참굴(
지금까지 우리나라에서는 양성용 참굴 인공종묘는 2배체(diploid)에 국한하여 생산되어져 왔으나, 최근 염색체 조작에 의한 3배체 굴 양식은 미국, 프랑스, 호주, 중국 등 많은 나라에서 각광받고 있다. 그 이유는 3배체(triploid) 참굴은 2배체에 비해 질병내성이 강하며, 생물학적으로 불임이기 때문에 성숙에 필요한 에너지를 성장과 육질부 비만에 이용할 수 있으며, 또한 주 산란철인 여름철에도 수확이 가능하기 때문이다(Guo et al., 1996; Park et al., 1999). 따라서 3배체 참굴 양식은 현재와 같이 열악해져 가는 양식환경에서 보다 높은 생산성과 수익성을 기대해 볼 수 있다. 3배체 굴은 1980년대 Stanley et al. (1981)에 의해 버지니아굴(
2배체 참굴의 정자동결보존은 국내외적으로 그 기술이 확립되어져 왔으나(McFadzen, 1995; Adams et al., 2004; park et al., 2013), 4배체의 경우 Dong et al. (2005a, 2006) 연구를 제외하고는 거의 찾아볼 수 없는 실정이며, 또한 이들 연구의 결과, 4배체 정자를 동결보존하여 해동하였을 때 운동성, 수정률 및 부화율이 2배체 정자보다 현저히 낮은 것으로 나타나, 아직 4배체 정자의 동결보존에 대한 연구가 더 이루어져야 할 것으로 여겨진다. 따라서 본 연구에서는 4배체 참굴 정자동결보존의 확립에 기여하고자 적정 동해방지제, 평형시간, 냉각속도 등을 탐색하였다.
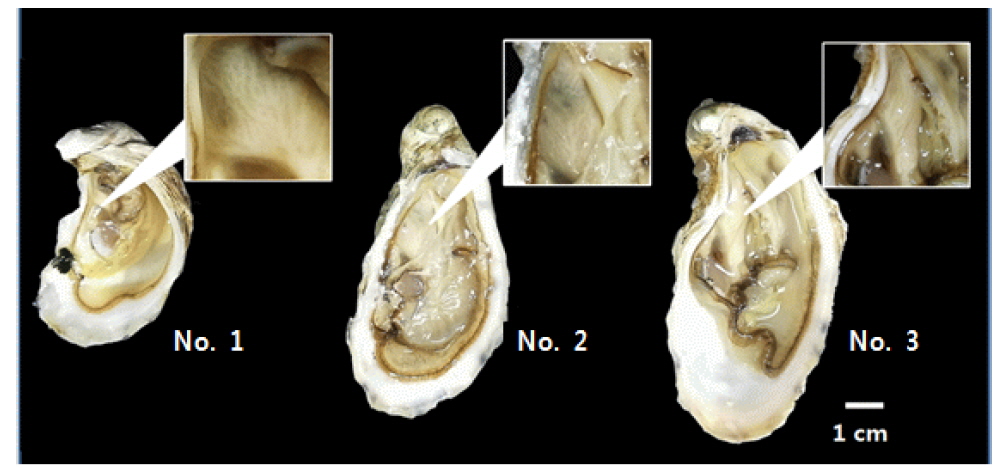
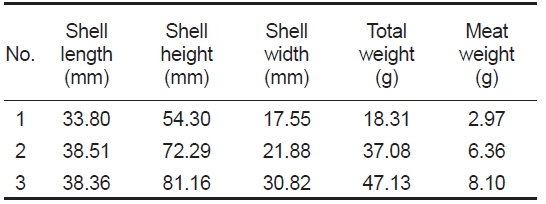
4배체 참굴 모패는 2014년 3월 20일에 미국 4Cs Breeding Technologies, Inc.로부터 구입하여 국립수산과학원 남동해수산연구소 남해센터의 1톤 FRP에 유수식으로 수용하였다. 구입당시 모패는 미성숙상태였으며, 모패의 성숙을 위해
[Table 1.] Basic parameters of tetraploid Pacific oyster Crassostrea gigas used for this experiments
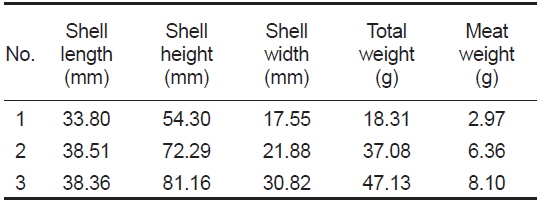
Basic parameters of tetraploid Pacific oyster Crassostrea gigas used for this experiments
수송 직후 각 개체의 생체로부터 정소를 적출한 후 Calcium-free Hanks’balanced salt solution (C-F HBSS: 450 mM NaCl, 18 mM KCl, 3 mM MgSO4·7H2O, 1 mM Na2HPO4·7H2O, 1 mM KH2PO4, 14 mM NaHCO3, 18 mM glucose)와 1:10 (w/v)으로 혼합하여 정자 현탁액을 만들어 4℃에 보관하면서 실험에 사용하였다.
