, 1974; Lewis and Morris 1986). 담수어류에 대한 아질산 독성과 생리적 영향에 관해서는 많이 연구되었으며, 아질산 독성은 어종에 따라 많은 차이가 있다(Lewis and Morris 1986; Haung and Chen, 2002; Jensen, 2003; Tomasso and Grosell, 2005).
어류 아가미를 통해 혈액에 유입된 아질산은 적혈구속에 확산되어 hemoglobin (Hb)을 산소결합 능력이 없는 methemoglobin (metHb)으로 산화시켜서 혈액의 산소운반능력이 감소된다(Jensen and Hansen, 2011). 어류 적혈구는 metHb를 기능적 Hb로 재전환하는 NADH-methemoglobin reductase (NMR)를 함유하고 있으며(Huey and Beitinger, 1982; Avilez et al., 2004), 메트헤모글로빈혈증의 어류를 nitrite-free water에 유지하면 Hb 비율은 정상적으로 회복된다(Huey et al., 1980; Knudsen and Jensen, 1997). 아질산에 노출된 어류의 Hb와 metHb농도는 아질산에 의해 생성된 metHb 및 NMR이 metHb을 Hb으로 환원시킨 생리작용의 결과이다(Lewis and Morris, 1986). 따라서 아질산에 노출되어진 뱀장어의 NMR 활성은 수중의 아질산 농도에 따라 차이가 있을 것으로 추정된다.
어류의 혈액학적·면역학적 parameter는 내생적 또는 외생적 환경변화에 따른 생리적 스트레스 반응의 지표로 이용되며 (Santos and Pacheco, 1996; Alexander et al., 2012), 특히 혈중 cortisol과 glucose 농도는 스트레스 반응의 일반적인 지표로 많이 활용된다(Iwama et al., 1997; Tsuzuki et al., 2001). 수계 오염원에 노출된 수서생물은 체조직의 산화적 손상을 유발하는 H2O2, O2− 및 OH−와 같은 활성산소가 증가한다(Livingstone et al., 1990; Livingstone, 2001; Martinez-Alvarez et al., 2005). Superoxide dismutase (SOD)는 O2−를 H2O2로 전환하는 효소로서 세포 항산화제이며, 세포 내외에 존재하는 SOD 함량은 활성산소의 산화적 스트레스에 의해 발생되는 질병 예방에 중요하다(Malstrom et al., 1975). Catalase (CAT)는 호기성 세포에 존재하는 항산화제로서 H2O2 해독에 관여하고, SOD와 CAT의 활성도는 수서생물의 환경스트레스에 대한 biomarker로 이용되고 있다(Martinez-Alvarez et al., 2005; Alexander et al., 2012).
아질산에 노출된 어류는 전해질 불균형이 나타나고(Jensen, 2003), 아질산 축적과정에는 골격근과 적혈구의 K+ 상실에 의해 K+ 항상성이 교란되어 고칼륨혈증이 야기될 수 있다(Jensen et al., 1987; Jensen, 2003). 또한, 메기류(
한편, Amano et al. (1981)은 아질산 2.14 mM에 1개월간 사육된 뱀장어는 모든 개체에서 15-68%의 메트헤모글로빈혈증을 관찰하였다. 그러나 아질산 노출어류에서 혈중 아질산이 최대에 도달하는 기간은 24-48시간이며(Eddy et al., 1983; Aggergaard and Jensen, 2001; Huang and Chen, 2002), 반수치사농도(LC50)는 24시간 이후에도 감소하지만 그 감소율은 낮기 때문에 다른 독성오염원과 같이 아질산에 대한 급성영향시험의 적절한 노출시간은 24-96시간이라고 하였다(Lewis and Morris, 1986).
본 연구는 국내 내수면 양식장의 양식주종인 뱀장어(
실험에 사용된 뱀장어는 양만장에서 사육중인 것(생체중량, 181-207 g)을 실험실로 운반한 다음 원형수조(500 L)에 수용하여 사육하였다. 사육 동안 수온은 28±0.5℃로 유지하였고, 먹이는 상품 배합사료를 물로 반죽하여 2일마다 체중의 약 5%를 공급하며 실험조건에 2주간 순치·사육하였다. 모든 실험어는 실험개시 전에 2일간 절식시켰다.
