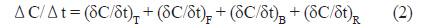우리나라 굴(
그러나 1970년대부터 환경오염, 적조, 빈산소, 과밀양식 등의 원인으로 대량폐사가 발생하여 굴 양식업과 양식 해역이 큰 피해를 입었으며(Cho, 1979; Bae and Han, 1998; NFRDI, 1998, 2009), 최근 들어 육상의 오염원 감소 등으로 폐사발생 횟수나 강도는 약화되었으나 여전히 지속되고 있는 실정이다. 양식장 자가오염, 해면의 지속 이용, 저질악화 등으로 유발되는 환경문제와 과밀양식에 대한 해결책을 제시하고자 굴 양식의 적정 수용력 산정연구가 2000년대에 수행되었다(Park et al., 2002; Oh et al., 2002; Jeong et al., 2009). 그 결과 지역에 따라 굴 양식에 적합한 환경유지를 위해 시설을 당시보다 30-40%를 감소해야 한다는 결과도 보고되었다(Park et al., 2002). 한편 굴 양식과 환경과의 상호작용을 정밀하게 파악하기 위한 연구도 시도되었다. 양식 해역에서 입자성 유기탄소의 일 변동을 관측하고 굴의 먹이로 유기탄소가 이용되는 효율을 파악하거나(Kang et al., 1993), 양식장 저질에서 유기물 분해와 영양염의 재용출율을 측정하여 환경에 미치는 영향을 파악하기도 하였다(Lee et al., 2011a). 이처럼 굴 양식은 우리나라 수산업과 수출 그리고 연안환경 측면에서도 중요한 산업이므로 1970년대부터 연구가 활발히 수행되고 있다(Lim et al., 1975; Choi et al., 1997; Lee et al., 2011b).
대기 온실기체 증가로 말미암은 기후변화현상은 해양의 수온상승뿐 아니라 해양산성화(ocean acidification)와 저산소화(deoxygenation)를 유발하여 세계적인 우려를 낳고 있다(IPCC 2007). 특히, 해수의 pH가 지속적으로 감소하는 해양산성화는 해양 및 수산생물의 생리활성, 성장, 생존율 등에 영향을 미치고, 특히 탄산염광물 형성을 저해하기 때문에 패류와 산호 등에 치명적인 영향을 미친다(Hoegh-Guldberg et al., 2007; Kroeker et al., 2013). 따라서 각종 패류와 산호를 대상으로 해양산성화 영향연구가 폭발적으로 수행되고 있으며, 대체적으로 굴의 성장, 생존, 패각 등에 영향을 끼친다는 결과가 우세하다(Miller et al., 2009; Parker et al., 2011; Barton et al., 2012). 연안의 경우, 기후변화에 따른 전지구적 해양산성화 영향과 더불어 육상 및 자가오염에 따른 부영양화로 인한 산성화도 있어왔으므로(Salisbury et al., 2008; Borges and Gypens, 2010) 대양보다 더욱 취약하다 할 수 있다. 따라서 해양산성화로 인해 연안의 굴 양식산업도 영향을 받을 수 있으며, 그에 따른 영향평가와 기초 조사가 필요하다. 우리나라 해양산성화 연구는 실내배양을 통해 해양생물의 생리/생존에 대한 영향평가 위주로 실시되었으며, 현장환경에서 산성화 진행 및 생물에 미치는 영향 등에 관한 연구는 많지 않다. 특히, 굴 양식해역에서 이산화탄소를 비롯한 무기탄소 분포 및 순환에 대한 연구는 거의 수행된 바 없다.
본 연구는 굴 양식이 성행하는 경상남도 고성만의 굴양식장에서 해양 환경 및 탄소계 인자를 연속 관측함으로서 고해상도의 환경변화를 파악하고, 특히 이산화탄소분압에 영향을 끼치는 인자들에 대해 정량적으로 평가하고자 한다. 이를 통해, 굴 양식장의 탄소순환 특성과 양식생물과 환경 사이의 상호작용을 파악하여, 양식생물의 폐사원인, 생육조건 등을 파악하는데 적극활용하고 수온상승, 해양산성화(ocean acidification) 등과 같은 미래 해양환경변화에 대비하고자 한다.
경상남도의 양식 굴(
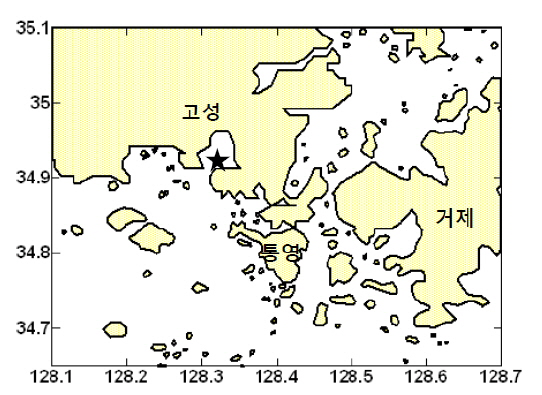
2011년 11월 2일~11월 4일에 고성만(경상남도 통영시 도산면 오륜리 앞)에 위치하는 굴 양식장(34°54.85′N, 128°19.2′E)에서 국립수산과학원 조사선 탐구10호를 타고 해양환경 및 탄소계 인자에 대한 연속 관측을 수행하였다(Fig. 1). 양식장은 육상에서 직선으로 2-3 km 거리에 위치하였으며, 수심은 6.5-7.8 m (약 7 m) 였다. 통영시 기상관측소에서 측정한 기온은 조사기간 중 11월 3일 오후 2시에 22.9℃로 가장 높았고 11월 4일 오전 7시에 16.5℃로 가장 낮았으며, 풍속은 11월 3일 오전과 4일 새벽에 3-4 m s-1로 높고 그 외 시간에는 2 m s-1 이하로 낮았다(http://www.kma.go.kr). 조사기간 통영지역의 조위변화는 105-191 cm이었으며, 만조는 11월 3일 2시, 15시, 4일 4시경이었으며, 간조는 11월 2일 20시, 3일 8시, 22시, 그리고 4일 9시경 이었다(http://www.khoa.go.kr). 조사가 진행된 굴 양식장은 막바지 출하를 위해 양성 중이었으며, 주변 양식장에서는 수확이 끝났거나 수확 중인 곳도 있었다.