4배체 정자동결보존에 사용된 동해방지제는 dimethyl sulfoxide (DMSO), methanol (MeOH), polyethylene glycol (PEG, weights of 200), propylene glycol (PG) 총 4가지를 사용하였으며, 동해방지제 농도는 동결보존할 총량(동해방지제+정자현탁액량)의 10%와 20%로 하였다. 본 실험에 사용된 모든 시약은 Sigma-Aldrich Co.의 제품을 사용하였다.
동결보존시 평형시간 조건을 구명하기 위하여 정자 현탁액을 동해방지제 종류별, 농도별로 혼합한 다음 평형시간을 3, 10, 30분으로 설정하였다. 이때 정자 냉각속도는 −80℃까지 분당 3℃로 냉각시킨 후, 액체질소(−196℃)로 옮겼다.
평형시간 노출 이후 냉각은 프로그램 동결기(Computer Controlled Rate Freezer 14S, SY-LAB, Austria)를 이용하여 4가지의 속도(냉각률)로 하였다:실온→ −80℃ (3, 5, 7, 10℃/분)→ 액체질소). 동해방지제 및 평형시간은 각각 10% DMSO 및 30분으로 하였으며, 0.25 mL straw를 사용하였다.
동결보존시 적정 straw의 크기를 조사하기 위하여 0.25 및 0.5 mL의 2가지 straw (Cassou, IMV, France)를 사용하였다. 이때 동해방지제 및 평형시간은 각각 10% DMSO 및 60분으로 하였으며, 동결은 −80℃까지 분당 3℃로 냉각시킨 후, 액체질소 (−196℃)로 옮겼다.
정자 동결후 액체질소에서 24시간 보관하였으며, 이후 straw를 30℃항온수조에서 7초간 급속 해동한 다음 정자의 생존율 및 운동성을 측정하였다.
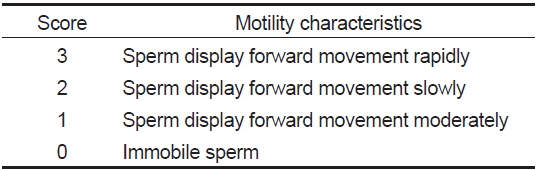
정자의 생존 및 운동성은 동결 전·후의 정자현탁액 2 μL를 C-F HBSS 40 μL와 섞어 정자 운동성 관찰용 슬라이드글라스 (Teflon Printed Glass Slide; 21 wells; diameter of each well, 4 mm; Funakoshi Co., Japan)에 2 μL를 분주하여 광학현미경 (Axio Scope A1, Carl Zeiss, Germany)으로 관찰하였다. 운동성을 가지는 모든 정자는 생존한 것으로 간주하였으며, 생존율이 1% 이하는 1%로 나타내었다. 정자 운동성은 운동지수(Table 2)에 따라 점수를 부여 하고, 각각의 운동점수와 운동정자의 비율에 따라 Strüssmann et al. (1994)의 방법을 변형하여 정자활성지수(sperm activity index, SAI)로 나타내었다.
[Table 2.] Score for the evaluation of sperm activity index (SAI)
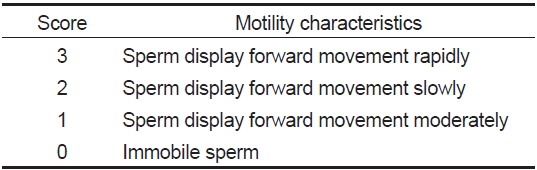
Score for the evaluation of sperm activity index (SAI)
모든 실험은 3회 반복으로 수행하였으며, 결과값은 평균±표준편차로 나타내었다. 유의차는 SPSS-통계패키지(version 18.0)를 이용하여 one or two way ANOVA-test (Duncan's multiple range test) 및
3마리의 4배체 참굴 정자의 동결보존 전 생존율은 88.8±2.1-92.6±1.2%였으며, 정자활성지수(SAI)는 2.7±0.1-2.8±0.1로 개체간의 유의적 차이를 보이지 않았다(Table 3).

