연안은 육상생태계와 해양생태계가 만나는 전이지대로서 다양한 해양생물들이 서식하는 중요한 서식지 중의 하나이다. 해양의 일차 생산자인 해조류는 물질순환을 주도할 뿐 만 아니라, 해양생물의 산란장, 서식지로 활용되며 그 자체로서 생물자원, 바이오에너지원, 산업용, 식품용 및 먹이원으로 이용된다(Choi et al., 2008).
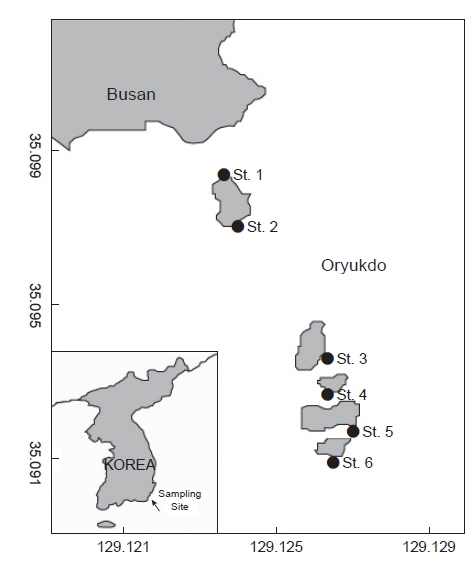
오륙도는 부산 연안에 위치하여 상징성이 높은 무인도서로서, 전형적인 수직 암반 조간대와 조하대로 구성된 연안과 매우 가까운 섬으로 6개의 돌섬이 나란히 뻗어 있는 형태를 갖는 도서이다(Kim et al., 2012). 각 섬은 간격이 좁으며 주변 해역은 30 m 내외의 낮은 수심을 형성하며 남쪽에서 북쪽으로 흐르는 해류의 영향을 받는다. 최근 우리나라 연안은 임해공단 건설에 따른 공업화, 도시화에 따른 주거단지 건설, 각종 해양 사고의 증가 등으로 인해 각종 기능이 임해지역으로 집중되고 연안 개발사업이 추진되고 있다(Choi, 2007). 부산항 및 부산만의 개발이 발표되고 진행되는 시점에서 부산지역 조간대 및 조하대 연안의 오염도는 날로 심각해질 것이며, 오륙도 해역 역시 직간접적으로 오염원에 노출된 섬으로 부산 북항과 각종 대규모 연안개발, 부산항 준설사업, 대단지 건설사업 등에 의한 영향으로 최근 생태계의 건전성이 위협받고 있으며 서식 해양생물의 종소성과 생물량도 감소할 것으로 판단된다.
해양생물의 종조성은 인위적인 요인과 자연적인 요인에 의해서 변화가 일어날 수 있으며(Kim et al., 2012), 인위적 요인이 작용하지 않더라도 자연적인 요인에 의해 종조성은 수시로 변할 수 있다. 해양생물 중 해조군집은 서식지 형태를 변화시키는 해양생물들의 존재 여부에 따라 영향을 받을 수 있는데(Thompson et al., 1996; Bates and DeWreede, 2007), 다양한 지역에서 해양 환경은 전 지구적 변화와 함께 지역적 특성도 변화시켜 생태적인 변화를 가속화시킬 수 있다(Russell et al., 2009). 따라서 해양환경 변화에 대한 해양생물의 자연적인 변화 양상과 이러한 변화에 대한 지속적인 생태학적 모니터링 연구가 필요한 시점이다.
Kang (1966)은 해역별 수온과 해류 등 해양의 물리적 환경을 토대로 우리나라 해조류의 지리적 분포를 동해안 북부, 동해안 남부, 남해안, 서해안, 제주도의 5개 해역으로 연안을 나누었다. 부산시 남구에 위치한 오륙도 주변 해역은 동해안 남부와 남해안 동부의 경계 지역에 위치하며 대마난류의 영향을 강하게 받는 곳으로(Kang, 1966), 우리나라 해조류의 지리적 분포를 구획하는데 있어서 매우 중요한 지점으로 여겨진다(Kang et al., 2008). 최근에 진행된 부산시 인근 해역인 동해안 남부 지역에서 수행된 해조류에 관한 연구는 Nam and Kim (1999), Yoo(2003a, 2003b), Choi (2007), Kang et al. (2008), Choi et al. (2010), Yoo et al. (2013) 등 일부 연구에 그쳐 있으며, 조간대를 대상으로 진행된 연구가 대부분이어서 조하대 해조군집에 관한 연구는 매우 빈약한 실정이다.
이 연구는 2003년 12월 31일에 지정된 해양생태계보호구역인 부산시 남구 오륙도 및 주변 조하대 해역에 서식하는 해조류 군집의 특성 및 변동을 명확하게 밝히기 위하여 연구 대상 해역의 계절에 따른 각 수심대별 해조군집의 변화를 조사하였으며, 서식 해조류의 군집구조와 분포 특성 등 시공간적 변화 양상을 파악하기 위하여 정량 및 정성조사를 토대로 연구를 실시하였다.
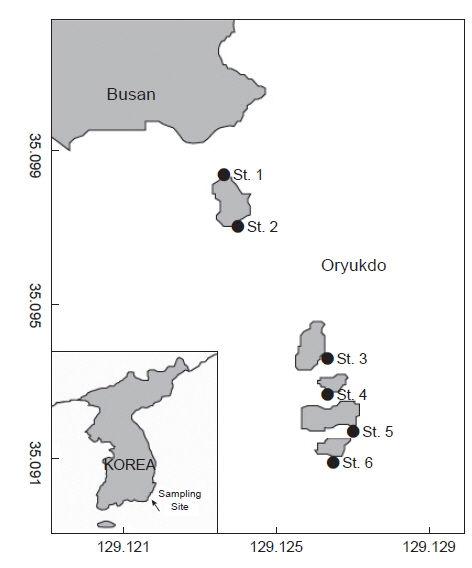
이 조사는 부산시 남구 용호동에 위치한 오륙도 주변 일대에서 조하대 조사지역의 해조류 식생을 대표 할 수 있는 곳을 중심으로 2011년 3월부터 2014년 2월까지 계절별로 1회씩 현장 샘플을 채집하여 해조상 및 군집조사를 실시하였다(Fig. 1).
해조상과 해조군락의 분석을 위한 정량조사는 각 조사 대상 정점에 0.5 m×0.5 m 크기의 방형구를 설치하고 방형구 내의 모든 해조류를 끌칼 등으로 부착기부까지 전량 채집하였다. 채집된 샘플은 신선도를 유지하며 현장에서 포르말린-해수 용액(5-10%)으로 고정하여 실험실로 운반하여 분석에 사용하였다. 또한 조사 정점 주변 기질을 대상으로 현장에 서식하는 해조류를 자세히 조사하면서 정성적으로 채집하였다. 정성채집으로 확보된 샘플은 추후에 종목록을 작성하는데 사용하였다. 현존량은 방형구내의 해조류를 실험실에서 담수로 충분히 씻어 불순물을 제거하여 페이퍼 타올 등으로 습기를 제거한 뒤 습중량을 측정하였으며 단위면적(m2)당 생물량으로 환산하였다.