양식장의 수질을 연속 관측하기 위한 시스템은 열염측정기(thermosalinograph) (SBE45, Sea-Bird Electronics, USA),
표층수의 수온과 염분을 연속 관측하는데 이용한 열염측정기 관측결과는 CTD (19 plus, Sea-Bird Electronics, USA)와 비교 검정하여 사용하였다. 해양
실시간 연속관측시스템의 결과를 보완하고 해수 수층 분포특성을 파악하기 위해 2시간 간격으로 CTD (19plus, SBE, USA)로 약 8 m 수심까지 수온과 염분을 관측하였으며, 다목적수질 측정기(YSI6000, YSI, USA)로 용존산소 농도와 포화도를 측정하였다. 또한 표층수의 영양염과 용존산소, 엽록소, 부유물질 분석을 위해 해수펌프로 공급되는 해수를 2시간 간격으로 채수하여 항목에 따라 전처리를 하여 추후 분석을 위해 냉장/냉동보관을 하였다. 용존산소는 현장에서 정밀적정기(Dosimat 876 system, Metrohm, Switzerland)를 사용하여 Winkler법으로 분석하였다(Parsons et al., 1984). 영양염은 현장에서 GF/F여과지로 여과 후 냉동 보관한 시료를 실험실에서 영양염자동분석기(QuAAtro system with 4 channel, BranLuebbe, Germany)로 분석하였으며, 표준물질(OSIL社 & Wako社)로 검정하였다. 엽록소와 부유물질도 해양환경공정시험기준(MLTM, 2010)에 의거하여 전처리와 분석을 하였다. 총알칼리도(total alkalinity, TA)는 개방형 용기법(KORDI, 2010)으로 정밀 적정하였으며, 미국 스크립스연구소의 A. Dickson 박사 실험실에서 제작한 표준물질 CRM Batch #99과 #113로 검정하였다. 용존무기탄소(Dissolved inorganic carbon, DIC)는 총알칼리도와 연속 관측한 이산화탄소분압으로 계산하였다(http://cdiac.ornl.gov/ftp/co2sys, Lewis and Wallance, 1998). 또한 해류 특성을 파악하기 위하여, 유속계(Anderaa, Norway)로 유속과 유향을 측정하였다.
굴 양식장 표층수에서 측정한 이산화탄소분압의 시간에 따른 변화를 평가하기 위해 다음의 단순 수지식(simple mass balance equation)을 적용하였다.

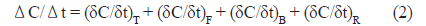
식(1)에서 Ct는 t시간일 때 이산화탄소 농도를 의미하며, ΔC는 t와 t+Δt사이 변화한 이산화탄소농도를 의미한다. 그리고 Δt는 표층수를 채수한 간격으로 2시간이며, 3일과 4일의 0시 전후에만 각각 6시간과 4시간이다. 식(2)에서 (δC/δt)T는 수온에 의한 이산화탄소변화율, (δC/δt)F 대기-해양간의 교환에 의한 변화율이며, (δC/δt)B는 생물생산에 의한 변화율이며, (δC/δt)R는 그 외 요인으로 발생하는 변화율을 총괄적으로 포함하는데 주로 물리혼합(조석, 이류, 수직혼합 등)을 의미한다.
대기-해양 이산화탄소 교환율은 Wanninkhof (1992)에 따라 계산하였다
FCO2 = k × s × Δ
여기서 k는 기체교환속도(cm h-1)이며 s는 수온과 염분에 따른 기체 용해도로 Weiss(1974)에 따라 계산하였으며, ΔpCO2는 대기-해양 이산화탄소분압 차(
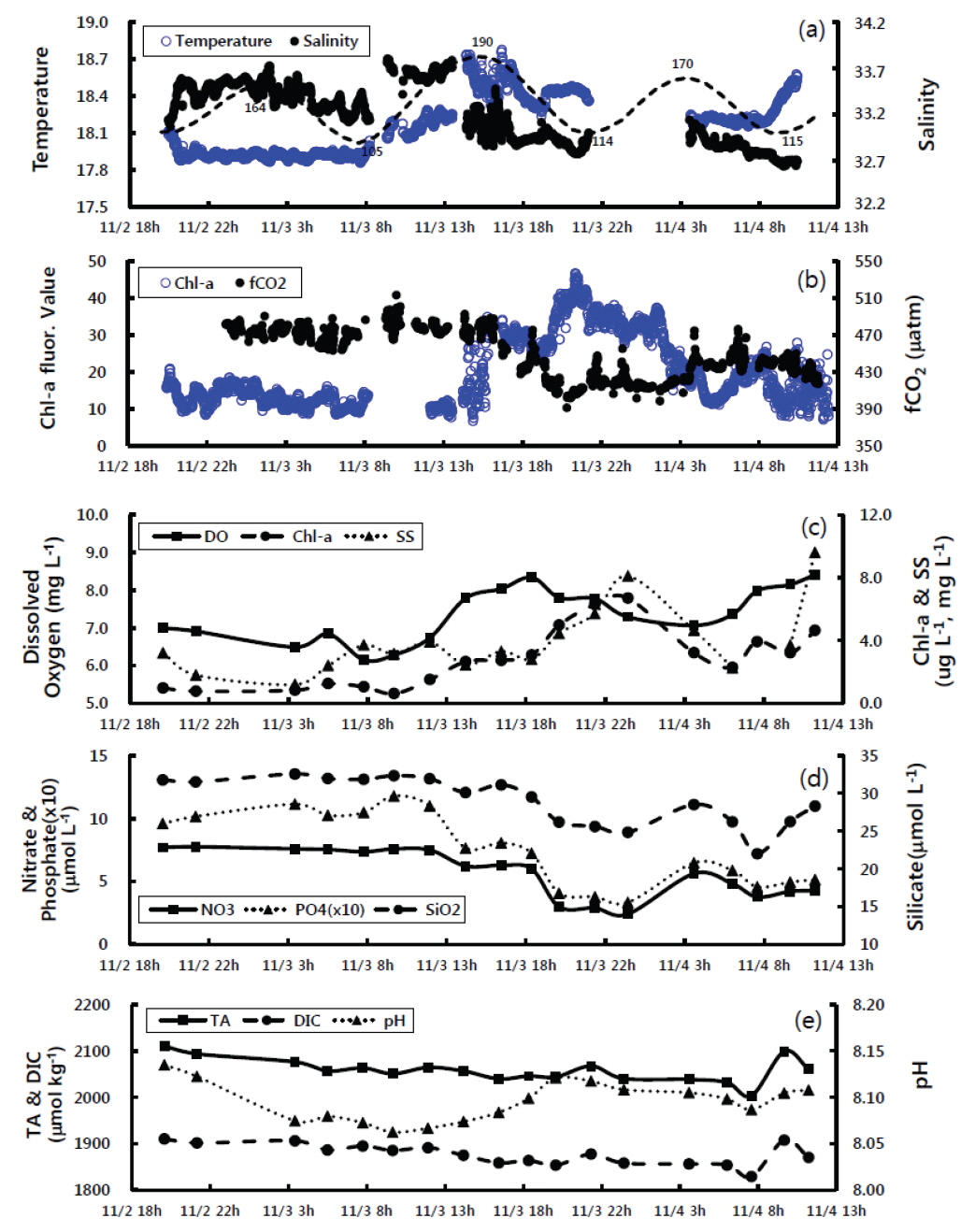
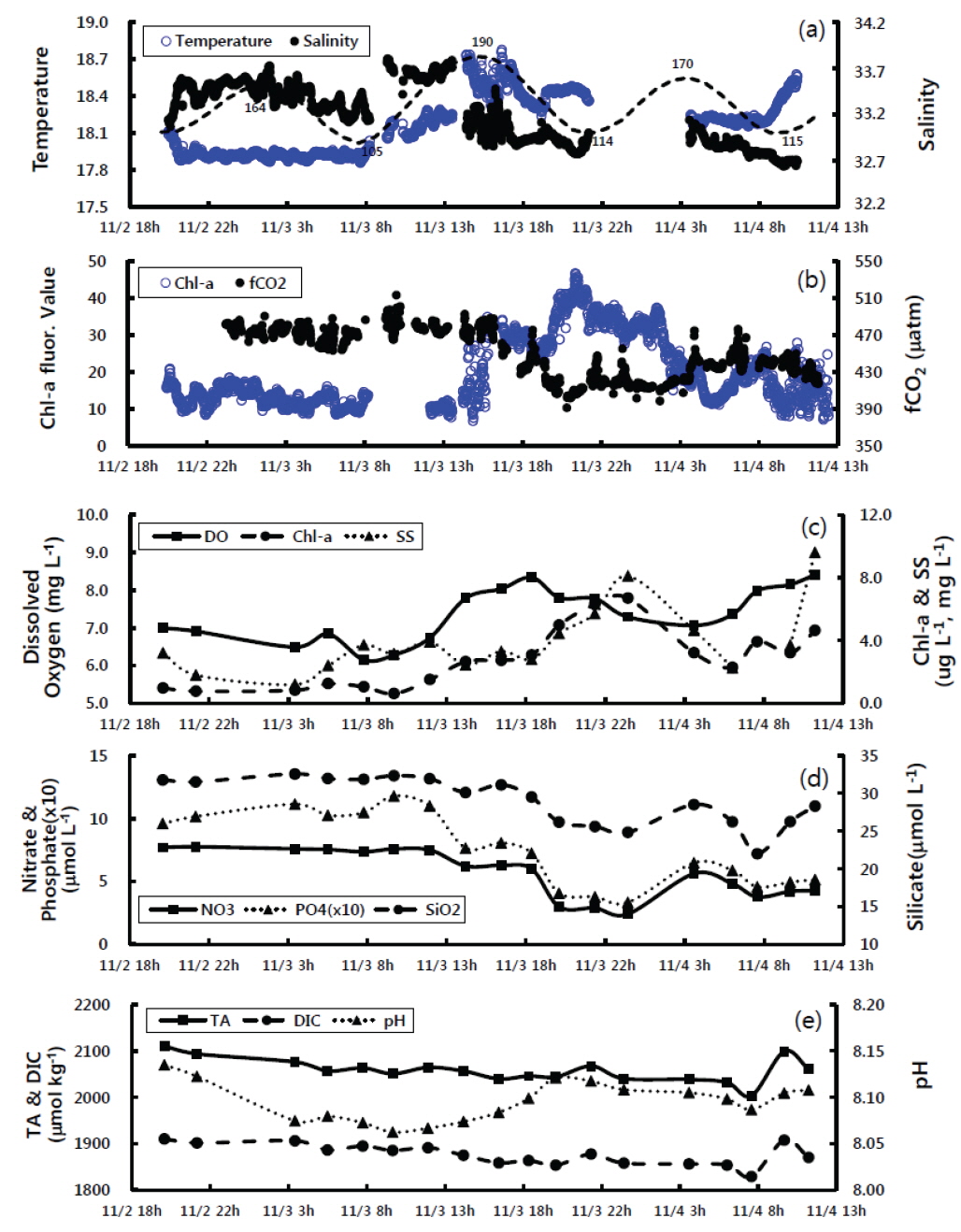