The survival and activity index (SAI) of sperm from 3 tetraploid Pacific oyster Crassostrea gigas before the cryopreservation
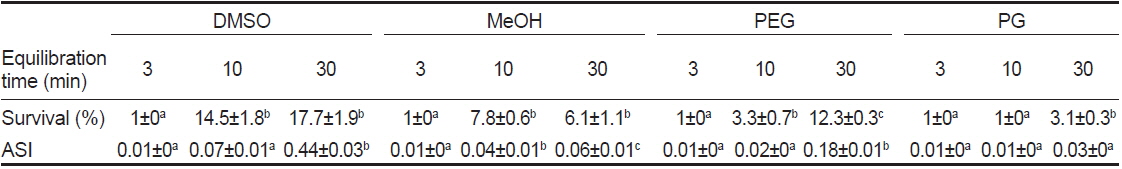
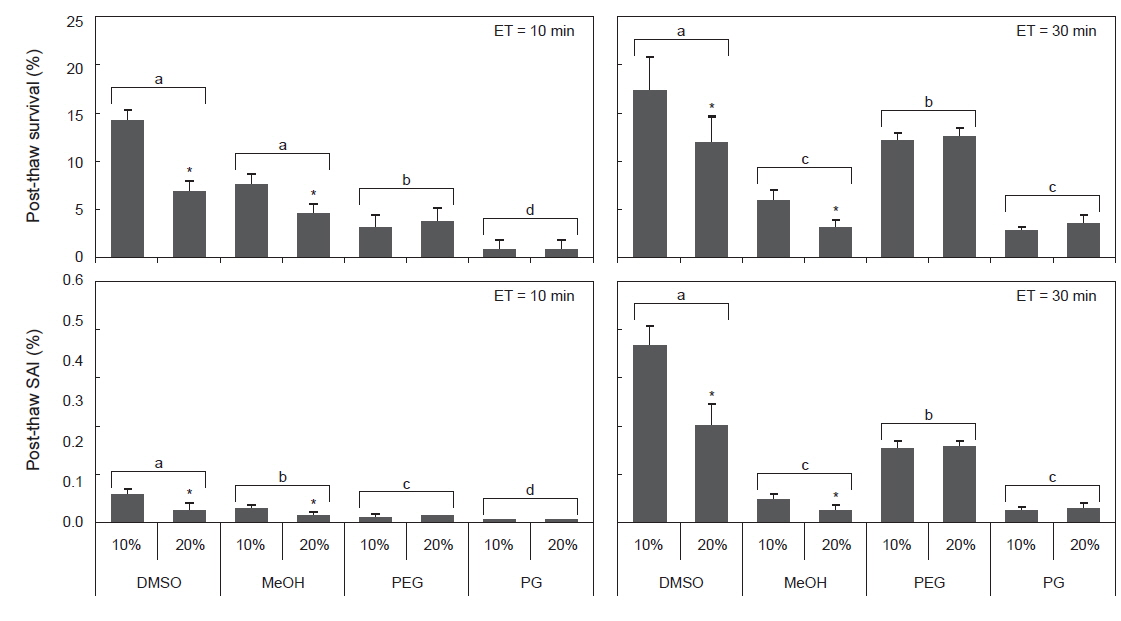
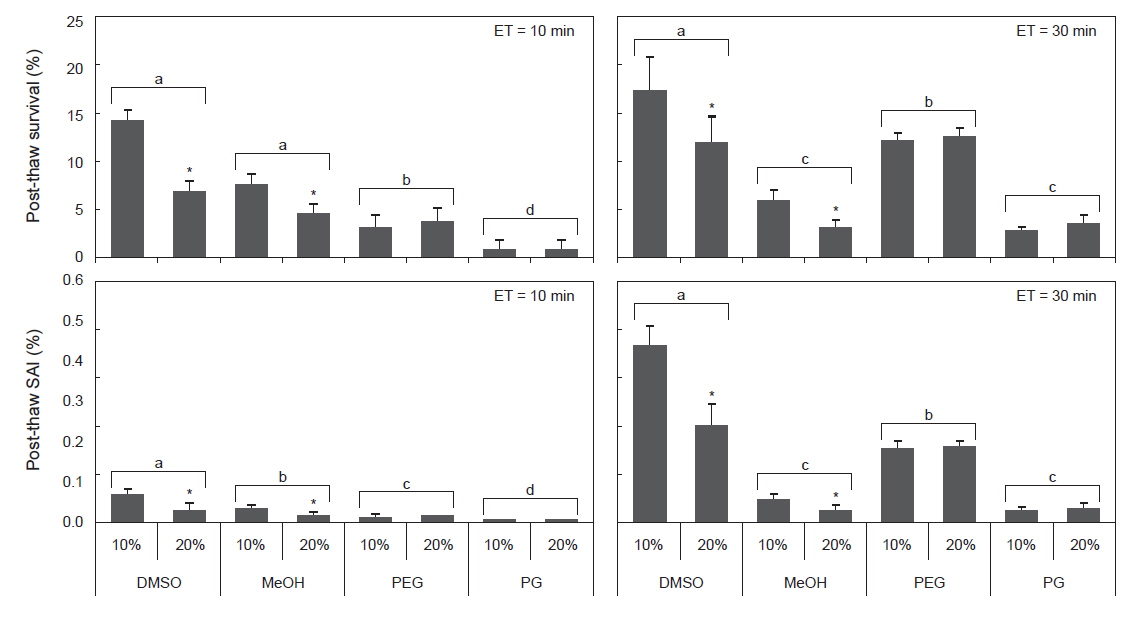
동결보존시 평형시간별 동해방지제에 따른 정자의 해동후 생존율 및 SAI를 조사한 결과, 평형시간이 3분일 때, 모든 동해방지제에서 해동후 생존율이 1%이하, SAI는 0.01로 나타났다(Table 4). 평형시간이 10 및 30분일 때, 해동후 생존율은 DMSO가 다른 동해방지제보다 유의하게 높았으며, 또한 DMSO와 MeOH의 10%가 20%보다 높았다(Fig. 2). 해동후 SAI는 생존율과 동일한 양상으로 나타났다.
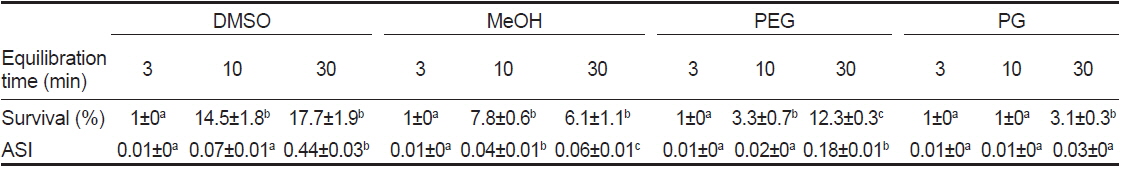
Post-thaw survival and activity index (SAI) of sperm suspended in various cryoprotectant agents (CPAs) of 10% at equilibration time of 3, 10 and 30 min
실험에 사용한 10% 동해방지제를 대상으로 평형시간별 해동 후 생존율 및 SAI의 유의차를 검증한 결과, DMSO 및 MeOH에서는 10 및 30분이 3분보다 유의하게 높았으며, PEG와 PG에서는 30분이 3 및 10분보다 유의하게 높았다(Table 4).
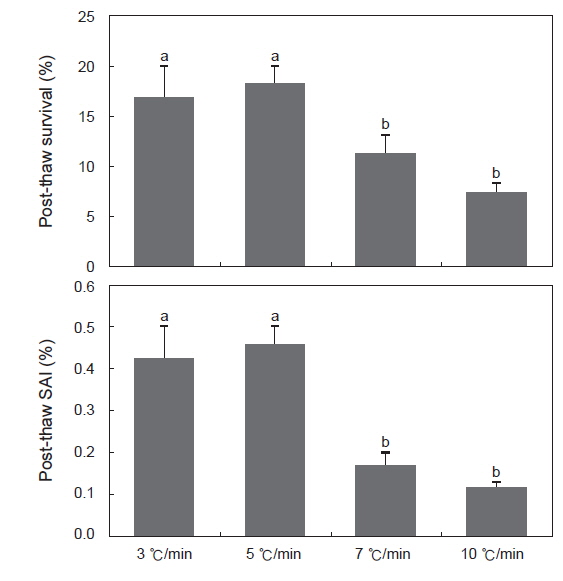
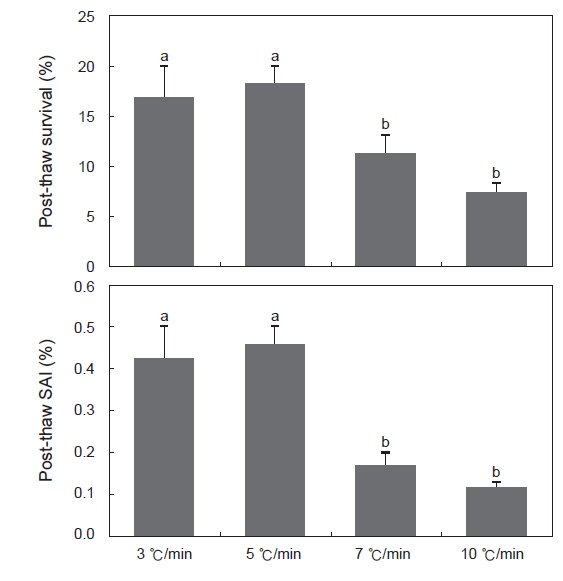
동해방지제로 10% DMSO를, 평형시간은 30분, 0.25 mL straw를 사용하였을 때, 냉각속도에 따른 정자의 해동후 생존율 및 SAI는 Fig. 3에 나타내었다. 냉각속도가 분당 3, 5℃/분일때 생존율은 각각 17.0±3.1%, 18.4±1.8%로 7, 10℃/분의 11.4±1.9%, 7.6±0.7%보다 유의하게 높았다. 또한 SAI 역시 생존율과 유사한 경향을 보였다.