해조류의 종 동정은 광학현미경과 해부현미경을 이용하였고, 학명과 목록 정리는 한국 해조목록의 분류체계(Kang, 1968; Lee and Kang, 1986, 2002), 일본해조류도감(Yoshida et al., 1995) 및 국내외 형태연구 논문을 참고하였다. 동정된 해조류 학명과 목록은 Lee and Kang (2002)의 분류 체계를 따라 작성하였다.
출현 종 자료를 이용한 각 지역 간의 유사도(similarity)를 파악하기 위하여, 유사도 지수 행렬로부터 각 조사 지역과 출현 종을 연결하는 방법으로 group-average를 이용하였다. 군집분석은 PRIMER (Plymouth Routines Multivariate Ecological Research) computer package를 이용하여 수지도(dendrogram)와 다차원배열(MDS)로 분석하였다(Clarke and Gorley, 2006). 조사 지역 간 해조류 형태적 차이와 생태적 특성 파악을 위한 기능형군 분석은 Littler and Littler (1984)의 6가지 기능형군별 분류형을 사용하여 분석하였다.
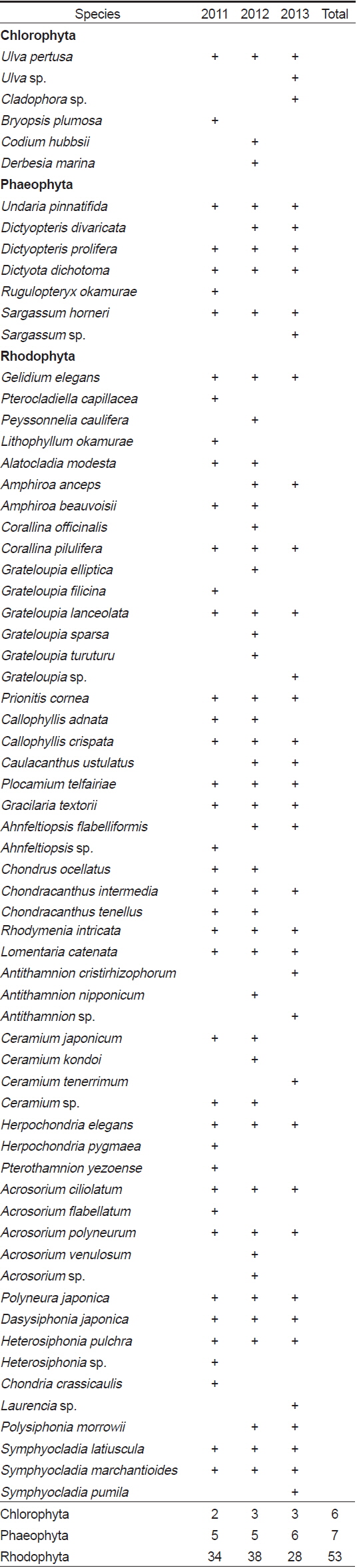
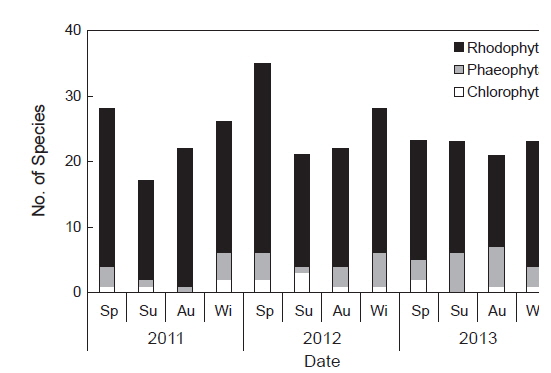
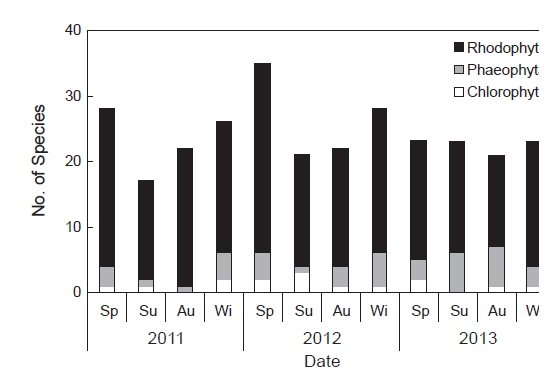
연구기간 동안 총 66종의 해조류가 출현하였으며, 분류군별로는 녹조류 6종(9.1%), 갈조류 7종(10.6%), 홍조류 53종으로 홍조류의 출현 비율이 80.3%로 가장 높았다(Table 1). 각 연구시기별 출현 종 수는 2011년에 녹조류 2종, 갈조류 5종, 홍조류 34종으로 총 41종이 출현하였고, 2012년에는 녹조류 3종, 갈조류 5종, 홍조류 38종으로 총 46종이 출현하여 연구기간 중 가장 많은 종이 출현하였다. 2013년의 경우, 녹조류 3종, 갈조류 6종, 홍조류 28종으로 총 37종이 출현하였으며 연구기간 중 상대적으로 가장 적은 종이 출현하였다. 계절별 출현 종 수는 17종(2011년 여름철)부터 35종(2012년 봄철)까지 연구시기에 따라 다르게 나타났는데, 주로 겨울철부터 봄철에 상대적으로 다른 계절에 비해 다양한 해조류가 출현하였다(Fig. 2).
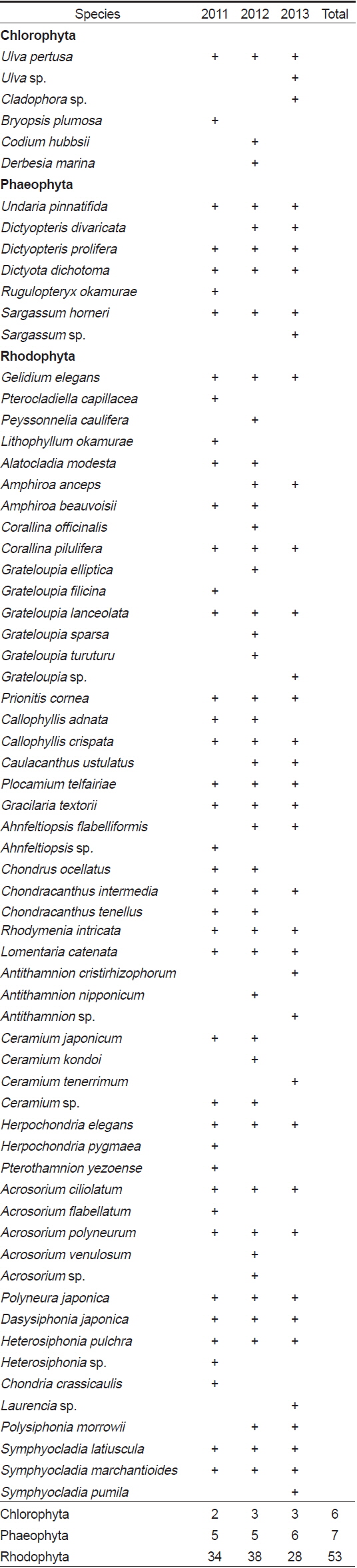
A list of marine algal species investigated at study sites according to the survey period. Sp, Spring; Su, Summer; Au, Autumn; Wi, Winter.