경상남도 통영시와 고성군에 둘러싸인 고성만의 굴양식장에서 2011년 11월 2일 ~ 4일 동안 실시간 연속 관측시스템으로 탄소계 인자를 포함한 해수특성을 관측하였다(Fig. 2). 굴 양식장의 표층수온은 조사기간 17.9-18.7℃ 범위를 보였으며, 11월 2일 20시부터 3일 8시 까지는 17.9-18.0℃ 사이로 비교적 낮았으며, 9시부터 점차 상승하여 15시에 최고 수온(18.7℃)을 보였다[Fig. 2(a)]. 이후 점차 감소하는 경향이었으나, 장비의 오류로 인해 11월 3일 22시부터 4일 4시까지 관측이 중단되었다. 관측이 재개된 11월 4일 4시부터 9시까지 표층수온은 약 18.2℃의 범위였다가 이후 증가하는 경향이었다. 한편 11월 4일 새벽(3-6시)은 수온이 18.2℃로, 11월 3일 비슷한 시간에 비해 약 0.3℃ 높았으며 8-9시부터 상승경향도 다소 빨랐다. 표층수의 염분은 32.65-33.75 psu의 매우 좁은 범위에서 변화하였다. 염분은 11월 2일부터 3일사이에는 조석영향으로 만조에는 높고 간조에 낮은 경향을 보였으나 3일 오후부터는 전반적으로 낮았다[Fig. 2(a)]. 조사기간 중 최대 만조였던 11월 3일 14시에 가장 높은 염분값을(33.7 psu) 보였으며, 이후에는 수온과 반대로 3일보다 낮게 나타났다. 이처럼 표층수의 수온과 염분 분포는 비슷한 시간대임에도 불구하고 다소 다른 양상을 보임으로서 일간(inter-daily)변화도 상당함을 알 수 있었다.
통영 굴 양식장 표층수의 이산화탄소분압은 390-510 μatm의 범위를 보여, 조사기간동안 대기 이산화탄소농도(398-420 μatm)보다 5-75 μatm 높았다[Fig. 2(b)]. 조사초기 이산화탄소 분압은 470±10 μatm 내외로 일정하였으나, 11월 3일 오후 6시부터 8시동안 감소하여 400 μatm 정도로 4일 3시까지 유지하다가 이후 소폭 증가하였다. 표층수의 형광특성은 조사초기 10-20범위에서 큰 변화가 없었으나, 11월 3일 13시경부터 지속적으로 상승하여 21시경 최고값을 보였으며, 이후 점차 감소하여 약 10까지 낮아졌다가 4일 아침부터 다시 상승하는 경향이었다[Fig. 2(b)]. 한편, 표층수를 2-4시간 간격으로 채수하여 측정한 엽록소-a는 조사초기 0.6-1.2 μg L-1범위의 낮은 값을 보이다가 11월 3일 오후 2시경부터 점차 증가하여 4일 0시경 최고값(6.7 μg L-1)을 보였으며 이후 약간 감소하다가 4시경부터 다시 상승하였다[Fig. 2(c)]. 엽록소-a의 시간에 따른 변화는 표층수를 연속관측한 형광특성의 시간에 따른 경향과 매우 일치하였다(R2=0.65). 따라서, 표층수의 형광특성은 수중의 엽록소-a에 대한 간접지시를 한다고 할 수 있었다. 용존산소는 6.2-8.4 mg L-1의 범위를 보였으며, 조사초기에는 7.0 mg L-1의 낮은 값을 보였으나, 3일 오후 2시부터 점차 증가하여 오후 6시경 가장 높았다[Fig. 2(c)]. 이후 밤시간동안 다소 낮아졌다가 11월 4일 4시경부터 다시 증가하는 경향으로, 엽록소-a의 변화경향과 매우 유사하였다. 부유물질은 1.2-9.6 mg L-1의 범위를 보였으며, 시간에 따른 변화는 엽록소-a와 매우 유사하게 조사초기에는 비교적 낮은 농도를 유지하다가 3일 오후부터 상승하여 밤에 최고치를 보였다[Fig. 2(c)]. 표층수의 영양염분포 경향을 Fig. 2(d)에 나타내었다. 질산염과 인산염 그리고 규산염은 조사기간 각각 2.4-7.8 μmol L-1, 0.3-1.2 μmol L-1, 22.0-32.6 μmol L-1의 범위를 보였다. 질산염을 포함한 영양염은 조사초기 비교적 높은 농도를 지속되다가 11월 3일 오후부터 점차 감소하여, 24시경 가장 낮은 농도를 나타내었으며, 이후 상승하다가 4일 아침에 소폭 감소하는 경향으로, 엽록소-a와 상반되는 변화를 보였다. 표층수의 총알칼리도는 2,000-2,110 μmol kg-1의 범위로, 조사초기 높은 농도를 보였다가 시간이 경과할수록 점차 감소하였으며, 4일 8~10시 사이에 소폭 증가하였다[Fig. 2(e)]. 용존무기탄소 농도 범위는 1,830-1,910 μmol kg-1였으며, 총알칼리도와 매우 유사한 변동경향을 보였다[Fig. 2(e)]. 표층수의 pH는 8.07-8.14의 범위를 보였으며, 조사초기 가장 높은 값에서 11월 3일 8시경까지 점차적으로 감소하다가 낮부터 20시까지는 증가하였다[Fig. 2(e)]. 이후 전반적으로 일정한 pH를 유지하였으며 4일 오전에는 총알칼리도와 마찬가지로 8시경 낮았다가 다시 증가하였다.