시험용수는 수돗물을 저수탱크에 채우고 하룻동안 aeration과 함께 수중펌프로 자외선살균기(P392, Samjee-tech, Korea)를 통과시킨 것을 실험에 사용하였다. 시험용액은 전자저울(Sartorius BP 221S)로 NaNO2 무게를 달아 시험용수에 희석하여 아질산-질소 용액을 제조하였다. 본 연구에서 적용한 아질산 농도는 Amano et al. (1981)이 뱀장어를 아질산에 1개월간 노출시킨 후 모든 개체에서 메트헤모글로빈혈증(15-68%)이 관찰된 2.14 mM을 최소농도로 삼았으며, 실험시작 시에 아질산 농도별 실험수조(실험구)에서 실측한 아질산 농도는 0 (대조구), 2.33, 4.60, 6.64 및 8.78 mM이였다(Table 1).

Levels of actual ambient nitrite and water quality of experimental chambers. Values are mean±S.E. (n=6). Hardness and electrolytes were measured for dilution water (fresh water) of nitrite
각 실험구는 뚜껑과 잠금장치가 달린 반투명 플라스틱 사각수조(20 L, 38×25×22 cm)에 공기공급 장치를 설치하였고, 대조구를 포함한 농도별 실험구는 6반복으로 구성되었다. 노출실험은 시험용액 10 L을 실험구에 채운 다음 실험어를 각각 1마리씩 수용하여 48시간 동안 노출되도록 하였고, 실험어의 탈출을 방지하고 시험용액의 증발을 줄이기 위해서 실험구 뚜껑을 닫고 잠금장치를 하였다.
실험구의 아질산 농도는 APHA (1995)의 NED법으로, 수온, pH 및 DO는 수질분석기(YSI 6920, USA)로 측정하였다. 시험용수의 전해질은 ICP/MS (PerkinElmer Nexion 300X)로서, 경도는 Ca2+과 Mg2+ 농도를 ICP/MS로서 측정하여 CaCO3 등 가량으로 환산하는 간접법(APHA, 1995)에 의거하였다.
아질산에 48시간 노출시킨 후 실험어 마취는 Chiba et al. (2006)이 뱀장어에 적용한 전기마취를 활용하였다. 마취는 실험구 양쪽 끝의 시험용액 속에 240V/AC 승압 전류가 흐르는 전극을 각각 넣은 뒤 30초 동안 전기충격 마취 후 heparinized syringe (23G)로 미병부 혈관에서 혈액을 약 1.5 mL 채취하였다. 채혈액의 1.0 mL은 4℃에서 5,000 rpm으로 5분간 원심분리한 다음 혈장과 침강된 적혈구를 각각 수거하여 −80℃의 deep freezer (DF-8510, Ilshin, Korea)에 두고 혈장 생화학적 분석 및 NMR 활성 분석시료로 사용하였고, 나머지 혈액은 PCR tube에 주입한 후 냉장보관하며 혈액분석에 사용하였다. 채혈이 끝난 실험어는 전자저울(Sartorius BP 3100S)로 생체중량을 측정한 후 어체를 해부하여 아가미와 간을 적출하였다. 적출 조직은 해부과정에 묻은 적혈구나 혈병을 제거하기 위해 즉시 냉장 PBS 용액(pH 7.4)으로 헹군 후 chilled conical tube (15 mL)에 담아 −70℃에 보관하였다. CAT의 분석에는 조직 g당 50 mM phosphate buffer (1 mM EDTA, pH 7.0) 냉장액 5 mL를, SOD 분석에는 조직 g당 20 mM HEPES buffer (1 mM EDTA, 210 mM mannitol 및 70 mM sucrose, pH 7.2) 냉장액 5 mL를 주입한 후 homogenizer (Sonopuls HD 2200, Germany)로 균질화하였다. 균질액은 4℃에서 1,500
Drabkin's reagent 5 mL에 전혈 20 μL를 주입하여 혼합한 후 실온에 20분간 방치한 다음 분광광도계로 540 nm에서 비색 정량하는 상법으로 분석하였다.
혈액 5 μL에 증류수 1 mL을 넣어서 적혈구를 용혈시킨 다음 0.1 M phosphate buffer (pH 6.8)를 0.5 mL 주입하였다. 용혈액은 450
혈액 1.0 mL에서 수거된 적혈구를 냉장 생리식염수 10 mL에 부유시켜 1,000
Griess reagent (Promega G2930) 및 그 manual에 따라 540 nm에서 흡광도를 측정하여 검량선으로부터 혈장 아질산 농도를 산출하였다.