>
Straw 크기에 따른 정자의 해동후 생존율 및 SAI
동해방지제로 10% DMSO를, 평형시간은 30분, 냉각속도는 3℃/분을 사용하였을 때, straw 크기에 따른 정자의 해동후 생존율 및 SAI는 0.25 mL에서 각각 17.0±3.1%, 0.43±0.08로 0.5 mL의 2.1±0.5%, 0.01±0보다 유의하게 높았다(Fig. 4).
4배체 참굴의 정자동결보존은 이미 Dong et al. (2005a, 2006)이 시도한 바 있으나, 해동 후 운동성은 2배체에 비해 매우 떨어지는 것으로 나타나고 있다. 본 연구는 Dong et al. (2006)이 확립한 동결보존 방법을 바탕으로 동해방지별 농도, 평형시간, 동결속도 등의 조건을 보다 확장하여 해동 후 운동성에 어떠한 영향을 미치는지 조사하였다.
동해방지제는 정자의 동결보존 시 가장 중요한 요소로 세포내 삼투질농도와 빙결정형성 등을 완화·조절하는 역할을 한다(Jamieson, 1991). 따라서 동해방지제는 중성물질이어야 하며, 친수성이 강하고, 세포막에 대한 투과성이 높고, 세포에 대한 독성이 적어야 한다(Kuwano and Saga, 2000). 그러나 각 어종의 정자는 동해방지제의 종류에 따라 종 특이성을 보이기 때문에 모든 어류에서 공통적으로 사용할 수 있는 동해방지제는 아직 밝혀진 바 없으며, 동해방지제가 세포 동결시 세포를 보호하는 자세한 메카니즘에 관해서도 아직 알려지지 않고 있다(Kho, 2007). 또한 동해방지제는 세포에 독성을 나타내므로 정자의 동결보존시 적정 동해방지제 종류 및 농도를 찾는 것은 매우 중요하다. 동해방지제는 크게 투과성과 비투과성으로 나눌 수 있는데(Meryman, 1971), 본 연구에서는 투과성인 DMSO, MeOH, PEG, PG를 사용하여 4배체 참굴의 정자를 동결한 결과, 10% DMSO가 가장 적합한 것으로 나타났다. Park et al. (2013)에 따르면 2배체 참굴의 정자동결보존 역시 DMSO가 가장 적합한 동해방지제로 보고하였으며, 그 농도가 15, 10, 20% 순으로 나타났다. 본 연구에서는 정액량의 부족으로 15% 농도를 설정하지는 못하였지만, 아마도 15% DMSO이 적정 농도의 가능성도 배제할 수는 없다. 