2011년 11월에 측정한 고성만 굴 양식장의 표층수의 환경성분은 조석과 일기변화에 따라 염분과 영양염, 용존무기탄소, 이산화탄소분압은 대체로 조사초기에는 높았다가 후반으로 갈수록 낮아지는 경향이었으며, 수온과 용존산소, 엽록소-a, pH는 조사초기에 낮았다가 후반으로 갈수록 높아지는 경향이었다.
>
굴 양식장 표층수의 일반성분과 탄소계인자 사이의 상관성
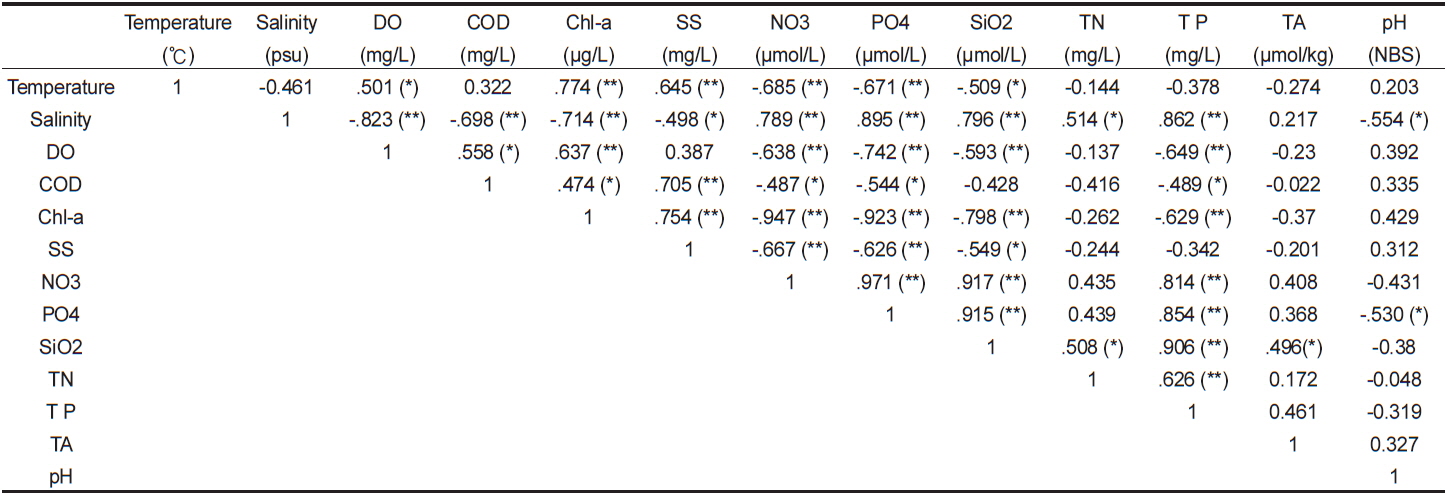
고성만 굴양식장에서 관측한 표층수의 물리화학특성 결과를 바탕으로 이들 성분간에 통계적인 상관성을 파악하였다(Table 1). 표층수의 수온과 염분사이에는 유의하진 않으나 음의 관계를 보였으며 이는 대체적으로 수온이 높을 때 염분이 낮다는 것이다. 그러나 관측시기 중 최대 만조(11월 3일 14시)전후에 수온과 염분 모두 최대값을 나타내었는데, 이는 염분의 경우 조석의 영향으로 최대값을 보인 반면 수온은 조석과 더불어 태양복사열의 영향으로 상승하였기 때문이다. 또한 표층 수온은 엽록소-a와 용존산소, 부유물질과 양의 유의한 상관성을 보이고 영양염류와는 음의 유의성을 보였다. 이는 수온이 높은 낮에 엽록소-a농도가 높아지고 이에 의한 일차생산결과, 영양염은 소모되고 용존산소는 생산되는 것으로 유추할 수 있다. 따라서 굴 양식장 표층수에서 엽록소-a가 증가할수록 용존산소도 유의하게 증가하였으며(P<0.01) 더불어 영양염류도 유의하게 감소하였다(P<0.01). 통영주변해역의 수질 월변화에서도 수온이 높고 엽록소-a 농도가 높은 시기에 표층수의 영양염이 낮았으며, 특히 양식해역은 수온이 낮은 시기에도 그런 경향이 지속된다고 하였다(Lee et al., 2011b).
