근해어업의 발달과 연안오염의 가중, 양식어업의 증가 등으로 연안어장의 생산성이 저하되면서, 우리나라의 연근해 어업의 총어획량은 1980년대 이후 감소하고 있는 실정이다(Fujita, 1987; Yotsui and Maesako, 1993; MIFFAF, 2008). 이러한 수산자원 감소를 방지하기 위한 여러 방안 중에 바다목장 사업은 수산자원의 방류에서 어획에 이르기까지 자연친화적으로 관리하는 어업생산시스템으로 인공어초, 해조장 등 인위적인 시설물의 설치를 통해 어패류 자원의 서식공간을 제공하고 수산종묘 방류를 통해 고갈되어 가는 수산자원을 증대시켜 안정적인 수산물 생산 및 공급을 목적으로 한다(MOMAF, 2005; MIFFAF, 2008; Lee et al., 2012). 또한, 바다목장 사업은 어장을 적극적으로 선택하고 조성하는 과정에서 생물자원의 현존량 변화 및 생태특성에 관한 자료를 축척함으로써 효율적인 생태계 관리모델을 개발하는데도 그 목적이 있다.
울진바다목장에서는 어류 및 패류의 자원량을 증대시킬 목적으로 조피볼락(
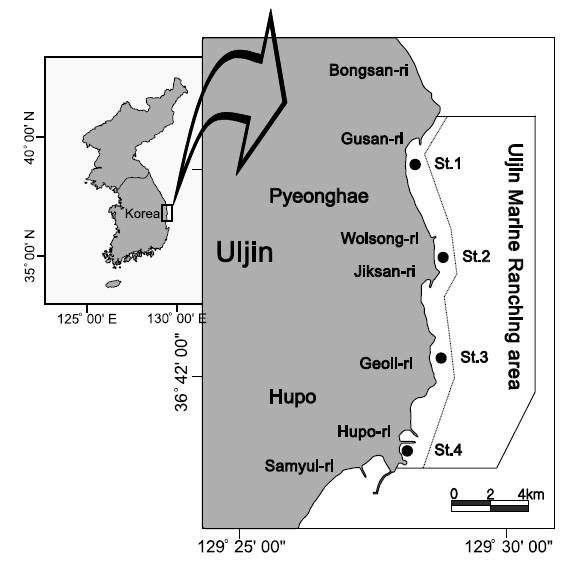
울진바다목장 해역은 경북 울진군 근남면 구산리에서 후포면 후포리 일대에 20 km2 면적으로 조성되어 있는데, 거일리를 중심으로 남쪽 해안은 암반이 잘 발달되어 있고 직산리, 월송리 및 구산리 일대의 낮은 수심의 퇴적물에는 입자의 크기가 큰 사질(평균 입도 1.5Ø)이 높은 함량을 보이는 특징을 가지고 있다(Paik et al., 2007). 본 조사는 2009년부터 2010년까지 1년에 4회씩(3월, 6월, 9월, 12월) 4개의 정점에서 형망을 이용하여 채집하였다(Fig. 1). 본 조사에서 사용된 형망은 틀의 길이가 140 cm, 갈퀴의 간격은 2 cm, 망목의 크기는 2.5 cm였으며, 예망에 사용된 로프의 길이는 평균 150 m였다. 울진바다목장 해역에서 채집된 저서동물은 각각 개체수와 생체량을 측정하였으며, 가장 우점하여 출현하는 대복과 개량조개(
형망에 의해 채집된 저서동물의 군집의 구조를 이해하기 위하여 출현 종수 및 개체수 자료를 이용하여 종다양도지수(
형망에 의해 채집된 저서동물 군집의 정점별·월별 유사성을 파악하기 위하여 정점별·월별로 출현한 저서동물의 종수(Cij)와 개체수(Si , Sj)를 바탕으로 하는 Bray-curtis 유사도지수(BC; Bray-Curtis, 1957)를 사용하여 유사도를 백분율(%)로 나타내었으며, 유사도분석에서 종수와 개체수 사이의 자료간 편중을 피하기 위아여 fourth root로 자료를 변환하여 사용하였다.
각각의 정점별·월별로 분석된 유사도를 바탕으로 정점별·월별 유사성의 차이를 분석하기 위하여 수지도(dendrogram)를 작성하여 집괴분석(cluster analysis)을 실시하였으며, 그룹간 연결 방식에는 complete linkage 방식을 사용하였다. 군집분석 결과 분리된 정점군과 정점군 사이의 생물군집의 차이를 유발하는 종을 분석하고 이들이 군집의 차이에 기여하는 정도를 분석하기 위하여 SIMPER 분석을 실시하였으며, 이들의 분석에는 PRIMER V5.0을 사용하여 실시하였다(Clarke and Warwick, 2001).
시간의 변화에 따른 저서동물의 종수, 개체수, 생체량, 종다양도와 종우점도의 변화를 분석하기 위해서 SPSS version 12.0 for Windows로 paired t-test를 실시하였고, 정범별로 2009년과 2010년 체장분포의 차이를 분석하기 위해서는 t-test를 실시하였다. 또한 정점별·월별로 출현한 종수, 개체수, 생체량, 종다양도, 종우점도 및 우점종의 각장의 차이를 분석하기 위해서 ANOVA test를 실시하였으며 사후분석으로 Turkey-T 분석을 사용하였다.