전체 연구기간인 3년 동안 매년 출현한 해조류는 녹조류인 구멍갈파래(
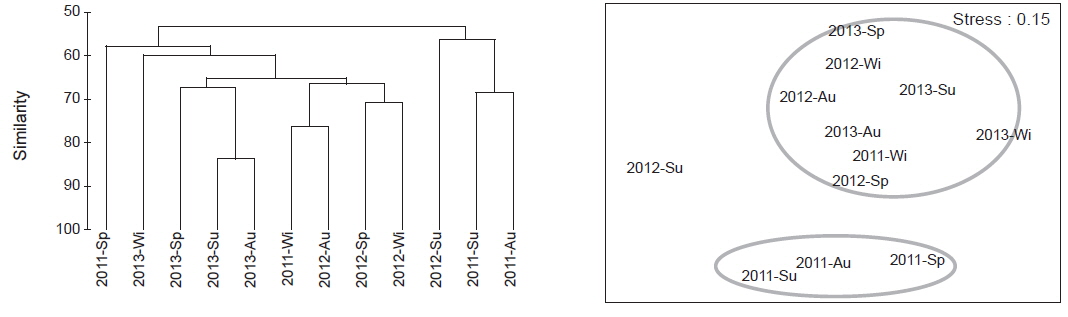
출현 종의 자료를 근거로 해조류의 군집구조 분석을 실시하여 수지도와 다차원배열법으로 분석하였다. 그 결과, 연구시기와 계절에 따라 다소 간의 차이는 있으나, 전반적으로 크게 3개의 그룹으로 구분되었다. 유사도 60% 수준에서 2012년 여름철 출현종과 2011년 봄철, 여름철, 가을철 출현종이 구분되었다. 나머지 한 그룹은 2012년과 2013년의 각 계절별 출현 종이 별개의 그룹으로 나뉘었다(Fig. 3).