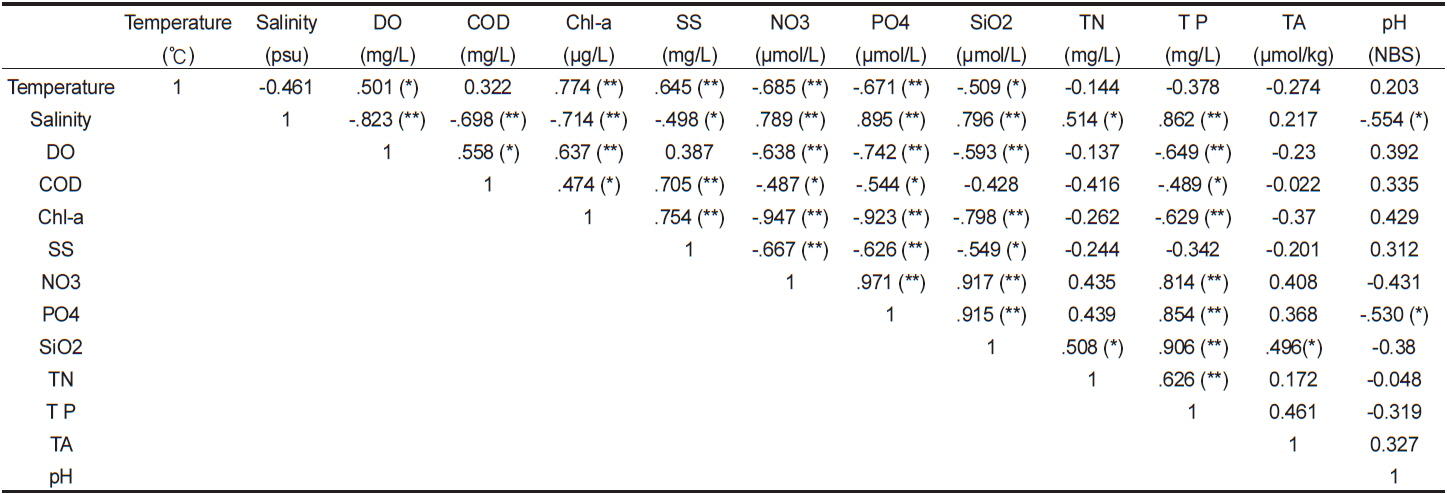
Correlations between environmental parameters measured at an oyster Crassostrea gigas farm in Goseong Bay in Nov. 2011 (*, P<0.05; **, P<0.001)
총질소(total nitrogen, TN)와 총인(total phosphorus, TP)도 양의 유의한 상관성을 나타내었다(
이상에서 굴 양식장의 표층수는 수온, 염분과 같은 물리특성과 엽록소-a와 영양염류 등과 같은 생물화학특성이 서로 유의한 상관성을 보였으며, 이는 물리환경 변화에 생물생리활동이 민감하게 반응하는 것을 의미한다. 따라서 굴 양식장 환경인자들 간에 물리-생물-화학적 고리가 강하게 연결된 건강한(풍요로운)복합 생태계임을 파악할 수 있었다. 1970년대 중반에 실시된 조사에서도 고성만은 저염, 고영양염, 풍부한 식물플랑크톤 등의 환경특성을 보였으며, 이로 인해 주변 해역(한실포, 사량도, 자란만 등)에 비해 굴 성장이 좋고 폐사비율이 낮은 해역으로 평가되었다(Lim et al., 1975).
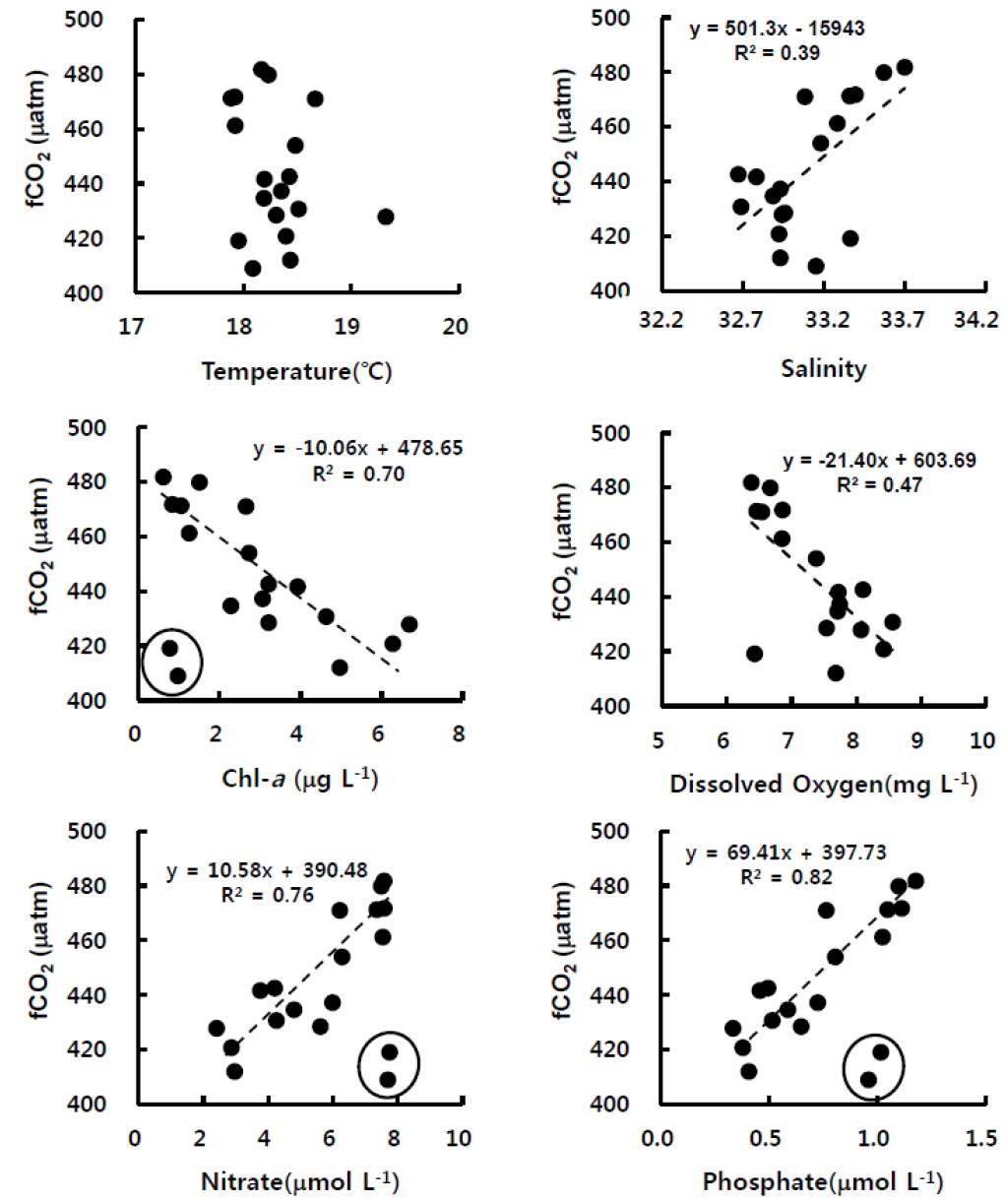
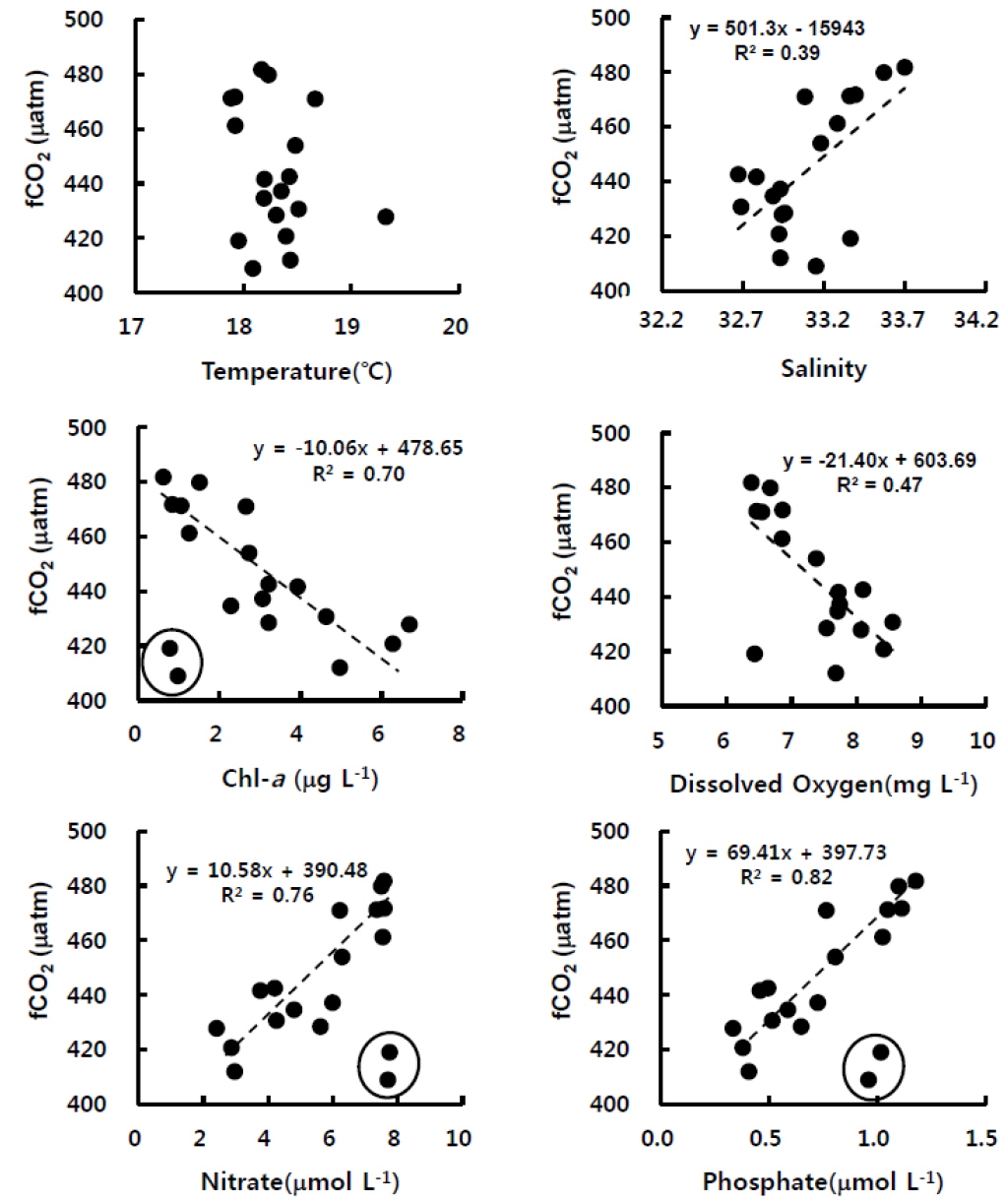
표층수의 이산화탄소분압은 해수 및 대기의 혼합과 교환 그리고 생물생산과 호흡에도 밀접하게 관련된 인자이다. 굴 양식장에서 연속관측한 이산화탄소 분압은 엽록소-a (R2=0.70), 질산염(R2=0.76), 인산염(R2=0.82)과 매우 높은 관련성을 각각 나타내었다(Fig. 3). 이는 앞서 언급하였지만, 엽록소-a로 대변되는 식물플랑크톤류가 수층의 이산화탄소와 영양염류를 이용하여 활발하게 광합성을 하면 그 결과 용존산소가 생성되는 것이다. 