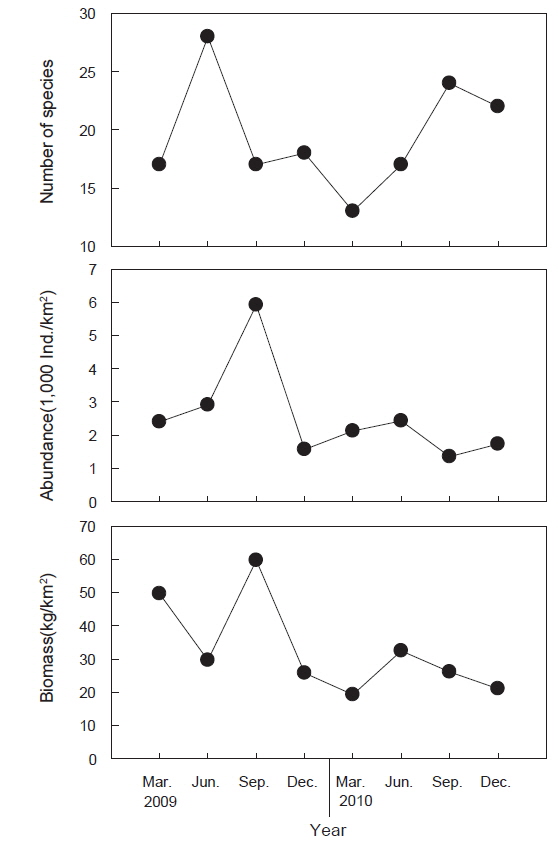
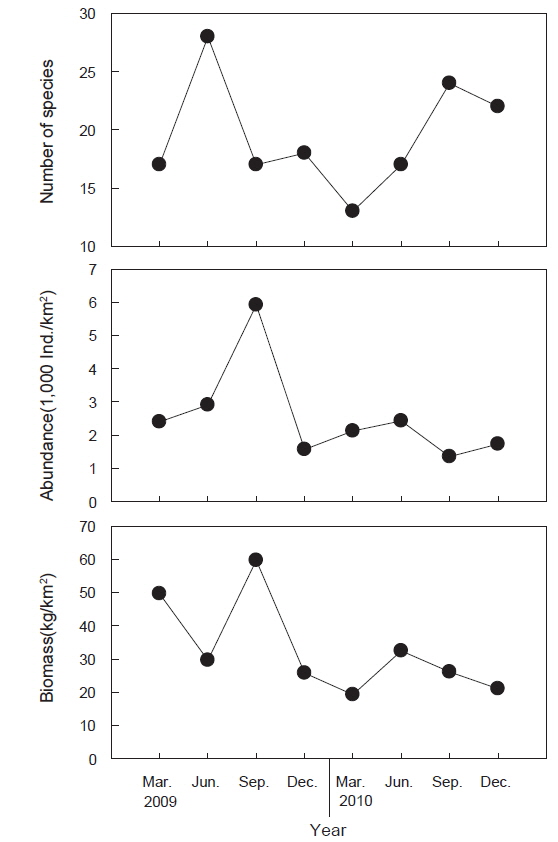
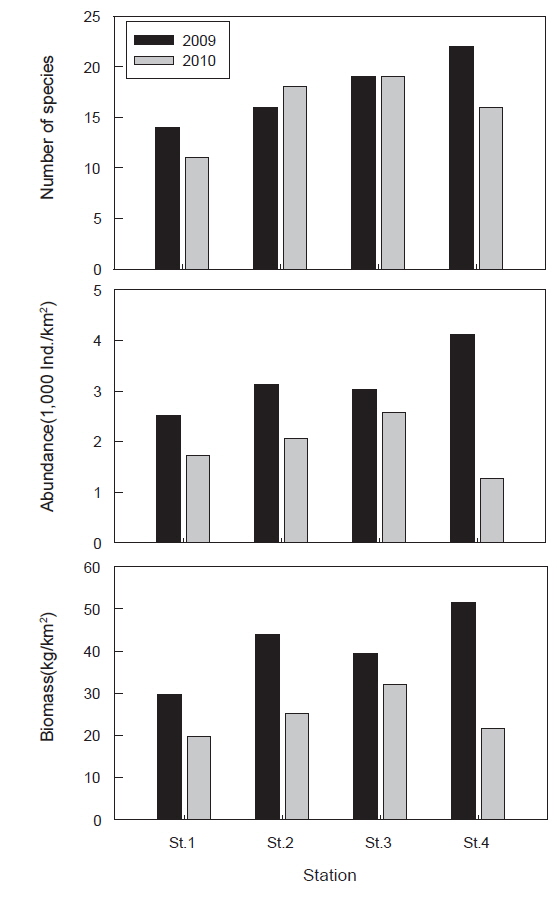
조사 기간 동안(2009-2010년) 울진바다목장 해역 4개 정점에서 출현한 저서동물은 총 41종으로 연체동물이 18종으로 가장 많았고, 절지동물 12종, 어류 6종, 극피동물 3종, 해면동물과 환형동물이 각각 1종이 출현하였다(Fig. 2). 시기별 출현 양상에 있어서는 2009년에 총 36종이 출현하였으며 연체동물이 15종으로 가장 많았고 절지동물 11종, 어류가 6종의 순이었다. 2010년에는 총 31종이 출현하였으며 연체동물이 14종, 절지동물 8종, 어류 4종의 순이었으며 년도별 종수의 차이는 없는 것으로 나타났다(paired t-test,
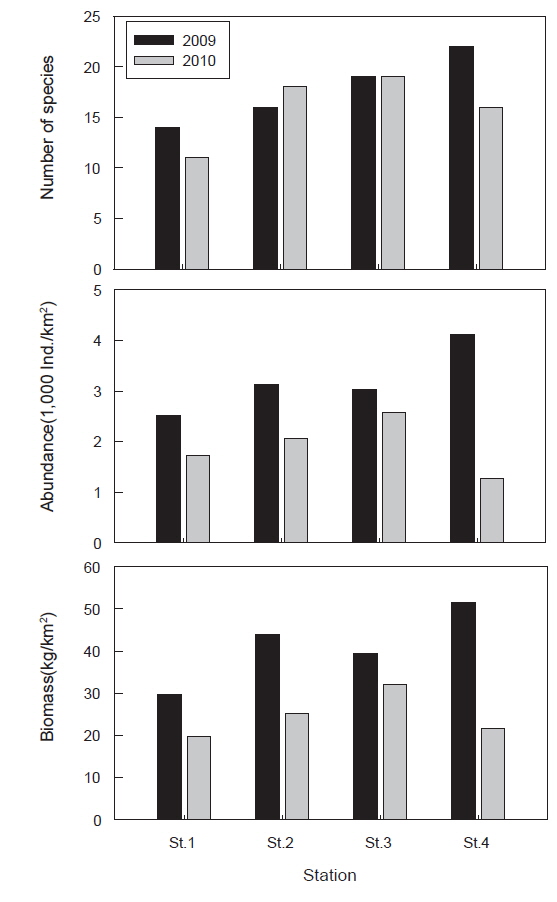
정점별로 출현 종수를 살펴보면 정점 1에서는 총 15종(평균 8.6종), 정점 2에서는 22종(평균 11.0종), 정점 3에서는 24종(평균 9.9종), 정점 4에서는 23종(평균 11.1종)이 출현하였으며, 정점 사이에는 유의한 차이를 보이지 않았다(Fig. 3, ANOVA,
조사기간 동안 출현한 저서동물의 평균 개체수는 2,554,633 ind./km2이었는데, 2009년에는 평균 3,198,793 ind./km2, 2010년에는 1,910,473 ind./km2로 2009년에 비해 2010년에 개체수가 감소한 것으로 나타났다(Fig. 2, paired t-test,
정점별로 출현한 개체수를 살펴보면 정점 1에서는 평균 2,125,134 ind./km2, 정점 2에서는 2,602,969 ind./km2, 정점 3에서는 2,882,125 ind./km2, 정점 4에서는 2,688,304 ind./km2로 나타났으며 정점간 유의한 차이를 보이지 않았다(Fig. 3, ANOVA,
조사기간 동안 출현한 저서동물의 평균 생체량은 32,946.4 kg/km2로 2009년에는 평균 41,176.6 kg/km2이었고 2010년에는 24,716.3 kg/km2로 2009년에 비해 2010년에 생체량이 감소한 것으로 나타났다(Fig. 2, paired t-test,