혈장 GOT, GPT, glucose 및 전해질(Na+, K+, Cl−)은 혈액생화학분석기(FUGI- Drichem 3500s)로 측정하였다.
혈장 cortisol은 cortisol ELISA kit (IBL RE52061, Germany)로 분석하였다.
혈장, 간 및 아가미 조직의 CAT 활성은 CAT assay kit (Cayman chemical, catalog No. 707002)로 분석하였다. 1 unit CAT는 25℃에서 1분당 단백질 1 mg이 반응하여 1.0 nmol의 formaldehyde를 생성하는 효소량을 나타낸다.
SOD 활성은 SOD assay kit (Cayman chemical, No. 706002)로서 측정하였다. 1 unit SOD는 25℃에서 단백질 mg당 O2−의 50% dismutation에 요구되는 효소량을 나타낸다.
혈장과 적출 조직의 단백질 함량은 Bradford protein assay kit (Bio-Rad, No. 500-0202)로 측정하였다.
각 분석 결과치는 one-way ANOVA를 실시하여 평균간의 차이를 검정하였다. 이로부터 95% 유의수준에서 그 차이가 인정된 측정치는 SPSS (version 19.0) 통계패키지를 이용하여 Duncun’s multiple range test를 실시하였다(
수온 28.3℃에서 아질산(0, 2.33, 4.60, 6.64 및 8.78 mM)에 뱀장어를 48시간 노출시킨 후 혈장 아질산 농도를 측정하였다(Fig. 1A). 혈장 아질산은 대조구에서 평균 15.5±1.38 μM이었으며, 아질산 2.336.64 mM에 노출된 실험어의 혈장 아질산은 23.1-62.3 μM로서 수중의 아질산 농도가 높을수록 혈장에 축적된 아질산이 많았다. 아질산 8.78 mM에 48시간 노출 후 생존 개체의 혈장 아질산 농도는 370.7 μM이였다.
아질산 4.60 mM에 노출된 실험어의 Hb 함량은 대조구와 비해 낮은 값을 보여주었고, 수중의 아질산 농도가 높을수록 Hb는 감소하는 경향을 나타내었다(Fig. 1B). Hb중에서 metHb 비율은 대조구에서 평균 3.7%로 측정되었으며, 수중의 아질산 농도가 높을수록 현저하게 증가하였다(
아질산에 노출된 뱀장어의 NMR 활성을 측정한 결과는 Fig. 2와 같다. 대조구의 NMR 활성은 0.019±0.0014 U/mg Hb이었고, 아질산 2.33~6.64 mM에 노출된 뱀장어는 평균 0.084~0.188 U/mg Hb로서 아질산 농도가 높을수록 증가하였다(
수온 28.3℃에서 농도별 아질산에 48시간 노출 후 혈장 N+, K+ 및 Cl−를 분석하였다(Table 2). 혈장 N+와 Cl−는 환경수중의 아질산 농도에 따른 차이는 없었다. 아질산 6.64 mM에 노출된 개체의 혈장 K+ 농도는 대조구에 비하여 유의하게 높은 값이 관찰되었다(

Levels of plasma electrolyte, cortisol and glucose of Anguilla japonica exposed to elevated ambient nitrite for 48 h at 28.3℃. Data expressed as mean±S.E. (n=6). Different letters indicate significant differences (P<0.05) between nitrite-N concentrations. Asterisks represent that the measurement of one survived fish following 48 h exposure