일반적으로 수온은 탄소계인자간의 구성비를 변화시킴으로 인해 이산화탄소분압 농도를 변화시키는 주요 요인으로 알려져 있으나, 상관성이 거의 없는 것으로 나타났다. 이는 조사기간의 수온 변화 자체가 크지 않았으며(1℃ 내외), 수온이 높았던 시간에 엽록소-a도 높아서 이산화탄소분압 변화에 서로 상쇄하는 역할을 하였던 것으로 생각된다. 그러나 표층수의 이산화탄소분압과 염분은 양의 선형관계를 나타내었는데, 이는 염분이 높을 때 염(salt)을 구성하는 용존무기탄소류의 농도가 상대적으로 높았기 때문이다. 이처럼 2011년 11월 고성만 굴양식장 표층수의 이산화탄소분압은 물리특성뿐 아니라 생물화학적특성 변화에 민감하게 변하는 것을 알 수 있었으며, 다음절에서 각 영향인자에 대해 정량적으로 파악하고자 한다.
통영 굴 양식장 표층수에서 관측한 이산화탄소분압은 390-510 μatm범위에서 증가와 감소를 반복하였으며, 엽록소-a, 영양염류, 용존산소와 상관성이 있는 것으로 나타났다(Fig. 3). 실제 해양에서 표층수의 이산화탄소분압 변화에는 수온, 대기-해양 교환, 생물생산(biological pump), 물리적 혼합 등의 요인이 복합적으로 영향을 미치는 것으로 알려져있다(Bakker et al., 1997; Louanchi et al., 1996; Shim et al., 2006; Shim et al., 2012). 따라서 굴 양식장에서 이산화탄소 분포에 영향을 끼치는 인자들에 대해 정밀하게 평가하고 각 항목의 상대 중요도를 시범적으로 파악하고자 단순 수지식 (1)과 (2)를 적용하였다.
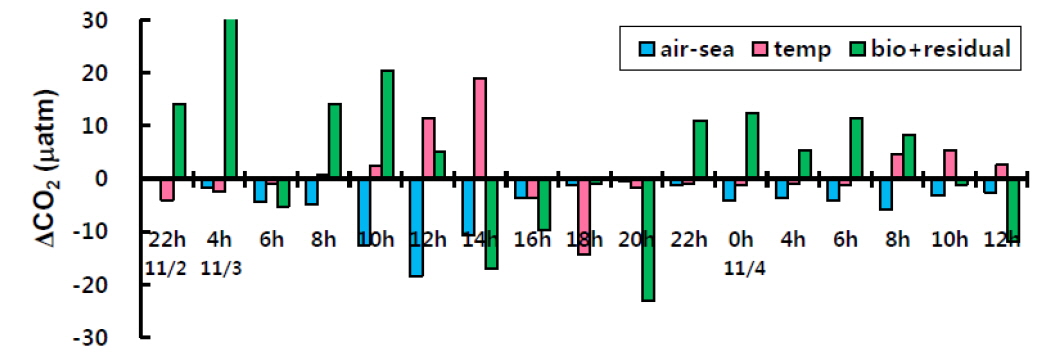
식(2)에서 Δt 동안 변화한 이산화탄소분압(ΔC)은 연속관측 결과에서 추출하여 파악할 수 있었으며, 약 -25~53 μatm 범위로 시간에 따라 증가와 감소를 반복하였다. ΔC은 Δt가 가장 길었던 2일 밤부터 3일 새벽 사이에 가장 크게 증가하였으며, 3일 낮부터 초저녁(10시~20시)동안에 가장 크게 감소하였다.