정점별로 출현한 생체량을 살펴보면 정점 1에서는 평균 24,817.8 kg/km2, 정점 2에서는 34,557.4 kg/km2, 정점 3에서는 35,827.1 kg/km2, 정점 4에서는 36,583.4 kg/km2로 나타났으며 정점간 유의한 차이를 보이지 않았다(Fig. 3, ANOVA,
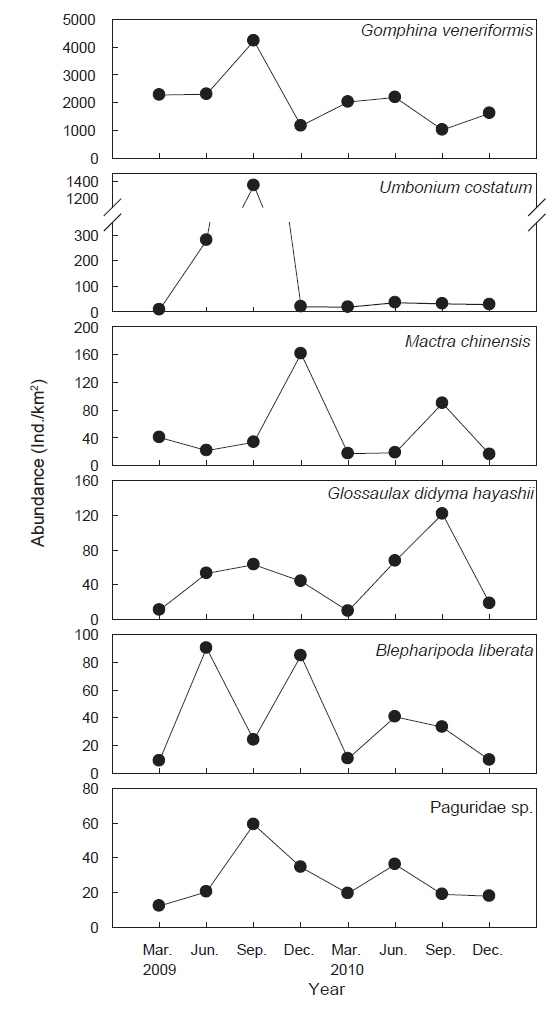
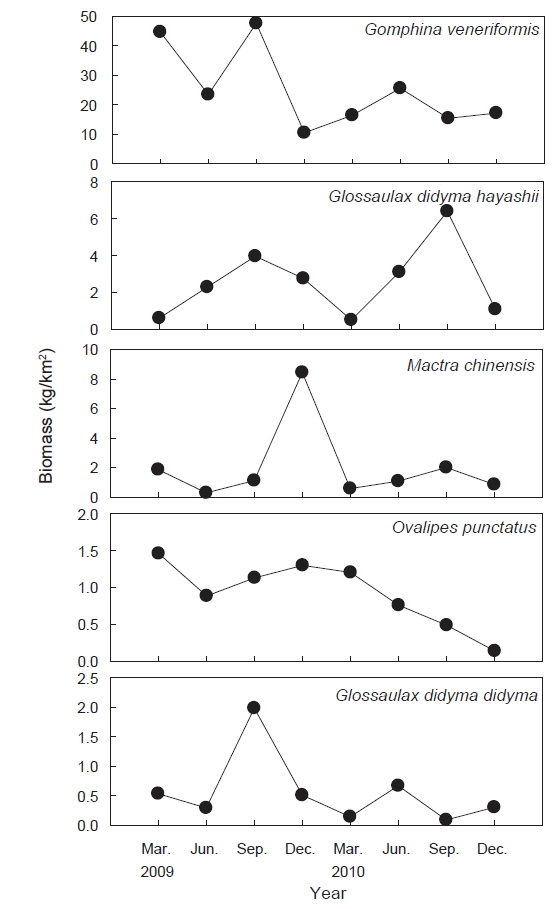
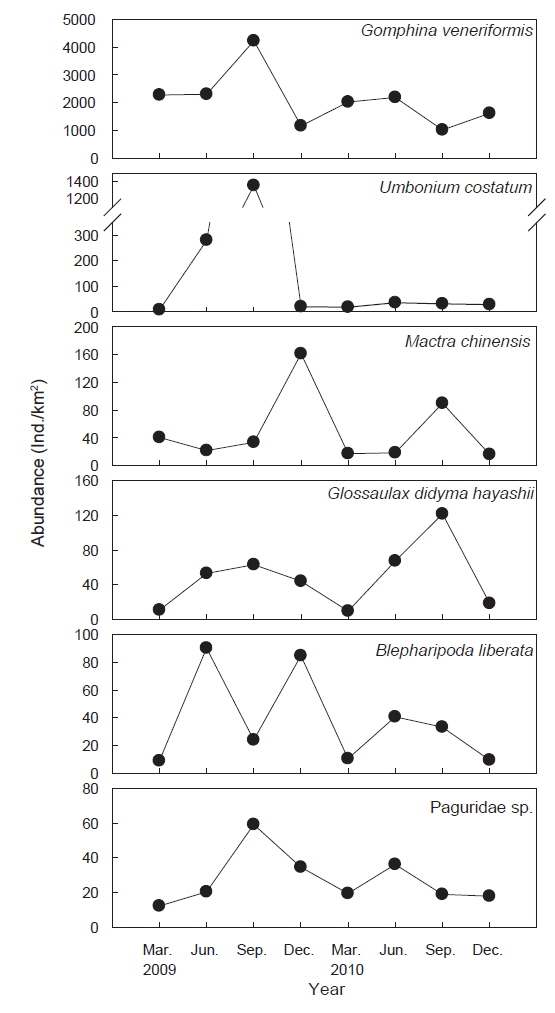
조사기간 동안 채집된 생물들 중에서 전체 개체수의 1% 이상을 차지하는 주요 우점종은 총 6종으로 연체동물이 4종, 절지동물이 2종으로 나타났으며 이들 개체수의 합은 전체 개체수의 97.3%를 차지했다. 대복은 평균 2,491,818 ind./km2(82.2%)로 출현한 저서동물 중에서 가장 높은 개체수를 보이며 극우점했으며, 다음으로는 비단고둥(
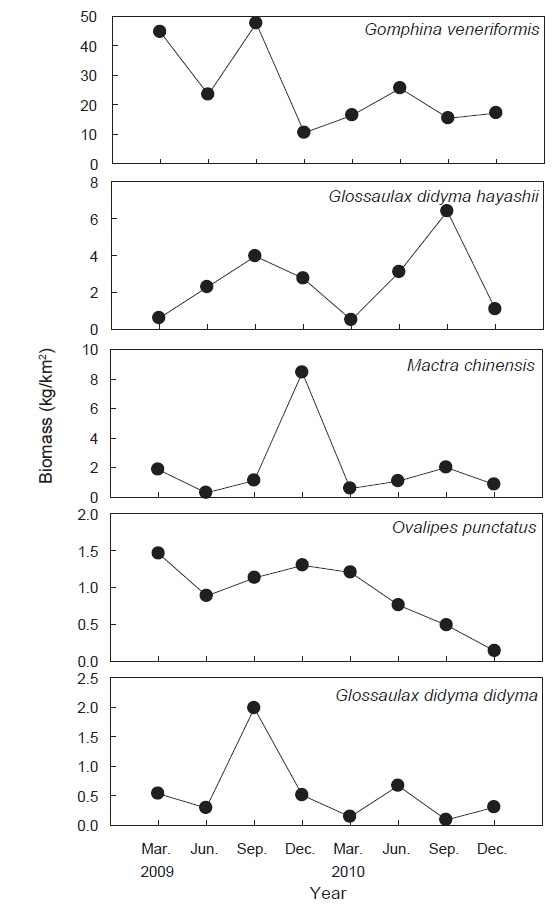
조사기간 동안 채집된 생물들 중에서 전체 생체량의 1.0% 이상을 차지하는 주요 우점종을 분석한 결과 총 5종으로 연체동물 4종과 절지동물 1종으로 이들 생체량의 합은 전체 생체량의 94.8%를 차지했다. 대복은 평균 25,124 kg/km2(76.3%)로 가장 높은 생체량을 보이며 극우점하는 것으로 나타났고, 높은탑이 색구슬우렁이는 평균 2,596 kg/km2(7.9%), 개량조개는 2,024 kg/km2(6.1%) 깨다시꽃게(
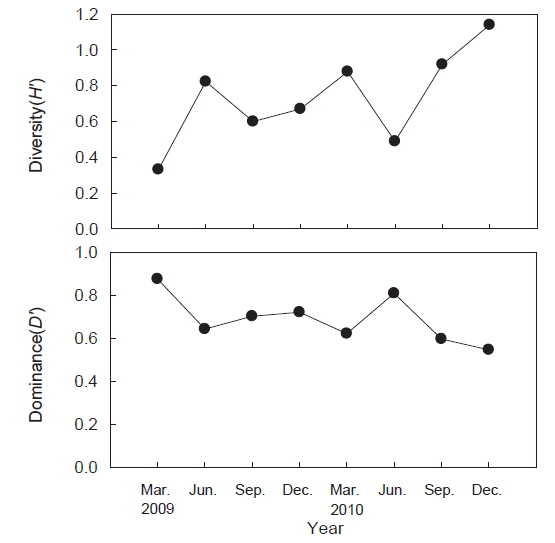
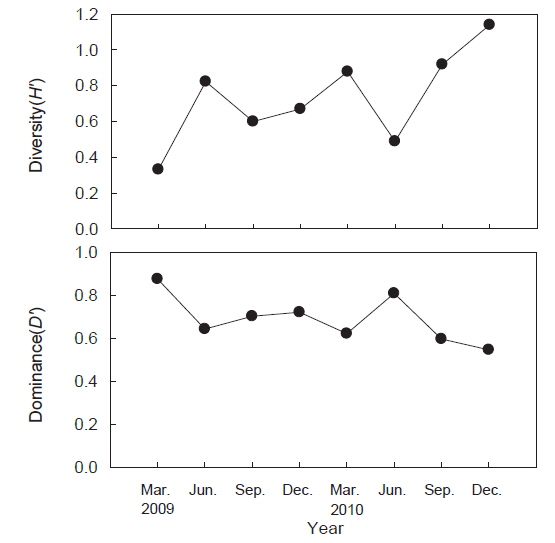
조사기간 동안 출현한 종수와 개체수를 바탕으로 계절별, 정점별 생태지수를 분석한 결과 종다양도(
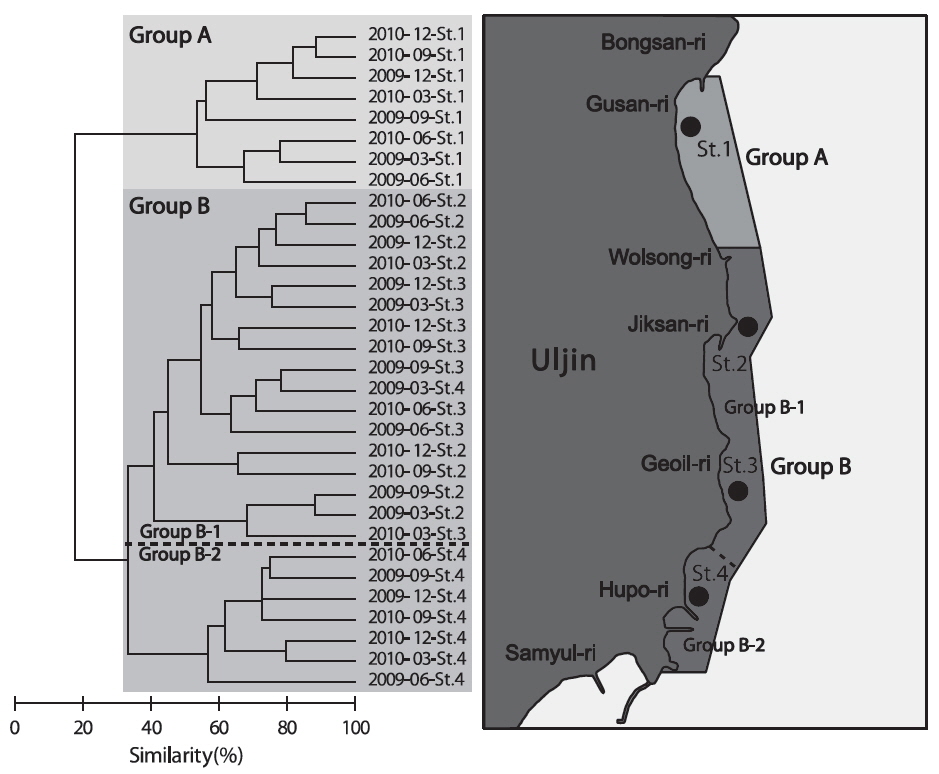
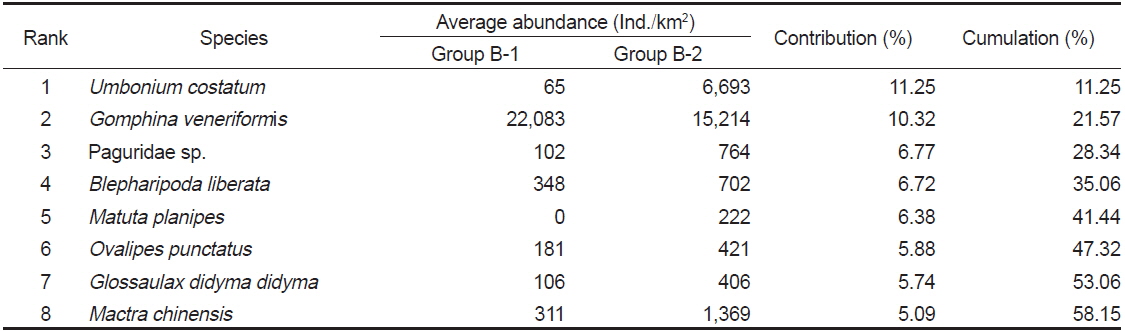
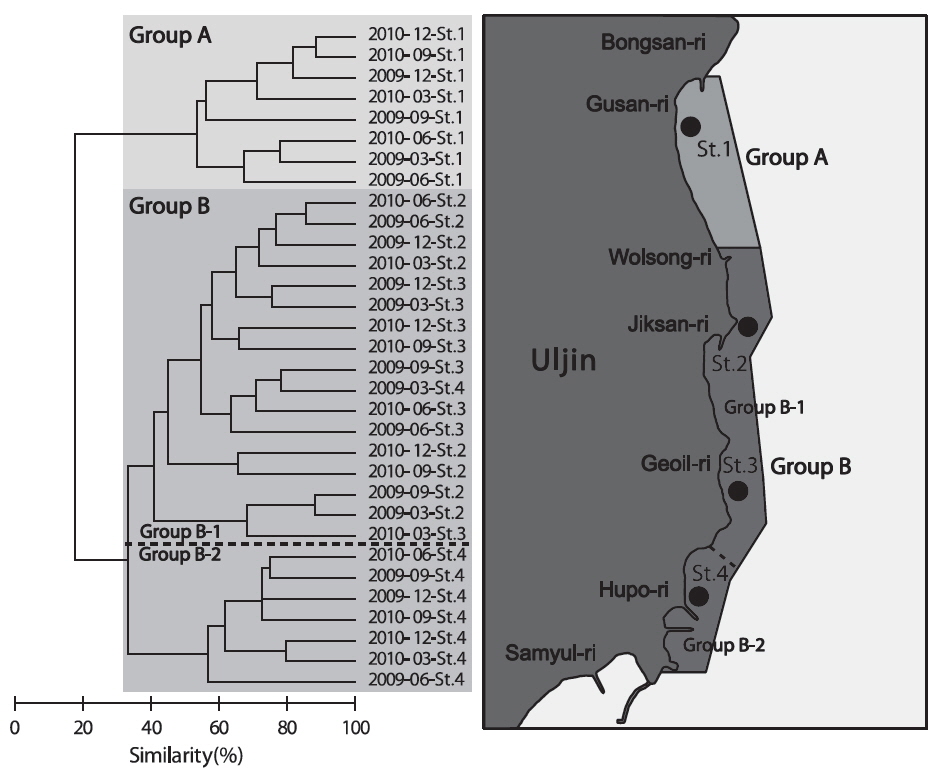