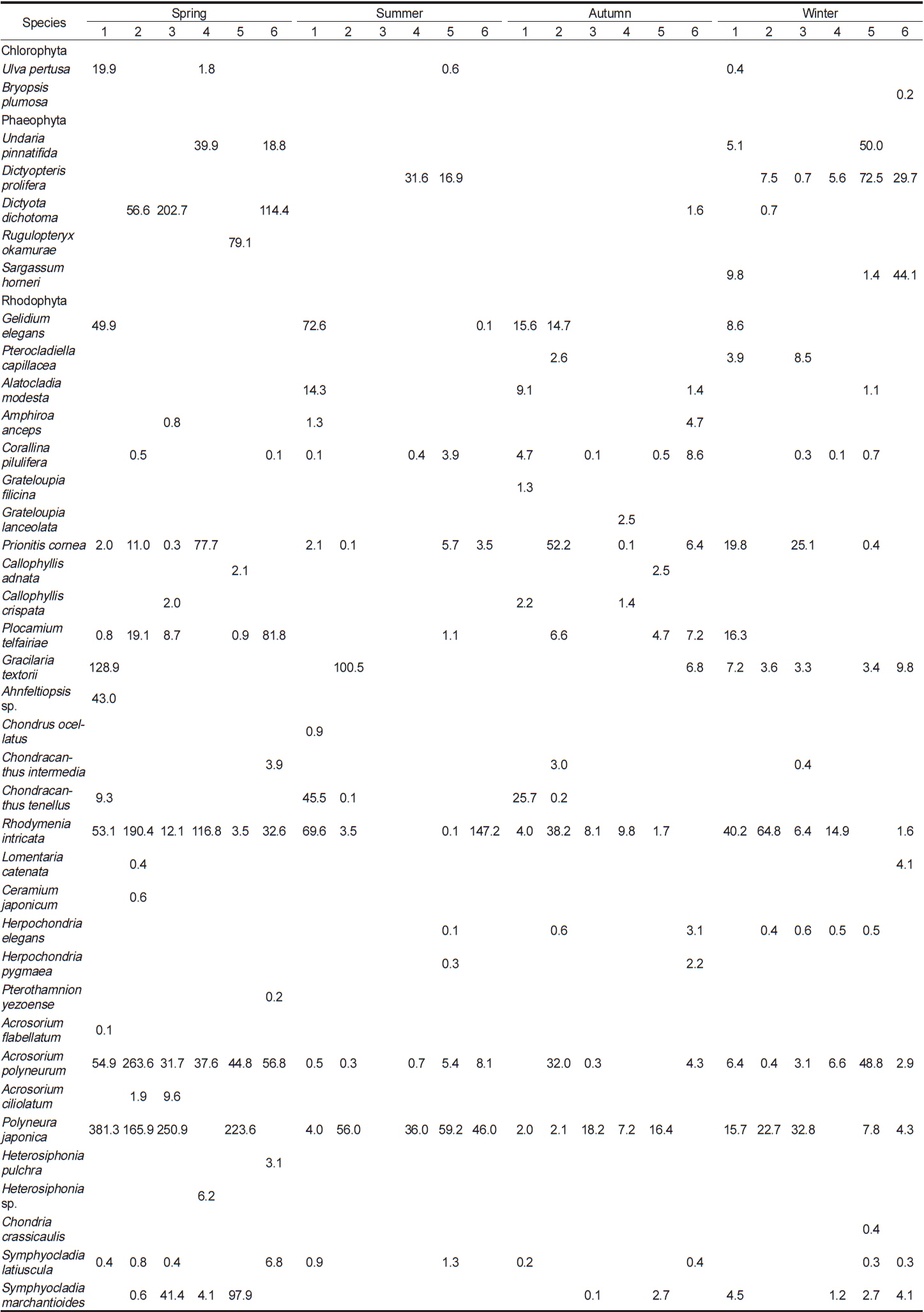
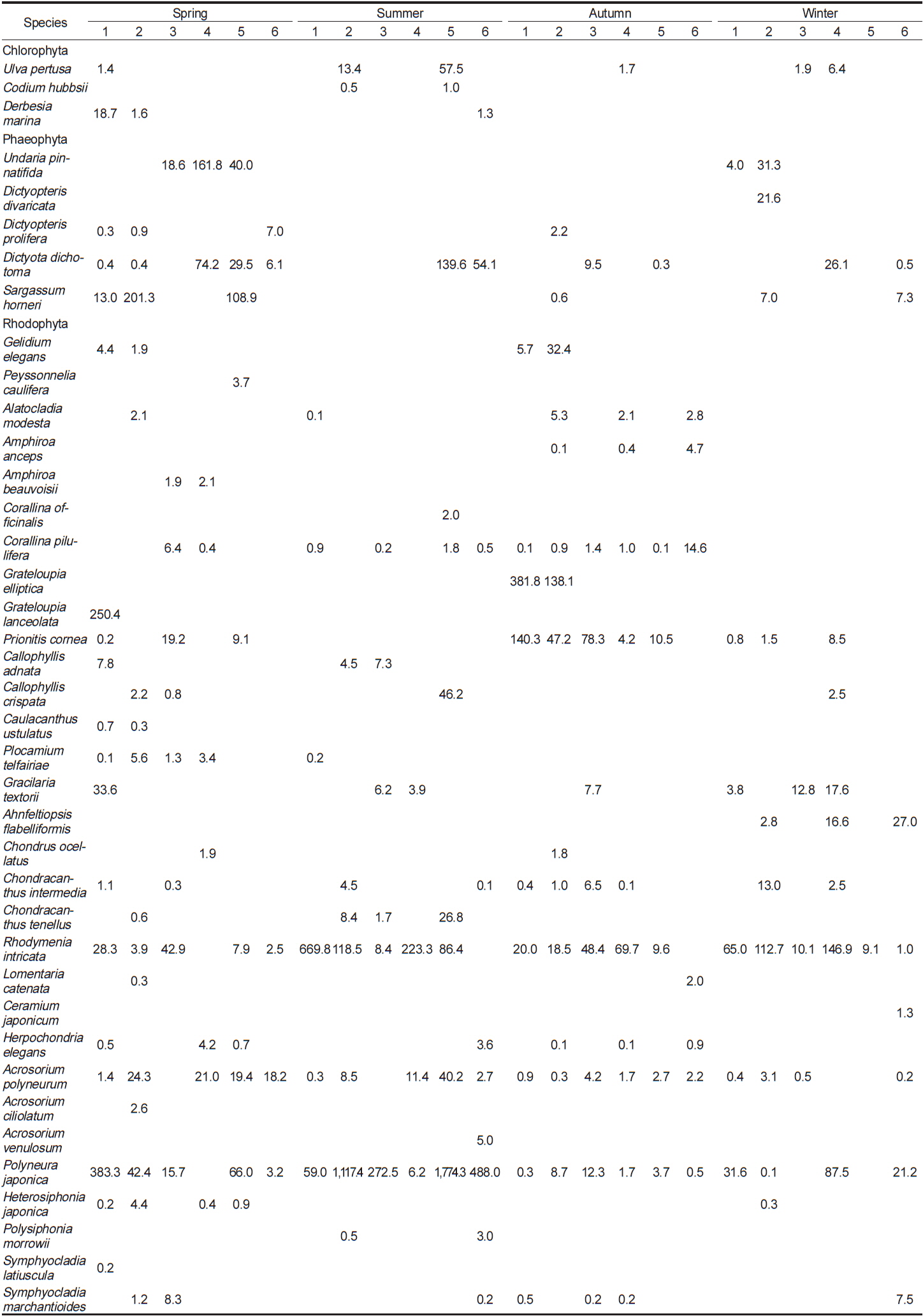
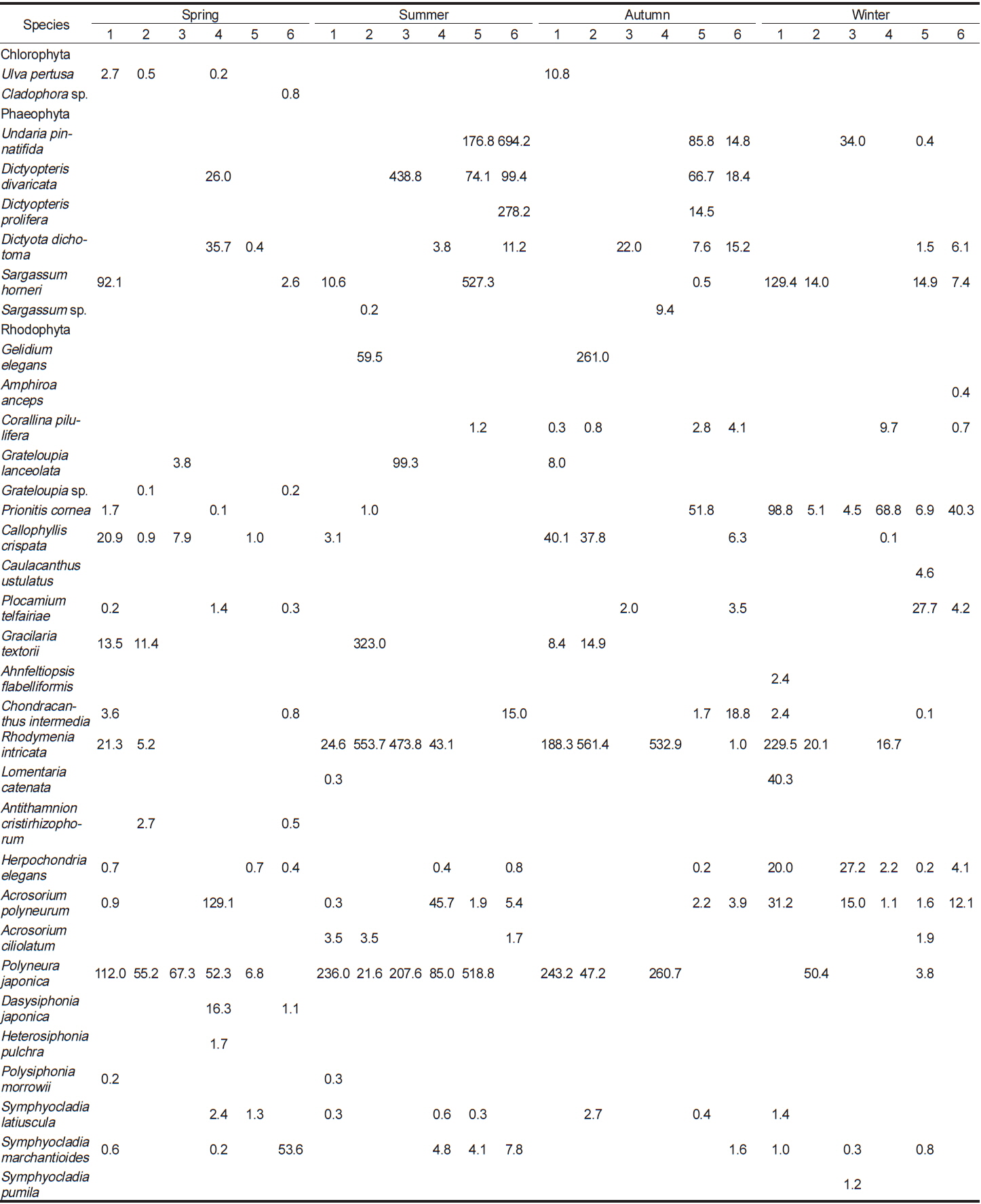
이번 연구를 통하여 분석된 각 정점별 평균 생물량은 Tables 2-4와 같다. 2011년 봄철의 경우, 바다참나무잎아재비(
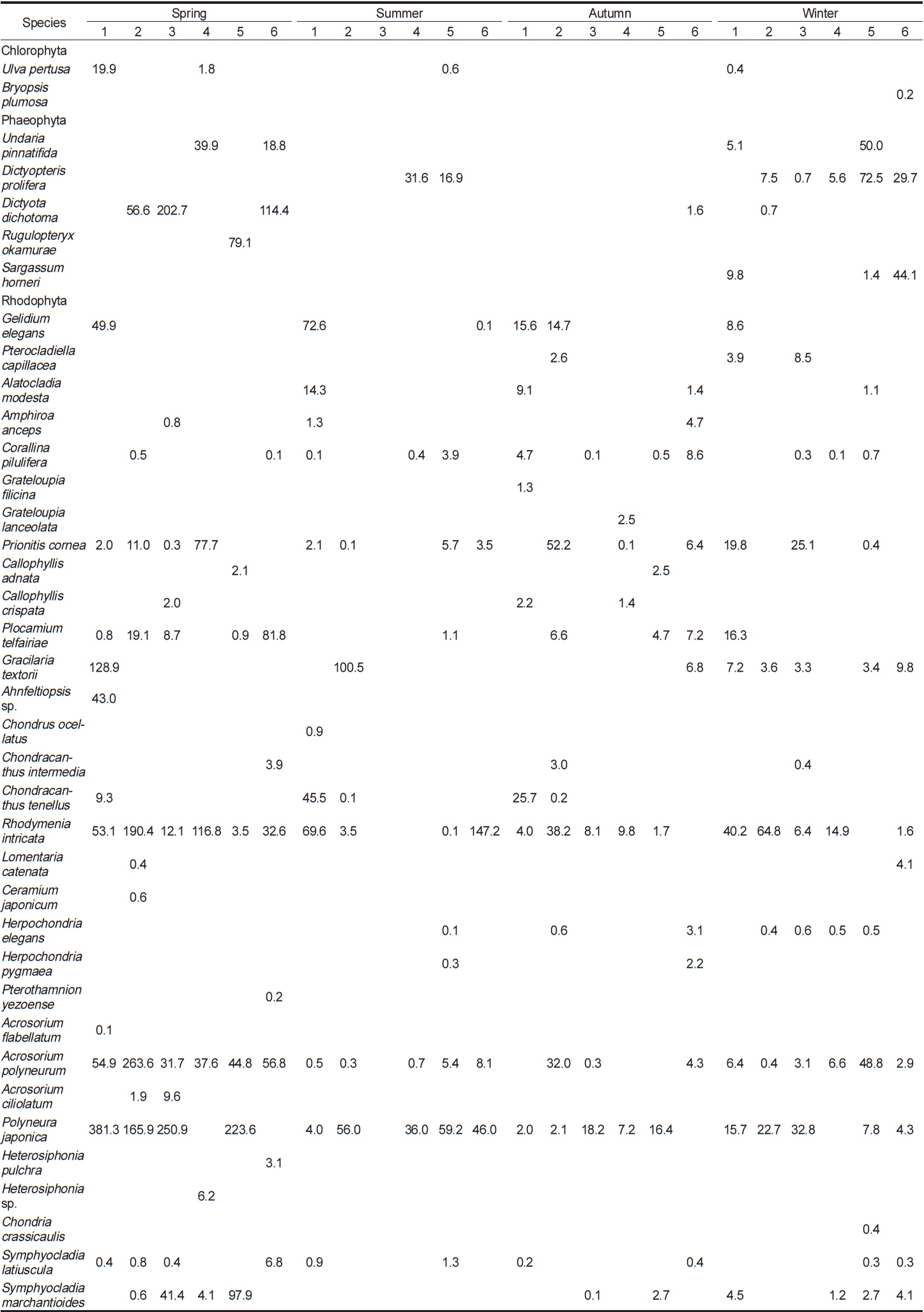