수온은 이산화탄소분압에 매우 강력한 영향을 미치는 열역학(thermodynamic) 인자로서, 수온이 1℃ 상승할 때 이산화탄소분압은 약 4.23%증가한다(Takahashi et al., 1993). 따라서 (δC/δt)T는 t와 t+Δt 사이에 상승/하강한 수온변화를 연속관측 결과에서 추출하여 계산할 수 있다. 조사기간 동안 수온변화에 의한 이산화탄소분압 변화양[(δC/δt)T]은 -14.5~18.9 μatm으로 수온의 일변화에 따라 오전 6시부터 오후 2시 까지는 양의 값을, 오후 2시부터 다음날 오전 6시까지는 음의 값을 보였다(Fig. 5). 따라서 수온은 낮에는 이산화탄소분압을 상승시키는 역할을 하였으며 수온이 낮아지는 밤에는 이산화탄소분압을 감소시키는 역할을 하였다. 특히 조사기간 최대 고조기(190 cm)였던 3일 14시 전후에 (δC/δt)T가 양과 음의 최대값을 보였다. 이는 고조기일수록 빨라진 조석류로 인해 수온을 비롯한 환경이 급변하였고 이로 인해 이산화탄소분압 또한 변동이 컸던 것으로 생각된다. 한편, 본 연구의 굴 양식장보다 수온변화가 컸던(약 5℃) 해조류 양식장의 (δC/δt)T(-11~12 μatm, Shim et al., 2012)가 더 낮았는데, 이는 굴 양식장 이산화탄소분압 농도가 해조류 양식장보다 절대값이 높았기 때문이다.
2011년 11월 조사시기동안 고성만의 대기 이산화탄소농도는 평균 405 μatm으로 우리나라 11월 대기 이산화탄소 평균 농도(2012년 11월 평균 403 ppm, KMA, 2013)와 매우 유사하였다. 고성만 대기 이산화탄소 농도가 해수 이산화탄소보다 낮아서 조사기간 동안 해양에서 대기로 이산화탄소가 지속적으로 방출되었으며 그 교환율(flux)은 0.01-2.4 mmol m-2 d-1 이었다. 대기-해양 이산화탄소 교환에 의한 해수의 이산화탄소분압 변화[(δC/δt)F]는 0.1-18.5 μatm (평균 약 5.0 μatm)이었으며, 풍속이 2-3 m sec-1으로 높았던 3일 오전에 10 μatm 이상의 높은 값을 보였(Fig. 5).
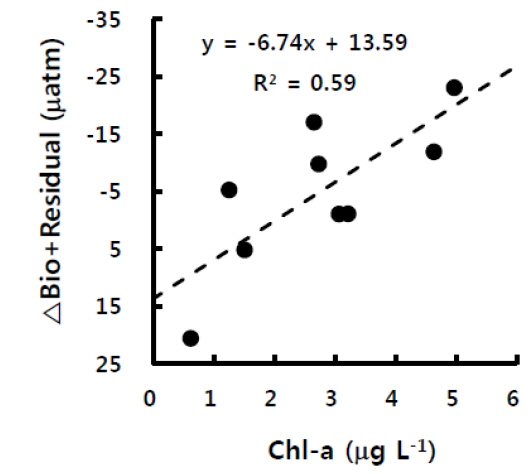
이상에서 이산화탄소분압의 총농도(ΔC)와 수온에 의한 변화[(δC/δt)T], 대기-해양교환에 의한 변화[(δC/δt)F]를 평가하였으며, 식 (2)에 대입하여 그 차이를 계산하면 생물에 의한 변화와 물리혼합에 의한 변화를 합한 값[(δC/δt)B+R]이 된다. 그 값은 -23.1~56.8 μatm의 범위를 보였으며(Fig. 5), 대체적으로 정오부터 늦은 오후 동안에는 음의 값을 보였으며, 밤부터 이른 아침까지는 양의 값을 보여, 생물생산에 의한 일변화와 유사한 경향이었다.
실제 굴 양식장에서 생물에 의한 이산화탄소분압 변화는 일반적인 해양환경보다 고려해야 할 대상이 더 많고 복잡하다. 식물플랑크톤을 비롯한 미소생물들에 의한 일차생산과 호흡도 있지만 양식 생물인 굴과 이에 부착된 생물에 의한 호흡, 그리고 굴의 패각(탄산염광물 CaCO3) 생성에 따른 이산화탄소 배출등이 있다. 따라서 생물에 의한 이산화탄소변화를 정밀하게 파악하기 위해서는 이들 각 항목에 대해 실내배양 또는 메조코즘(mesocosm) 등으로 정량화해야하나 본 연구에서는 현장 연속 관측을 통한 탄소수지 평가가 목적이므로 직접 측정하지 않았다.
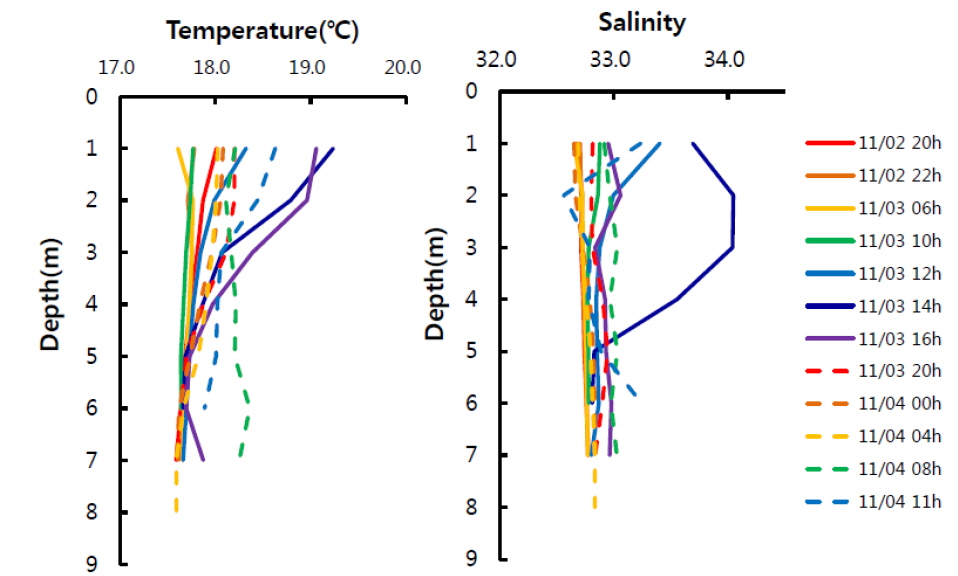
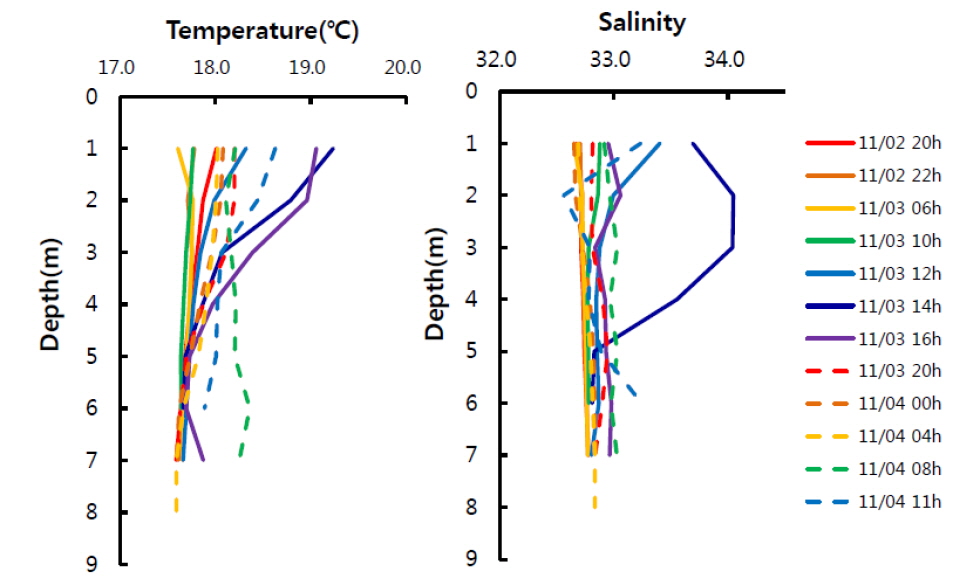