아질산에 48시간 노출시킨 뱀장어의 혈장 cortisol과 glucose를 분석한 결과는 Table 2와 같다. Cortisol은 환경수중의 아질산 농도가 증가할수록 유의하게 높은 값을 보여주었다(
아질산 ≤4.60 mM에 노출된 뱀장어에서는 혈장 GOT와 GPT의 활성 차이는 없었으며(Fig. 3), 6.64 mM에 노출된 개체는 대조구에 비하여 높은 활성을 나타내었다(
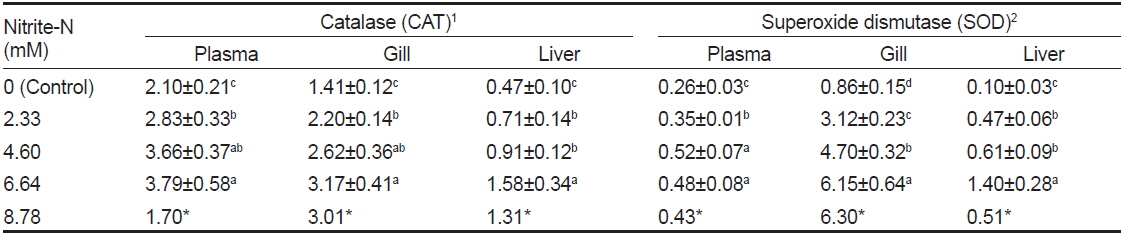
수온 28.3℃에서 농도별 아질산에 48시간 노출 후 혈장, 아가미 및 간의 CAT와 SOD 활성을 측정한 결과는 Table 3과 같다. 아질산 2.33 mM에 노출된 뱀장어의 CAT와 SOD 활성은 대조구에 비하여 유의하게 높았으며, 환경수중의 아질산 농도가 높을수록 증가하였다(

Antioxidative enzyme activity of Anguilla japonica exposed to elevated ambient nitrite for 48 h at 28.3℃. Data expressed as mean±S.E. (n=6). Different letters indicate significant differences (P<0.05) between nitrite-N concentrations. Asterisks represent that the measurement of one survived fish following 48 h exposure
어류는 환경수중에 아질산이 존재하면 염류세포의 Cl− 흡수부위의 일부가 아질산 흡수부위로 이용되어 혈중에 아질산이 유입된다(Jensen, 2003). 따라서 아질산은 염류세포의 Cl− 흡수과정에서 경쟁자로 작용하고, 이 때문에 수중에 Cl−가 일정한 농도로 존재하는 조건에서는 어류의 아질산 흡수 및 아질산 독성이 완화된다(Williams and Eddy, 1986; Madison and Wang, 2006).
경골어류의 혈중 아질산 축적은 담수어에 대하여 많이 연구되었다. 담수산 송어류의 일종인
어류 혈액으로 유입된 아질산은 적혈구의 막단백질을 경유하여 적혈구속에 유입되어 Hb을 metHb으로 산화시킨다(Jensen, 2003). MetHb은 산소결합 능력이 없기 때문에 궁극적으로 혈액의 산소운반 능력이 감소되며, 이로 인하여 초래되는 산소 부족은 어체에 스트레스 요인으로 작용할 수 있다(Kroupova et al., 2005). 어류가 저산소에 대응하는 일반적인 반응은 조혈작용에 의한 적혈구수 증가이며, 이것은 어체내 요구 산소를 일정 수준으로 유지하기 위하여 나타나는 현상이지만(Peterson, 1990), 여러 어종에서는 상반되는 결과도 보고되어 있다(Hilmy et al, 1987; Tucker et al., 1989; Avilez et al., 2004). 본 연구에서 아질산에 노출된 뱀장어의 Hb 함량은 환경수중의 아질산 농도가 높을수록 감소하였다(Fig. 1B). 환경수중의 아질산 농도가 ≥4.60 mM인 조건에서 Hb 함량이 감소하는 현상은 순환혈액중의 적혈구 사멸 증가 및 적혈구의 용해에 기인(Jensen, 1990a; Knudsen and Jensen, 1997)되거나 체내 저산소에 직면한 어류의 조혈기능 저하에 의한 것(Gill et al., 1991)으로 생각된다.