조사기간 동안 계절별, 월별로 출현한 저서동물의 종수와 개체수를 바탕으로 계절별, 월별 유사도를 구하고 집괴분석을 실시하여 생물군집의 유사성을 분석한 결과 생물군집은 크게 2개의 Group으로 구분되는 특징을 보였다(Fig. 7). Group A는 2009년과 2010년에 정점 1에서 조사된 군집이였고, Group B는 2009년과 2010년도에 정점2-4에서 조사된 군집으로 나타났는데, Group B는 다시 2009년과 2010년도에 정점2와 정점 3에서 조사된 군집(Group B-1)과 2009년과 2010년에 정점 4에서 조사된 군집(Group B-2)로 구분되었다.
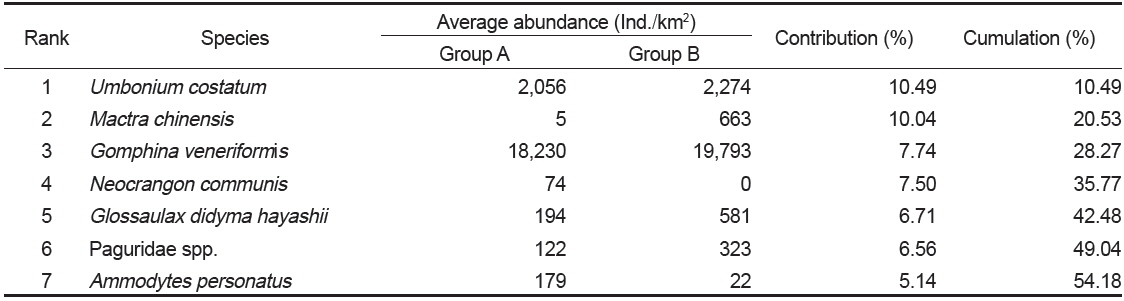
군집분석에서 Group A와 Group B 군집의 차이에 대한 종기여도를 분석하기 위하여 SIMPER분석을 실시한 결과 종기여도에서 5% 이상을 차지하는 종들은 총 7종으로 나타났다(Table 1). 비단고둥의 비유사도에 대한 종기여도는 10.49%로 Group A에서는 2009년 6월과 9월에 각각 2,295 ind./km2와 14,153 ind./km2가 출현하였을 뿐 다른 조사에서는 전혀 출현하지 않았던 반면 Group B에서는 총 24회의 조사 중에서 21회 출현하였으며 개체수는 평균 2,274 ind./km2로 나타나 그룹별 큰 차이를 보였다. 개량조개는 Group A에서는 평균 5 ind./km2로 매우 낮은 개체수를 보였던 반면 Group B에서는 평균 663 ind./km2의 개체수를 보이며 10.04%의 종기여도를 나타냈다. 또한 높은탑 이색구슬우렁이와 집게류는 Group A에 비해 Group B에서 높은 개체수를 보였고, 반면 두가시자주새우(
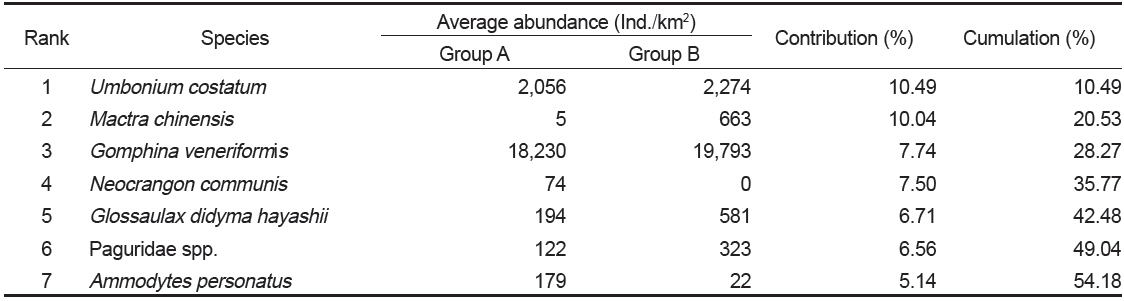
Species contributions to dissimilarity between Group A and Group B in the Uljin marine ranching area from 2009 to 2010
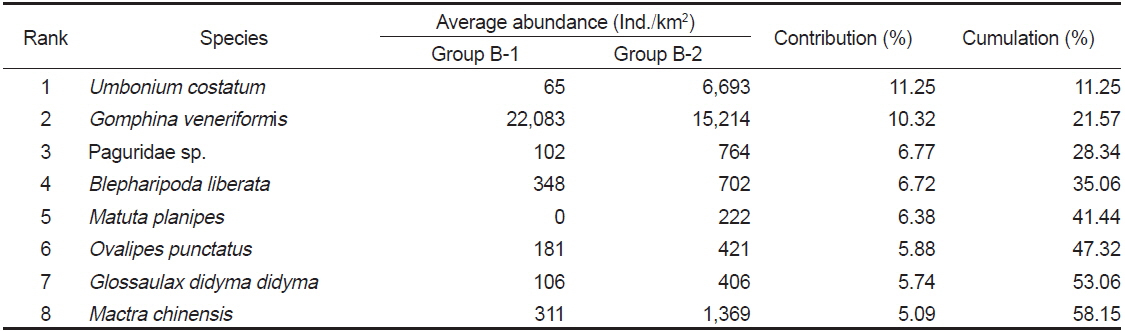
Species contributions to dissimilarity between Group B-1 and Group B-2 in the Uljin marine ranching area from 2009 to 2010
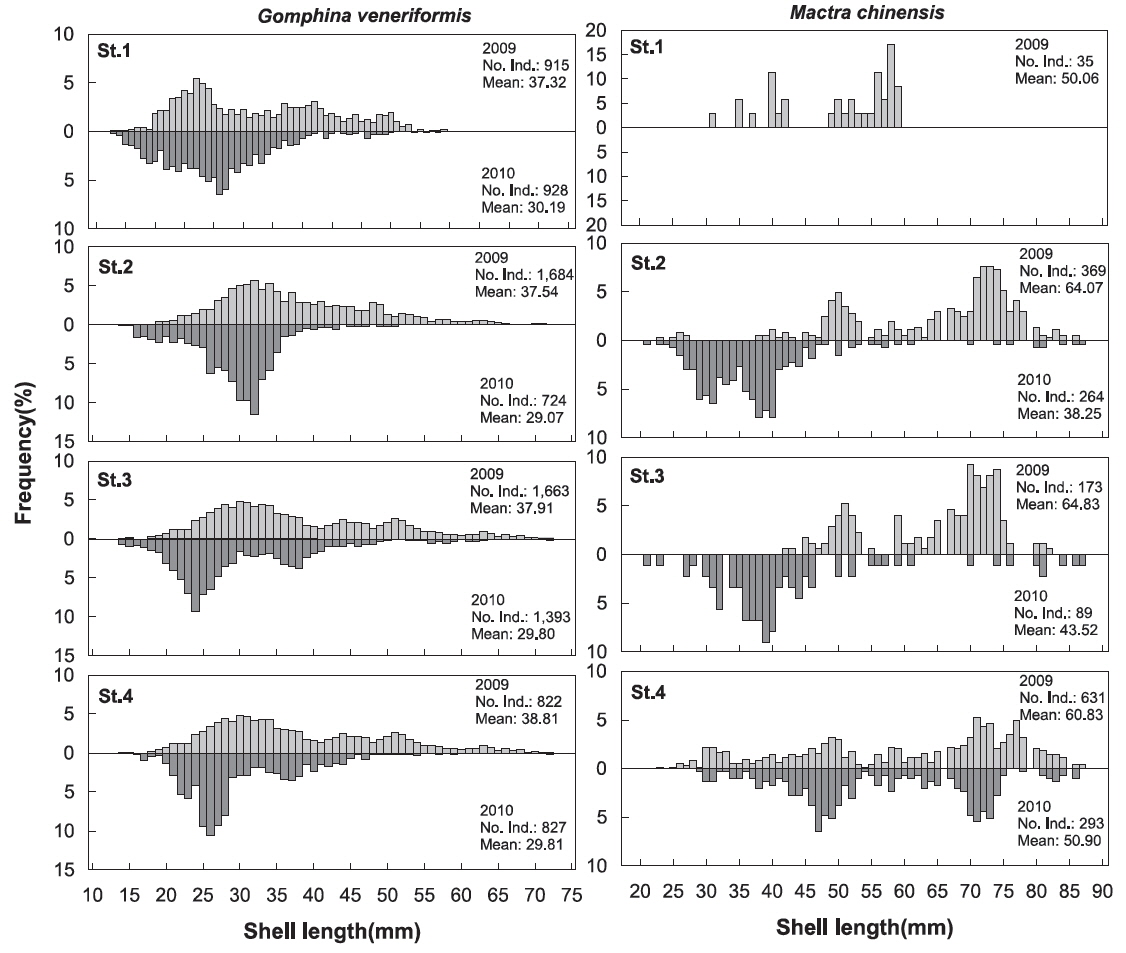
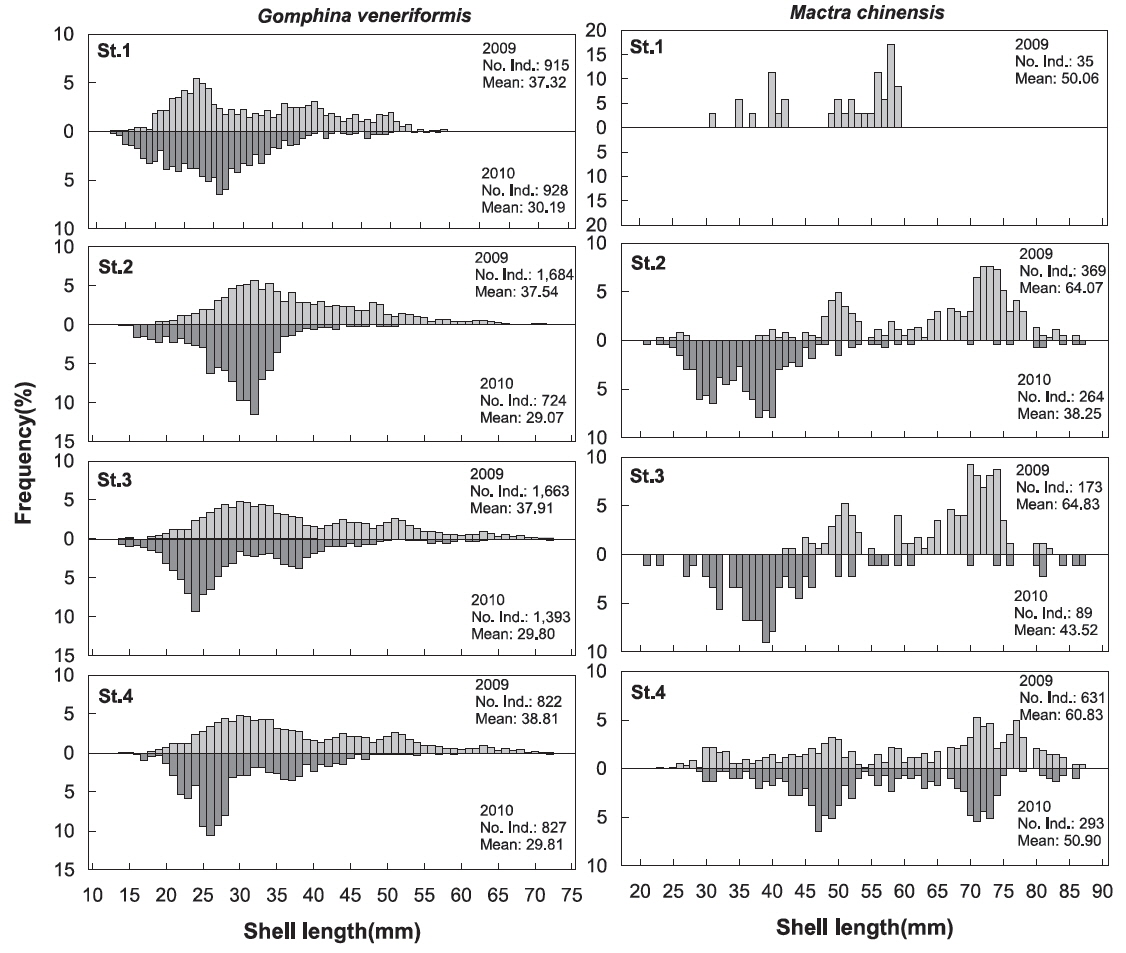