한편 고성만은 입구가 좁으며 수심이 얕은(평균 7 m) 전형적인 반폐쇄성 만이며, 만 내에 작은 섬들이 위치하고 있다. 따라서 고성만의 물리혼합을 주도하는 해류는 조석류이며, Oh et al. (2002)에 의하면 대조기의 최강류 시 만 입구에서 40-50 cm s-1, 북쪽 내 만은 약 10 cm s-1, 동쪽 내 만은 약 5 cm s-1 이하의 느린 유속분포를 보인다고 하였다. 특히 조사해역은 고성만의 동남쪽에 위치하여 유속이 가장 느린 해역이며, 조사기간 유속계로 측정한 표층과 저층의 유속도 0.3-3.0 cm s-1 범위로 낮았었다. 또한 얕은 수심으로 인해 표∙저층간의 환경이 균일하여 수직혼합에 의한 영향은 무시할 정도로 생각된다. 실제 수층의 수온과 염분분포에서도 최대 만조였던 11월 3일 14시 전후를 제외하면 표층과 저층의 수온과 염분 차이가 각각 1℃과 0.2이하로 아주 낮아 수직적으로 균일한 분포를 보였다(Fig. 4). 따라서 조사지역은 조석류의 흐름이 약하고 얕은 수심으로 표저층이 균일하여 전반적으로 물리혼합이 약한 지역이라 할 수 있다. 이러한 이유로 물리혼합에 의해 해양환경 변화가 크지 않으므로 이에 따른 이산화탄소분압 변화도 적으므로, 앞서 계산한 [(δC/δt)B+R]은 주로 생물에 의한 변화라고 할 수 있다. 한편, 수심이 약 15 m이며 표층과 저층의 수온 차가 컸던(3-5℃) 해조류양식장의 경우도 물리혼합에 의한 이산화탄소 증가는 약 10 μatm 이하(평균 4 μatm)로 낮은 것을 고려하면(Shim et al., 2012), 앞선 가정이 무리하지 않은 것으로 생각된다.
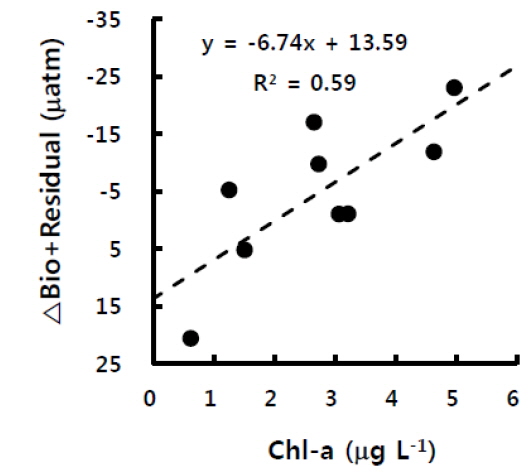
앞서 언급한대로 굴 양식장의 생물영향은 식물플랑크톤에 의한 광합성을 제외하면 대부분이 이산화탄소분압을 상승시키는 활동이므로, 광합성에 의해 상쇄가 되지 않으면 양의 값이 될 것이다. 실제 (δC/δt)B+R는 광합성 활동이 일어나지 않는 밤에 주로 양의 값을 보였으며, 특히 Δt가 가장 길었던 2일 밤부터 3일 새벽에 가장 높은 값을 보였으며, 이는 그 시간 동안의 생물호흡 결과가 축적된 것이다. 또한 낮 동안에는 엽록소-a 농도가 높을수록 (δC/δt)B+R의 감소가 더 컸는데(Fig. 6) 이는 (δC/δt)B+R가 물리혼합보다는 생물에 의한 변화량임을 뒷받침한다고 할 수 있다. 결국, 고성만 굴 양식장에서 이산화탄소분압에 미치는 영향인자 중 생물에 의한 변화를 간접 평가한 결과, 양식생물과 그 외 생물들에 의한 호흡으로 밤에는 이산화탄소가 증가하지만 낮에는 식물플랑크톤에 의한 광합성이 이를 상쇄하는 것으로 나타났다. 실제 고성만의 10월과 11월 일차생산력 평균은 약 0.15-0.17 gC m-2 day-1으로 다른 달에 비해 낮으나, 조사지역이 위치한 만의 남쪽은 북쪽보다 일차생산력이 높다고 하였으며, 양식 굴의 과밀정도를 평가하였을 때 11월은 0.6으로 오히려 적정 수용량보다 낮은 밀도로 양성중인 것으로 평가되었다(Lee, 2009).
Shim et al. (2012)은 해조류양식장의 경우 생물에 의한 이산화탄소 흡수[δC/δt)B]는 하루 중 5-21시 사이에 7-28 μatm범위(평균 12 μatm)라 하였다. 본 연구와 달리 양식생물이 해조류이고 호흡을 고려하지 않았기 때문에 밤에 호흡에 의한 이산화탄소 증가가 없었다. 한편, 양식생물, 양식환경, 조사시기 등이 다름에도 불구하고, 두 양식장의 낮 동안 생물에 의한 이산화탄소 변화[(δC/δt)B], 즉 생물펌프(biological pump)는 상당히 비슷한 범위에 속하였다.
이상에서 2011년 11월 고성만 굴 양식장의 이산화탄소분압변화에 미치는 요인에 대해 정량적 평가를 하였으며, 수온변화와 생물생산은 오후부터 초저녁에 이산화탄소분압을 감소시키는 요인이었으며, 초저녁부터 새벽에는 생물호흡이 주된 상승요인으로 작용하였다. 대기-해양교환에 의한 변화는 풍속이 2-3 m s-1 이상일 때 주요 요인으로 작용하였다.
한편 굴 양식장에서 굴의 육질부와 패각 성장은 궁극적으로 해양의 유·무기탄소를 제거하는 역할을 한다(Tang et al., 2011). 본 연구를 바탕으로 굴 양식장의 유기탄소순환과 양식생물의 섭식과 호흡에서의 탄소교환 등을 추가하여 굴 양식을 통한 탄소제거 및 연안 탄소순환에 기여하는 바를 평가할 필요가 있다.