아질산에 노출된 어류의 특징적인 반응은 Hb이 산화되어 metHb이 증가하는 현상이며, nitrite-free water에서도 어류는 Hb의 자가산화에 의해 측정이 가능한 량의 metHb을 함유한다(Lewis and Morris, 1986; Kroupova et al., 2005). 뱀장어는 Hb의 3.7%를 metHb 바탕농도로 함유하였고, 아질산 2.33-6.64 mM에서 48시간 후에는 9.5-56.0%, 8.78 mM에 노출 후 생존 개체는 90%로 나타났다(Fig. 1C). 수온 25.2℃에서 유럽산뱀장어의 metHb 바탕농도는 8.4%, 아질산 0.356-1.549 mM에 24시간 노출 후에는 16.6-45.6% (Haung and Chen, 2002), 그리고 Amano et al. (1981)은 아질산 2.14 mM과 2.86 mM에 1개월간 노출된 뱀장어의 metHb 비율은 각각 15-68% (평균, 38%)와 15-65%(평균, 40%)로 보고하였다. Kiese (1974)는 Hb의 자가산화율은 pH 7일 때 보다 pH 5.7에서 훨씬 높으며, Hb의 50%가 탈산소 되는 낮은 산소분압에서 최대에 이른다고 하였다. Huey et al. (1984)은 양식장 사육수의 pH는 양식 어류의 호흡과 수중 호기성 미생물이 배출하는 CO2에 의해 빠르게 산성화 되지만 질화세균,
뱀장어 적혈구의 NMR 활성을 측정한 바, 대조구에서는 Hb mg당 0.019 U, 아질산 2.33-8.78 mM에 노출된 것은 0.084-0.272 U로서 환경수중의 아질산 농도가 높을수록 NMR 활성이 유의하게 증가하는 현상을 보여주었다(Fig. 2). 어류 적혈구는 metHb을 Hb으로 환원하는 효소인 NMR을 함유하며(Huey et al., 1984), 아질산에 노출된 어류를 nitrite-free water에 두면 NMR의 환원기작에 의해 Hb 함량은 정상수준으로 회복된다(Huey et al., 1980; Knudsen and Jensen, 1997; Lefevre et al., 2011). 열대산 송어류의
환경수중의 아질산과 Cl−는 아가미에서 동일 흡수경로를 경쟁함에 따라 아질산에 노출 시에는 능동적 Cl− 흡수 부분의 일부는 아질산흡수 부위로 전환된다(Jensen, 2003). 반면에 저삼투성 환경수중으로는 Cl−의 수동적 유출이 계속되어 Cl
한편, cortisol과 glucose는 아질산 농도가 높을수록 증가하였다(Table 2). 아질산에 노출된 뱀장어에서 혈장 cortisol의 증가하는 것은 cortisol이 간에서 glucose 신합성을 촉진하여 NMR 활성을 높이도록 유도하는 일련의 생리적 조절반응(Kiese, 1974) 내지는 메트헤모글로빈혈증에 의한 어류의 요구산소 부족에 따른 스트레스 반응으로 생각된다.
혈중으로 유입된 아질산은 어류의 질소대사를 교란시킬 뿐만 아니라 간의 손상을 초래하여 GOT와 GPT 활성이 증가한다(Michael et al., 1987; Haung and Chen, 2002). 이들 아미노기 전이효소의 상당 부분은 손상된 간세포에서 분비되어 혈액으로 방출됨으로 혈장 GOT와 GPT 활성 측정치는 간 손상의 지표로 이용되고 있다(Michael et al., 1987). 아질산 6.64 mM에 노출된 뱀장어는 대조구에 비하여 GOT와 GPT 활성이 증가하였다(Fig. 3). Haung and Chen (2002)은 아질산 0.751 mM에 24시간 노출된 유럽산뱀장어는 GOT 활성이, 1.547 mM에서는 GPT 활성이 유의하게 증가하는 결과를 보여 주었다.
세포 내외에 존재하는 CAT와 SOD 함량은 활성산소의 산화적 스트레스에 의해 발생되는 질병 예방에 중요하며(Malstrom et al., 1975), 항산화효소의 활성도는 수서생물에 있어서 환경 스트레스에 대한 biomarker로 이용될 수 있다고 하였다(Martinez-Alvarez et al., 2005; Alexander et al., 2012). 아질산 2.33 mM에 48시간 노출된 뱀장어는 CAT와 SOD 활성이 유의하게 높았고, 환경수중의 아질산 농도가 높을수록 이들의 활성은 증가하였다(Table 3). CAT는 간에 비하여 혈장과 아가미에서, SOD는 아가미에서 높은 활성을 나타내었다.
본 연구의 결과, 수온 28.3℃에서 농도별 아질산에 48시간 노출된 뱀장어는 환경수의 아질산 농도가 높을수록 혈장 아질산, metHb 비율, NMR, CAT 및 SOD의 활성은 증가하였고, Hb 함량은 감소하였다. 환경수의 아질산 농도가 ≥2.33 mM인 조건에 뱀장어가 급성으로 노출되면 생리적 스트레스 반응과 더불어 혈액학적 parameter와 항산화효소의 활성이 현저하게 변화되는 현상이 관찰되었다.