그러나 2배체 및 4배체 참굴의 정자동결보존시 20%의 DMSO 농도는 정자에 독성을 미치는 것으로 확인 되었다. Dong et al. (2006)은 4배체 참굴 정자동결보존시 동해방지제 2가지를 혼합하였을 때(4% DMSO+6% PEG), 해동후 운동성이 가장 높았으나, 동해방지제 단독 사용시에는 10% DMSO가 가장 양호하다고 보고하여 본 연구와 유사하였다. 일반적으로 DMSO 이외에도 MeOH은 수서 생물의 정자 동결보존에 광범위하게 사용되는 동해방지제로 알려져 있지만(Tiersch, 2000), 굴에서는 MeOH을 동해방지제로 적합하지 않은 것으로도 보고된 바 있다(Smith et al., 2001). 그러나 Dong et al. (2005b)은 2배체 참굴 정자동결보존시 동해방지제로 MeOH이나 MeOH+PEG를 혼합이 가장 적합하다고 발표하였다. Arctic charr
정자의 동결전 평형시간은 동해방지제가 정자에 침투하는데 필요하며, 독성을 최소화해야 한다. 일반적으로 적정 평형시간은 어종, 동해방지제와 동해방지제 종류, 온도에 따라 달라진다(Christensen and Tiersch, 1996; Billard and Zhang, 2001; Sansone et al., 2002). 본 연구에서는 10% DMSO를 동해방지제로 사용하였을 때, 평형시간이 3분인 경우 해동후 생존율 및 SAI가 각각 1% 이하, 0.01 이하로 매우 낮았던 반면 30분일 때에는 생존율 및 SAI가 각각 17.7%, 0.44로 가장 높았으나, Park et al. (2013)은 2배체에서는 평형시간이 3분일 때 10% DMSO에서 생존율이 60%이상, SAI는 2이상으로 보고하여 본 연구의 4배체와는 다른 결과를 보였다. 4배체의 정자동결보존시 적정 평형시간이 2배체보다 긴 이유는 4배체 정자의 세포막이 2배체보다 두꺼워 동해방지제가 침투하는 시간이 더 소요되기 때문인 것으로 추정된다. 실제로 4배체 정자의 첨체, 정자머리, 미토콘드리아, 꼬리는 2배체의 것 보다 유의하게 크므로(Dong et al., 2005c), 이는 막의 두께 또한 클 가능성을 뒷받침해준다.