대복의 각장분포를 살펴본 결과 2009년에 평균 37.73 mm, 2010년에 평균 29.76 mm로 평균 각장이 감소한 것으로 나타났는데(t-test,
개량조개의 각장분포를 살펴본 결과 2009년에 평균 62.03 mm, 2010년에 평균 44.94 mm로 평균 각장이 감소한 것으로 나타났는데(t-test,
울진바다목장 해역 4개 정점에서 2009년부터 2010년까지 출현한 저서동물은 총 41종으로 연체동물 18종, 절지동물 12종, 어류 6종, 극피동물 3종, 기타 2종으로 연체동물의 비율이 높은 것으로 나타났는데, 본 조사 해역의 경우 수심이 10 m보다 얕고 퇴적물 입자의 크기가 큰 사질해안으로 파도의 영향으로 저층의 교란이 심한 특징을 가지고 있고, 형망의 갈퀴 간격 2 cm, 그물코 2.5 cm로 크기 때문에 다른 분류군에 비해 크기가 작고 부드러운 특징을 가진 환형동물의 출현 종수가 적었다. 연체동물은 단단한 패각을 가지고 절지동물은 키틴질의 갑각을 가지고 있어 다른 분류군에 비해 사질의 함량이 높은 곳에서 높은 종수와 서식밀도를 보이는 것으로 잘 알려져 있는데(Choi et al., 2000; Hwang and Kim, 2003; Ko et al., 2012), 본 조사에서도 다른 분류군에 비해 연체동물과 절지동물의 종수와 개체수가 많은 경향을 보였다. 절지동물 중에서 게류가 많은 종이 출현하였고, 연체동물 또한 크기가 큰 이매패류나 복족류가 주로 출현하는 경향을 보였지만 정점별로 유사한 퇴적물 입도 조성이여서 정점간 종수의 차이는 없었던 것으로 판단된다. 형망을 이용한 저서동물 조사가 적어 직접적인 비교가 어렵지만 상대적으로 크기가 큰 연체동물의 종수를 비교하면 Cha et al. (2012)이 영일만에서 형망을 사용한 이매패류 자원조사에서는 연체동물이 2종, Jung et al. (2013)이 안면도 사질 조간대에서 실시한 조사에서 연체동물은 총 10종이 출현하여 본 조사에 비해 적은 종수가 출현하였다. 반면 Choi et al. (2000)의 강릉 사질해변 조사에서는 연체동물이 24종, Ko et al. (2012)의 제주 사질해변 연구에서는 29종이 출현하여 본 조사에 비해 많은 종수가 나타났는데, 강릉 연안과 제주 사질해변의 조사는 본 조사에 비해 정점수가 많을 뿐 아니라 30 m 수심까지 조사가 이루어져 더 많은 종이 출현했던 것으로 판단된다. 