Change of biomass (wet wt. g m?2) of marine algal species investigated at study sites in 2011
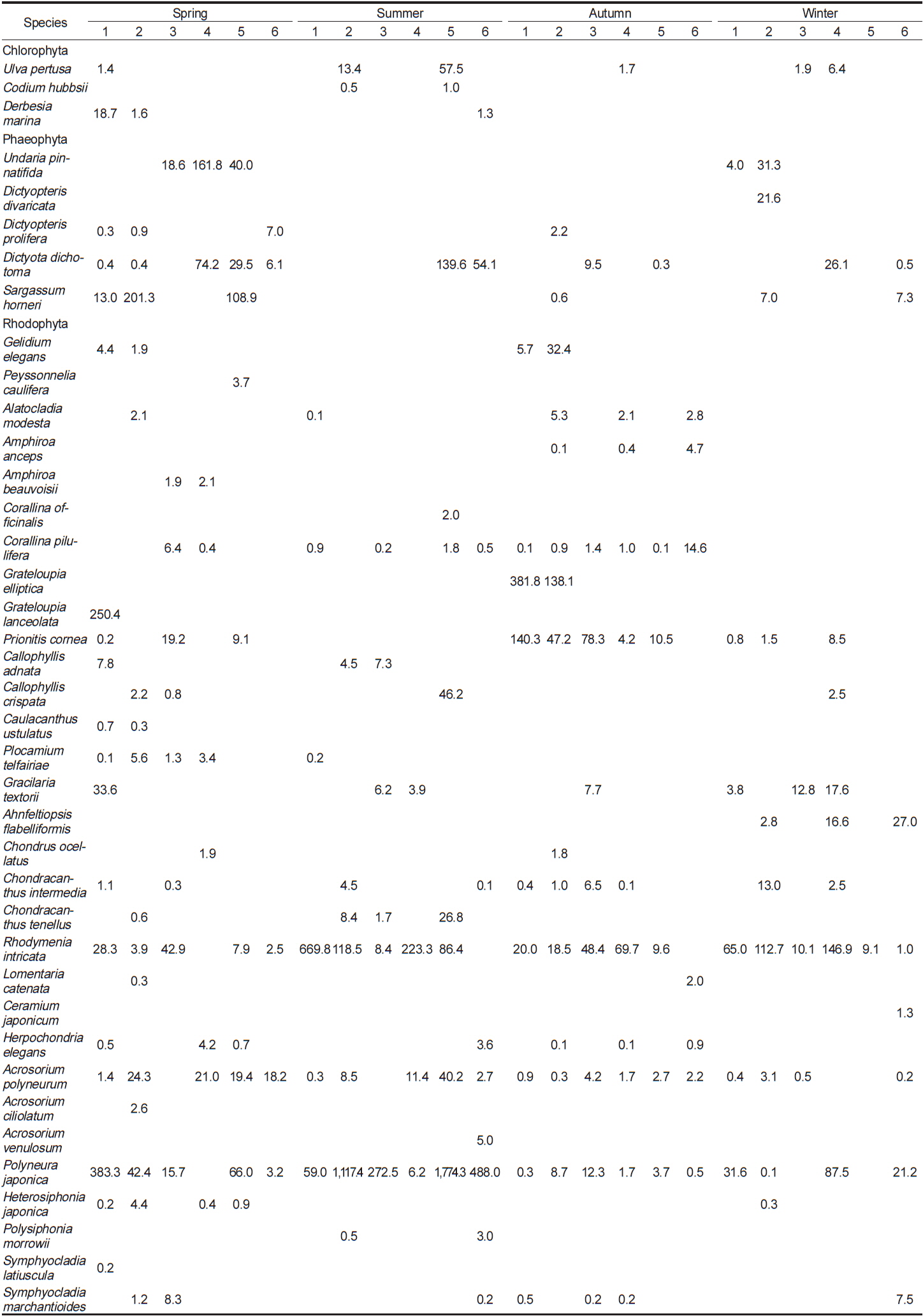
Change of biomass (wet wt. g m?2) of marine algal species investigated at study sites in 2012
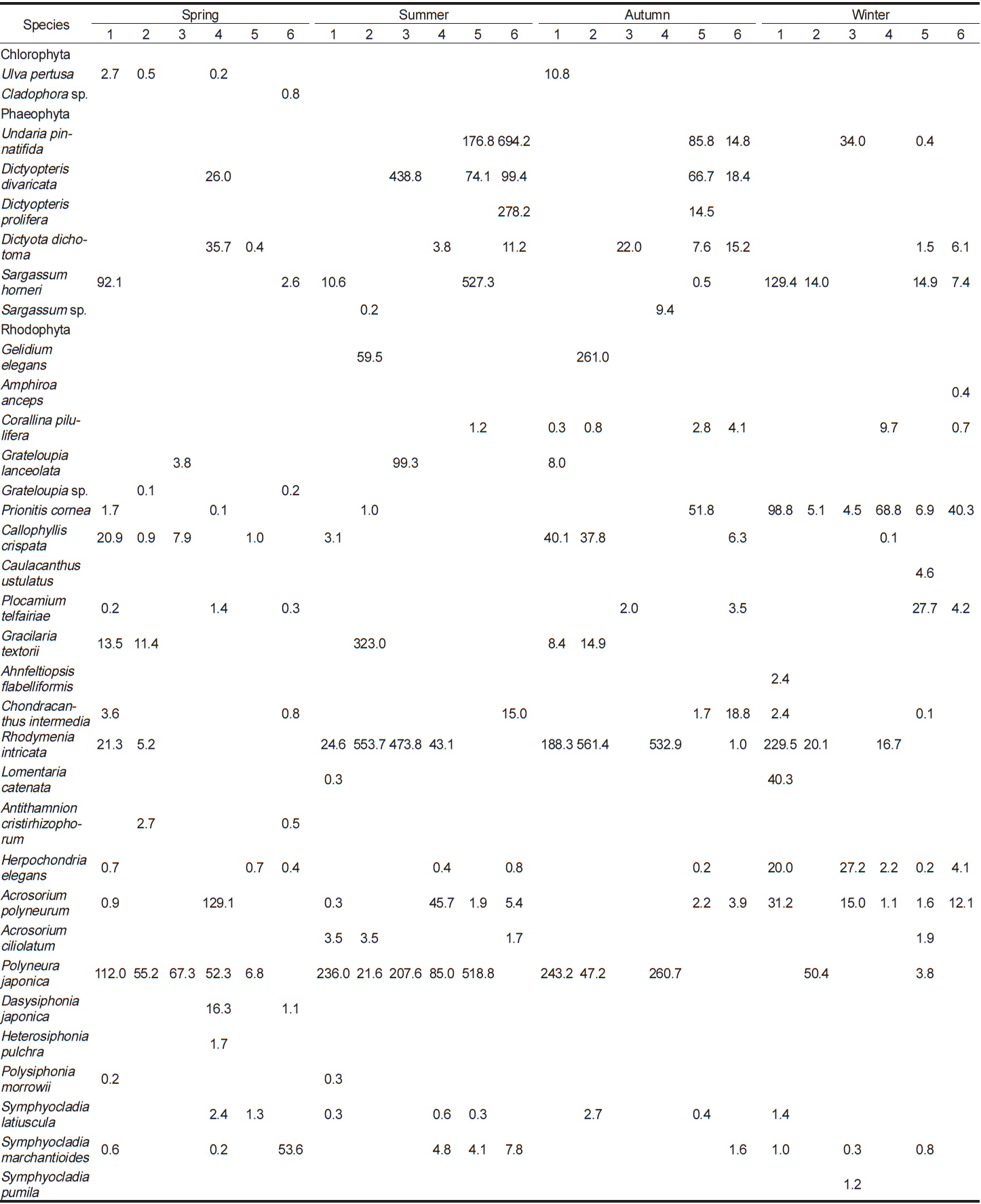
Change of biomass (wet wt. g m?2) of marine algal species investigated at study sites in 2013
전체 연구기간 동안 계절별 연구 정점에 출현한 해조류는 몇몇 종들이 높은 생물량으로 우점하였다. 이들 우점 해조류의 평균 생물량은 연구 시기별로 높게 나타난 경우가 대부분이었으며, 우점하는 해조류의 종 조성에는 큰 차이를 나타내지 않고 유사하였다. 각 정점에서 연구 시기별로 우점하는 해조류는 갈조류인 미역, 가시뼈대그물말, 참그물바탕말, 홍조류인 두갈래분홍치, 갈고리분홍잎, 바다참나무잎아재비 등으로 나타났다.