정자의 동결보존시 적정 냉각속도는 동해방지제의 종류와 농도뿐만 아니라 희석액, 평형시간, 냉각방법에 따라 좌우된다(Babiak et al., 1999). 굴의 정자동결보존을 위한 냉각속도는 1℃/min부터 액체질소로 바로 냉각하는 것까지 광범위하게 보고된바 있다(Hughes, 1973; Hwang and Chen, 1973; Dong et al., 2006). 본 연구에서는 냉각속도를 분당 3, 5, 7, 10℃로 설정하였으며, 5℃에서 가장 해동후 생존율 및 SAI가 높았으나 3℃와는 유의한 차이를 보이지 않았으며, 냉각속도가 빠를수록 생존율은 낮아지는 것으로 나타났다. Dong et al. (2006) 역시 4배체 참굴의 정자동결보존시 적정 냉각속도는 분당 5℃로 보고한 바 있으며, 본 연구와 일치하였다.
수서생물의 정자동결보존시 사용되는 다양한 크기의 straw 또는 cryovial (동결보존용기)은 그 목적에 따라 다르게 사용되는데, 일반적으로 대용량 용기는 주로 현장에서 종묘생산에 사용되며(Richardson et al., 2000; Cabrita et al., 2001), 0.25 mL straw와 같은 소용량 용기는 정자량이 제한적인 어종에 주로 사용된다(Huang et al., 2004). 정자동결보존시 straw의 용량이 작을수록 해동후 운동성이 높다는 결과가 무지개송어(
이상의 결과를 종합해 보면, 4배체 참굴 정자동결보존시 적정 동해방지제 및 농도는 10% DMSO이며, 평형시간은 30분, 냉각속도는 분당 5℃, straw 크기는 0.25 mL로 나타났다. 그러나 이때의 해동후 생존율 및 SAI는 각각 18.4% 및 0.46으로 2배체 참굴 정자의 해동후와 비교해 볼 때 4배 정도 낮은 것으로 비교되었다. 이러한 이유는 4배체 참굴 정자가 2배체보다 그 크기가 커므로 4배체 정자에서는 동결보존시 동해방지제 침투가 원활하지 않아 동해를 많이 받았기 때문인 것으로 추정되며, 추후 이에 대한 현미경적 관찰이 필요할 것으로 보인다. 앞으로 4배체 정자의 해동후 생존율 및 운동성을 향상시키기 위해서는 다양한 동해방지제를 대상으로 그 농도, 평형시간, 냉각 및 해동 속도에 대한 연구가 더 많이 이루어져야 할 것으로 사료된다.