또한 울진바다목장 해역에서 2007년(MIFFAF, 2008)과 2008년(MIFFAF, 2009)에 실시한 조사에서는 2007년에 총 25종과 2008년에 40종이 출현하였는데, 연체동물은 각각 12종과 15종으로 본 조사의 18종에 비해 적은 종수를 보였고, 다음으로 많은 종수를 보였던 어류는 2007년과 2008년에 각각 7종과 10종으로 본 조사의 6종에 비해서는 높은 종수를 보였다. 반면 절지동물은 2007년과 2008년에 각각 4종과 5종으로 적았던 반면 본 조사에서는 12종이 출현하여 상대적으로 절지동물의 종수가 늘어난 것으로 나타났다.
조사기간 동안 출현한 저서동물의 평균 개체수는 시간이 지남에 따라 감소하는 경향을 나타냈는데, 이는 연체동물 중에서 상업적으로 이용되는 대복과 개량조개 개체수의 급격한 감소가 원인으로서 산란기가 끝나고 본격적으로 어획이 이루어지는 8월 이후부터 9월까지 과도한 어획으로 인하여 크기가 큰 개체들이 감소하였기 때문으로 판단된다. 평균 생체량은 2009년에 비해 2010년에 감소한 것으로 나타났는데, 대복과 개량조개의 개체수와 평균 각장이 줄어들면서 연체동물의 생체량이 급격히 감소하여 전체 생체량이 감소한 것으로 판단된다. 정점별 개체수와 생체량의 변화를 살펴보면 모든 정점들에서 대복과 개량조개의 개체수와 생체량은 감소했고 또한 비교적 큰 개체의 높은탑이색구슬우렁이, 깨다시꽃게와 큰구슬우렁이 등의 개체수와 서식밀도도 같이 감소하여 형망어업에 의해 상업적으로 이용되는 종들의 전체 자원량이 감소한 것으로 판단된다.
조사기간 동안 가장 높은 서식밀도와 생체량을 보였던 대복은 평균 2,491,818 ind./km2 (82.2%)의 서식밀도와 25,124 kg/km2(76.3%)의 생체량을 보였는데, 이러한 생체량은 울진바다목장에서 2007년과 2008년에 조사된 평균 생체량 56,556 kg/km2 과 43,839 kg/km2 에 비해 생체량이 감소하는 경향을 나타냈다(MIFFAF, 2008; 2009). 2007년과 2008년의 조사에서 대복은 전체 어획량의 95% 이상을 차지할 정도로 극우점하는 경향을 보였지만 본 조사에서는 본 조사에서는 2009년과 2010년에 각각 전체 생체량의 76.7%와 75.6%를 차지하여 우점비율이 낮아지는 경향을 보였다. 