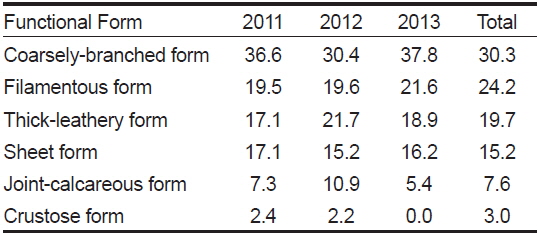
전체 연구기간 동안 각 정점에서 출현한 해조류를 6개의 기능형으로 구분하면, 성긴분기형 30.3%, 사상형 24.2%, 다육질형 19.7%, 엽상형 15.2%, 유절산호말형 7.6%, 각상형 3.0%로 나타났다(Table 5).
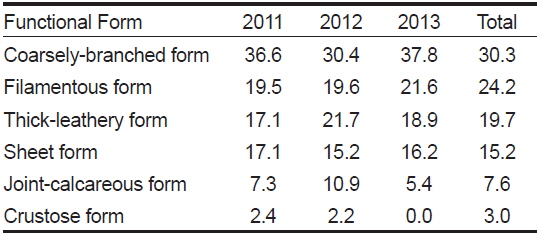
Composition ratio (%) of marine algal functional form group at study sites according to the survey period
각 연구 기간별 기능형의 구성 비율을 살펴보면, 성긴분기형은 30.4-37.8%, 사상형은 19.5-21.6%로 다른 기능형별 해조류에 비해 출현 비율이 높았으며 이들 기능형의 출현 종 수는 대부분 유사하며 높게 나타났다. 출현 해조류의 생태학적 상태그룹(ecological state group, ESG)으로 구분하면, ESG I에 속하는 해조류가 20종(30.3%), ESG II에 속하는 해조류가 46종으로 전체 출현종의 69.7%를 차지했다.
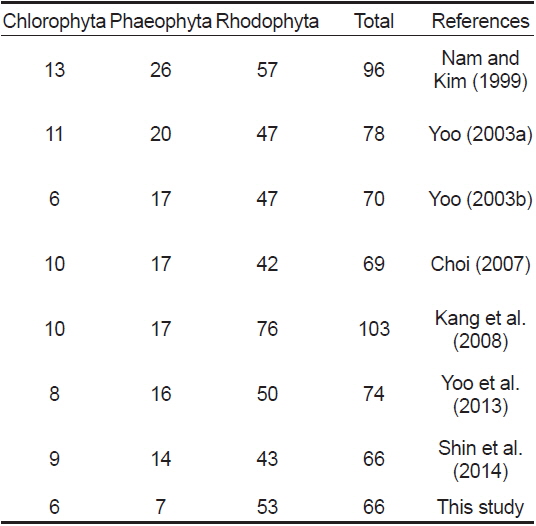
오륙도 주변 해역에서 생육이 확인된 해조류의 분류군별 출현 종 수는 인근 해역인 용호동(Nam and Kim, 1999)과 일광만(Kang et al., 2008)에서 보고된 출현 종 수에 비해 다소 감소한 것으로 나타났지만, 동백섬(Yoo, 2003a), 서암(Yoo, 2003b), 영도(Choi, 2007), 기장(Yoo et al., 2013), 이기대(Shin et al., 2014) 등과 비교시 유사하였다(Table 6). Nam and Kim (1999)은 용호동 주변 해역에서 연중 출현하는 35종의 해조류를 보고하였고, 이들 중 구멍갈파래, 참그물바탕말, 모자반류, 작은구슬산호말, 도박류, 붉은까막살, 애기돌가사리, 마디잘록이, 잔금분홍잎, 참보라색우무 등의 해조류가 이번 연구 결과와 유사한 보편적인 종으로 나타났다. 또한 계절별 출현 종 수에 있어서도 겨울과 봄철에 보다 많은 출현 종 수를 기록하여 이번 연구 결과와 유사한 경향을 보였으며(Fig. 2), 이는 온대해역에 분포하는 해조류의 전형적인 계절적 출현양상을 반영하는 것이라 할 수 있다(Round, 1981). 오륙도 인근에 위치한 영도의 경우(Choi, 2007) 보편적인 해조류 역시 대부분의 종들이 유사하게 나타났고(구멍갈파래, 개그물바탕말, 톳, 우뭇가사리, 작은구슬산호말, 참도박, 붉은까막살, 참곱슬이, 잎꼬시래기, 애기돌가사리 잔금분홍잎, 참보라색우무), 계절별 출현 종 수의 경향도 이번 연구 결과와 매우 비슷한 양상을 나타내 부산 인근 해조상의 보편적 해조류와 출현 경향이 유사한 것으로 연구되었다.
[Table 6.] Marine benthic algal species investigated at vicinity area in previous and this study
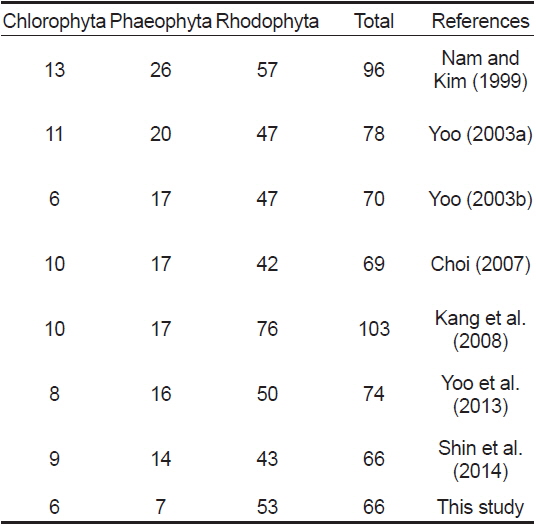
Marine benthic algal species investigated at vicinity area in previous and this study
Yoo (2003a, 2003b)는 동일한 지역에서 시간 경과에 따른 해조상을 분석한 결과, 해조류 출현 종 수와 유사도 지수가 변하는 결과를 해조류 식생이 시간 경과와 함께 변화한다고 제시하였고 이런 결과는 서식지 특성이 영향을 받으며 변화 할 수 있다고 하였다. 이번에 분석된 오륙도의 유사도 및 다차원배열 분석 결과(Fig. 3) 대부분의 정점이 하나로 묶이며 유사도가 매우 높게 분석되었다. 이러한 결과는 오륙도의 모든 정점에 영향을 미치는 염분, 수온, 저질 특성 등의 해양환경이 유사하기 때문에 기인하는 것으로 판단된다. 하지만, 이전 연구 결과가 없기 때문에 시간 변화에 대한 오륙도 서식 해조상 변화 등을 비교, 분석할 수가 없었으며 단지 각 연구 정점간 서식지 특성이 유사할 것이라고 예측할 수 있다.