비단고둥은 평균 221,973 ind./km2(8.7%)로 개체수에서는 많은 비중을 차지했지만 생체량은 평균 282 kg/km2 (0.9%)로 나타났는데, 이 종의 경우 2007년과 2008년의 조사에서는 출현하지 않았던 종으로 급격하게 서식밀도가 증가했다. 비단고둥은 퇴적물의 표면에 반잠입하여 움직이면서 퇴적물에 함유된 유기물을 섭식하는 종으로 일반적으로 퇴적물의 교란이 적고 고운모래의 함량이 높은 곳에서 서식밀도가 높은 것으로 알려져 있는데(Choi et al., 2000; Yoon et al., 2011), 울진바다목장의 경우 후포연안에서 방파제 공사를 실시하고 2007년부터 연안의 얕은 수심에 인공어초 등을 설치하여(MIFFAF, 2008; 2009) 연안의 퇴적물 교란이 적어지고 인공어초에서 탈락되어 연안으로 유입되는 해조류의 조각들이 먹이원으로 작용하면서 본 연구지역에서 비단고둥의 개체수가 증가한 것으로 판단된다. 개량조개는 평균 49,879 ind./km2(2.0%)와 2,024 kg/km2(6.1%)로 우점하는 경향을 보였는데 어획량의 경우 2009년과 2010년에는 각각 2,925 kg/km2, 1,123 kg/km2을 보였지만 2007년과 2008년의 조사에서는 전체의 1% 미만으로 매우 적었던 것에 비하면 생체량이 크게 증가한 것으로 나타났다. 높은탑이색구슬우렁이와 큰구슬우렁이는 평균 49,879 ind./km2(1.9%)와 17,650 ind./km2(0.7%)의 서식밀도를 보였으며, 생체량은 각각 평균 2,596 kg/km2(7.9%)와 567 kg/km2(1.7%)을 보였는데, 2007년과 2008년에는 이 2종을 모두 큰구슬우렁이로 동정한 것으로 추정되며 이들의 생체량은 각각 평균 2,208 kg/km2과 988 kg/km2로 본 조사에서 생체량이 더 높은 것으로 나타났다. 절지동물중 게가재는 평균 37,714 ind./km2(1.5%)와 201 ind./km2(0.6%), 집게류는 평균 27,266 ind./km2(1.1%)와 195 ind./km2(0.6%), 깨다시꽃게는 22,310 ind./km2(0.9%), 921 kg/km2(2.8%) 등이 우점하는 것으로 나타났는데, 이 종들의 경우 2007년과 2008년에는 거의 출현하지 않거나 매우 적었지만 2009년부터 개체수와 생체량이 증가한 경향을 나타냈다(MIFFAF, 2008; 2009).
군집분석에서 분리된 Group A와 Group B 의 군집은 비단고둥의 분포에서 가장 큰 차이를 보였는데, Group A에서는 2009년 6월과 9월에 각각 2,295 ind./km2와 14,153 ind./km2로 2번만 출현했던 반면 Group B에서는 총 24회의 조사 중에서 21회 출현하였으며 평균 2,274 ind./km2의 서식밀도를 보였다. 이는 비단고둥의 주서식지는 Group B 해역이지만 해류와 퇴적물의 이동 등의 영향으로 북쪽으로 이동하면서 채집된 것으로 판단된다. 또한 개량조개, 높은탑이색구슬우렁이와 집게류는 Group A에 비해 Group B 해역에서 높은 서식밀도를 보여 두 해역의 군집구조에 영향을 미치는 것으로 나타났는데, Group A 해역의 경우 북쪽과 5 -15 m의 수심에 암반이 넓게 발달해 있어 상대적으로 암반이 적은 Group B 해역에 비해 퇴적물의 교란이 심하기 때문에 상대적으로 Group B 해역에서 높은 서식밀도를 보였던 것으로 판단된다. 이러한 경향은 Group B-1과 Group B-2 해역의 분리에서도 유사하게 나타나는데, Group B-1 해역은 최근 바다목장이 조성되면서 인위적으로 많은 인공구조물을 설치되여 해변에서 모래가 유실되어 밖으로 이동하고 인공구조물에서 생기는 해류의 변화 등이 퇴적물을 교란하여 상대적으로 Group B-2 해역에 비해 생태교란이 심한 환경으로 인하여 비단고둥, 집게류, 게가재, 그물무늬금게, 깨다시꽃게, 큰구슬우렁이 등 대부분의 종들이 Group B-2 해역에서 높은 서식밀도를 보이는 것으로 나타났다.
본 조사지역에서 대표적인 상업종인 대복과 개량조개의 각장분포를 살펴본 결과 대복은 2009년에 평균 37.73 mm, 2010년에 평균 29.76 mm로 평균 각장이 감소하였고, 개량조개 또한 2009년에 평균 62.03 mm, 2010년에 평균 44.94 mm 로 평균 각장이 감소한 것으로 나타났다. 대복의 군성숙 각장은 Lee et al. (1999)은 25.00 mm, Park et al. (2003)은 31.10 mm에서 모두 성숙한다고 하였고, Kim (2013)은 암컷의 경우 27.71 mm로 보고하였는데, 2007년과 2008년의 조사(MIFFAF, 2008; 2009)에서 2007년에는 평균 40.79 mm, 2008년에는 46.86 mm로 50%성숙각장인 28.00 mm 미만의 크기는 전체 어획량에서 평균 10% 이하로 매우 낮은 비율을 보였지만 본 조사해역에서 2009년에는 20.9%, 2010년에는 50.8%로 미성숙 개체의 비율이 크게 증가하면서 재생산에 참여하는 개체수가 감소하는 경향을 보였다. 대복은 Lim (2009)에 따르면 1세에 28.33 mm, 2세에 37.75 mm까지 성장한다고 하였고, Ahn (2009)은 1세에 27.01 mm, 2세에 37.76 mm까지 성장한다고 하였는데, 본 조사해역에서는 2009년과 2010년에 걸쳐 미성숙 개체들의 남획이 이루어지고 있어 재생산에 참여하는 개체수의 감소로 인하여 자원량의 감소가 예상된다. 이러한 경향은 개량조개에서도 뚜렷하게 나타났는데, Park and Zhang (2008)은 부산 동리지역에서 개량조개의 적정어획 개시 연령을 5세(51.94 mm)로 제시하였는데, 본 연구에서 2009년에는 5세 미만의 개체들이 전체의 33.0%를 차지했지만 2010년에는 전체의 57.2%로 증가하는 경향을 보여 개량조개의 자원량의 유지를 위해서는 형망틀의 간격과 그물코의 크기를 조정하거나 어린 개체의 어획을 자제해야 할 것으로 판단된다.
울진바다목장 해역에서 저서동물의 군집구조를 살펴본 결과 이전의 2007년과 2008년에 비해서 극우점했던 대복의 서식밀도가 감소하면서 소형 갑각류와 기타 연체동물의 종수와 개체수가 증가하는 경향을 보였다. 특히 인공어초와 같은 인공구조물이 설치된 지역과 방파제 안쪽에 위치하여 상대적으로 환경교란이 비교적 적은 해역에서 절지동물과 연체동물의 종수와 개체수가 증가하는 경향을 보여 인공어초 등에서 유입되는 새로운 먹이원 가입과 적합한 퇴적환경이 소형 갑각류와 기타 연체동물의 종수와 개체수의 증가에 영향을 미친 것으로 판단된다. 반면 상업적으로 이용되는 대복은 미성숙개체의 비율이 2009년 20.9%에서 2010년에 50.8%로 증가하고, 개량조개의 적정어획개시 크기보다 작은 개체의 비율은 2009년에 33.0%에서 2010년에 57.2%로 모두 증가하는 경향을 보여 미성숙개체에 대한 남획이 이루어지는 것으로 대복과 개량조개의 자원량의 관리를 위해서는 대복은 100% 성숙각장인 30.00 mm 이상의 개체, 개량조개는 적정어업개시 크기인 52.00 mm 이상의 개체들을 어획하는 것이 필요하다고 판단된다.