오륙도 주변 각 연구 정점의 계절별 생물량은 2011년 봄철에 318.5 g m−2에서 743.6 g m−2, 여름철에는 68.7 g m−2에서 211.8 g m−2, 가을철에는 21.0 g m−2에서 152.2 g m−2, 겨울은 28.9 g m−2에서 190.0 g m−2로 겨울과 봄철에 높았다. Yoo et al. (2013)은 기장 주변 해역에서 계절별 생물량이 겨울과 봄철에 높았으며, 봄철의 생물량이 가장 높았다고 보고하여 이번 연구 결과와 유사한 경향을 나타냈다. 생물량의 정량적인 비교시 기장 주변 해역의 생물량이 오륙도에 비해 현저하게 높았는데, 이는 출현종 수는 비슷했지만 주요 우점 해조류가 갈조류 감태(Ecklonia cava)와 모자반류였기 때문으로 분석되었다. 용호동(Nam and Kim, 1999)의 경우, 현존량이 여름(1,648.0 g m−2)철에 가장 높았고 가을, 겨울, 봄으로 갈수록 점차 감소하는 경향을 보였다. 일광만(Kang et al., 2008)에서의 생물량은 겨울철에 629.8 g m−2로 가장 높았고, 가을, 여름, 봄철 순으로 감소하였다. 이는 생물량이 높은 해조류가 구멍갈파래, 붉은까막살, 지충이(
부산 인근 해역에서 생물량으로 본 우점종은 일광만의 경우(Kang et al., 2008), 구멍갈파래, 붉은까막살, 지충이, 개서실, 돌가사리, 외톨개모자반, 괭생이모자반, 개도박 등이었고, 용호동에서는(Nam and Kim, 1999) 구멍갈파래, 톳, 괭생이모자반, 검둥모자반, 지충이, 작은구슬산호말, 잔금분홍잎, 넓은 보라색우무 등으로 나타났다. 이번 연구에서도 높은 생물량은 나타낸 해조류는 갈조류인 미역, 가시뼈대그물말, 참그물바탕말, 홍조류인 두갈래분홍치, 갈고리분홍잎, 바다참나무잎아재비 등으로 오륙도 인근 해역에서 분석된 결과와 유사했으며 대부분 남해안에서 우점하는(Sohn, 1987) 종들과 큰 차이를 보이지 않았다.
우리나라 서해안과 동해안에 출현하는 해조류의 기능형군별 구성비는 성긴분기형과 사상형의 비율이 다른 기능형군의 비율에 비해서 높고, 엽상형, 다육질형, 각상형, 유절산호말형 순으로 출현비율이 높다고 보고되었다(Sohn, 1987; Park et al., 2007). 동해안에서 수행된 연구를 보면(Choi, 2008; Kwon and Choi, 2014), 성긴분기형, 사상형, 엽상형, 다육질형, 유절산호말형, 각상형이 순차적으로 높은 비율로 출현하여 다른 해역과 유사한 결과를 얻었다. 동해안에 위치한 왕돌초의 경우, 사상형(34.4%), 성긴분기형(25.4%), 엽상형(24.6%), 다육질형(9.7%), 각상형(5.7%), 유절산호말형(0.8%)의 비율을 보여 많은 연구에서 성긴분기형과 사상형의 구성 비율이 높게 나타난다는 연구 결과들과 유사한 패턴을 보였다. 이번 오륙도 주변 해역의 연구에서는 성긴분기형(30.3%), 사상형(24.2%), 다육질형(19.7%), 엽상형(15.2%), 유절산호말형(7.6%), 각상형(3.0%) 순으로 구성 비율을 보여, 이전 결과들과 큰 차이 없이 대부분 유사하게 출현하였고 결과적으로 구성 비율의 차이는 크지 않았다.
해조류의 기능형군을 근거로 생태학적 특성에 따라 다년생의 느린 생장을 나타내는 해조류인 다육질형, 유절산호말형, 각상형 해조류를 생태학적 상태그룹 I (ESG I), 교란이나 환경오염 등이 상대적으로 높아 비교적 빠른 생장 특성을 보이는 엽상형, 사상형, 성긴분기형의 해조류를 생태학적 상태그룹 II (ESG II)로 구분하였다(Orfanidis et al., 2001). 이번 연구에서 ESG I에 해당하는 해조류는 30.3%, ESG II에 속하는 해조류 비율은 69.7%로 나타나, ESG II에 해당하는 기회종 성격의 빠른 생장형 해조류 비율이 높았다. Shin et al. (2014)은 인위적인 활동에 의해 발생하는 오염물질 유입 지역이나, 환경스트레스에 따라 오염도가 높은 해역에서 ESG II에 속하는 해조류가 우세하다고 보고하였는데 오륙도의 경우에도 ESG II의 비율이 높아 연안 환경이 오염에 노출되었을 가능성이 높음을 판단할 수 있다.
이상의 결과로 보았을 때, 오륙도 인근 해역에 생육하는 해조류 식생은 인근 해역에서 수행된 용호동(Nam and Kim, 1999), 동백섬(Yoo, 2003a), 서암(Yoo, 2003b), 영도(Choi, 2007), 일광만(Kang et al., 2008), 기장(Yoo et al., 2013), 이기대(Shin et al., 2014) 등의 연구결과와 유사하게 나타났지만, 1970년대에 부산 동백섬에서 수행된 연구결과(Lee and Kang, 1971)에 비해서는 대체적으로 빈약한 해조상을 나타냈다. 생육 해조류의 감소 경향은 육지로부터 유입되는 오염물질 및 각종 공사로 인한 오염 부하량의 증가도 한 원인으로 연관성이 있는 것으로 판단된다(Nam and Kim, 1999). 해조류 출현 종 수, 종 다양성 및 생물량의 변화는 전국적으로 확산되는 갯녹음, 연안 개발 등과 함께 더욱 가속화 될 것으로 여겨진다(Yoo et al., 2013). 오륙도 및 주변해역은 해양생태계를 종합적이고 체계적으로 보전, 관리함으로써 국민 삶의 질을 높이고 해양자산을 보호하는 것을 목적으로 해양생태계 보호구역으로 지정이 되었다. 하지만, 해양보호구역 지정 자체가 정책의 목표였기 때문에 지금까지 보호구역 지정의 목적 달성을 위한 효과적인 정책적 관리수단은 미흡한 수준이었다. 오륙도는 기암괴석의 무인도서로, 수직암반의 생물상 보호가 필요하기 때문에 해양보호구역의 특성을 대변할 수 있는 몇 개의 정점과 보호구역 설정으로 인한 변화를 비교하기 위한 몇 개의 비교구역을 설정해서 연구 및 모니터링하는 것이 바람직하다고 할 수 있다. 따라서 오륙도 및 주변 해역의 해조 식생을 보전하기 위해서는 과학적인 관리가 될 수 있도록 주기적인 모니터링과 함께 생태공학적 해중림 조성 및 유지 관리 등 보다 적극적이고 체계적인 연구가 이루어져야 할 것으로 판단된다.