염증의 발병은 외부 자극에 대한 인체의 정상적인 방어 시스템이지만(De Heredia et al., 2012; Osborn and Olefsky, 2012) 염증에 대한 지속적이고 과도한 면역반응은 오히려 조직의 손상을 촉진하고 만성 염증을 유발하게 된다(Brown et al., 2007). 이것이 지속화 되면 관절염, 당뇨병, 동맥경화, 암, 노화 및 알츠하이머병을 포함하는 퇴행성 신경 질환의 원인이 된다(De Heredia et al., 2012; Osborn and Olefsky, 2012) .
선천성 면역과 후천성 면역에 모두 관여하는 대식세포의 주작용은 reaction oxygen species (ROS)와 활성질소(reactive nitrogen species, RNS)를 무기로 사용하여 세균, 바이러스, 암세포에 대해 공격하며 T cell 활성화 및 B cell 활성화에 참여함으로써 면역계에서 총체적이고 핵심적인 역할을 하는 세포이다(Hakim et al., 2004). 또한 대식세포는 염증 촉진성 cytokine인 tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), IL-1β 등과 병원균의 내독소로서 여러 염증세포들이 생산하는 cytokine들의 생산을 촉진하는 lipopolysaccharide (LPS)에 의한 자극으로 활성화된다(Marriot et al., 1998). 활성화된 대식세포는 inducible NOS (iNOS), cyclooxygenase-2 (COX-2) 같은 효소를 생산하여 nitric oxide (NO) 및 prostaglandins E2(PGE2)같은 다양한 염증 매개체들을 생성한다(Nathan., 1992). 염증성 cytokine 및 iNOS, COX-2의 발현은 전사인자인 nuclear factor-kappa B (NF-κB)에 의해 조절되는데 NF-κB 또한 산화적 스트레스, LPS, cytokine등의 외부 자극에 활성화되어 핵으로 이동하여 면역 및 염증 반응에 관여하는 매개체들의 발현에 관여한다(Choi et al., 2003). NF-κB는 p65, p50, Inhibitor kappa B (IκB) subunit의 trimer로 구성되어 있고 산화적 스트레스에 의해 IκB가 분해되면 p65/p50 heterodimer가 핵 속으로 이동하여 DNA 결합을 하는 것으로 알려져 있다(Munoz et al., 1991). 이러한 과정에서 IκB를 분해시키는 IκB kinase의 활성화는 extracellular signal-regulated kinase (ERK), c-jun Nterminal kinase (JNK), p38, Akt 등의 kinase에 의하여 조절된다(Nair et al., 2004).
생체는 산소호흡으로 인한 대사과정에서 산화적 스트레스를 유발하는 물질을 생성하게 되는데 이것의 방어기전으로 우리 몸은 superoxide dismutase (SOD), catalase 등의 항산화효소를 생성하여 활성산소종으로부터 생체 기관을 방어한다고 알려져 있으며 활성산소종과 염증성 cytokine은 이러한 산화적 스트레스를 유발하는 기작과 관련이 있다(Cho et al, 2008).
홍조류에 속하는 김은 예로부터 한국을 포함한 중국, 일본 등 아시아 지역에서 섭취하여 왔으며 영양학적 측면으로 열량은 낮지만 비타민, 무기질, 식이섬유가 풍부한 특징을 가지고 있다. 또한 김은 폴리페놀을 함유하여 항산화활성을 가지는 것으로 확인되었으며(Lee and Oh, 2002) 김에 함유된 betaine은 혈중콜레스테롤을 저하시키는 효과가 있다고 보고되었다(Jung et al., 2001) 특히 건조 김의 10% 내외로 함유되어있는 수용성 다당인 포피란은 항종양 활성(Noda and Arashima, 1989)과 항산화 활성(Zhang et al., 2004), 지질대사 개선에 대한 효과(Lee et al., 2010) 등이 확인되어 있으나 김의 항염증 효과와 그에 대한 분자기전에 대한 연구는 미비한 실정이다. 따라서 본 연구에서는 염증반응으로 변화되는 세포 내부의 지표물질과 신호단백질을 측정하였으며, 김 추출물이 이에 어떠한 영향을 미치는지를 확인하였다.
본 실험에 사용한 방사무늬 김(
실험에 사용된 RAW 264.7 세포는 American Type Culture Collection (Manassas, VA, USA)에서 구입하여 사용하였다. 세포는 10% fetal bovine serum (Gibco BRL, Gaitherberg, MD, USA)과 5% penicillin/streptomyocin (Gibco BRL, Grand Island, NY, USA)이 함유된 Dulbecco's modified Eagle's medium (DMEM)배지에 37℃, 5% CO2가 유지되는 incubator에서 배양하였다. Cell culture plate에 RAW 264.7 세포가 70-80% 정도 자라면 PBS로 씻어낸 후 계대배양하고 배지는 2-3일 마다 교체하였다.
PYE의 처리에 따른 RAW 264.7 세포의 증식에 미치는 영향을 알아보기 위해 96-well plate에 96-well plate에 10×104 cells/well에 세포를 동일하게 분주한 후 세포부착을 위하여 24시간 배양하고, phenol red free-DMEM 배지에 12시간을 더 배양한 후 PYE를 0, 5, 10, 20, 40 µg/mL씩 농도별로 처리한 후 24시간 배양하였다. 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulphenyl)-2H-tetrazolium, inner salt/phenazine methosulfate. (MTS/PMS) solution (Promega Co., Madison, WI, USA)을 첨가하고 37℃에서 30분 반응시킨 후 ELISA plate reader (Benchmark microplate reader; Bio-Rad laboratories, Hercules, CA, USA)로 490 nm에서 흡광도를 측정하였다.
RAW 264.7 세포를 100-mm dish에 배양하고 세포가 60-80%정도 증식하면 serum free media (SFM)으로 교체하였다. 6시간 뒤에 PYE (10, 20 µg/mL)가 처리된 SFM으로 교체하고 4시간 후에 1 µg/mL LPS를 20시간 동안 처리한 후 phosphate buffered saline (PBS)용액으로 세척하고 1 mL Lysis buffer [1% Igepal CA-360, 20 mM Tris- HCl (pH 8.0), 137 mM NaCl, 10% glycerol, 2 mM EDTA, 10 µg/mL aprotinin, 10 µg/mL leupeptin, 10 µg/mL pepstatin]로 세포를 취한 다음 12,000
단백질 발현의 분석은 RAW 264.7 세포 세포를 100-mm dish에 배양하고 세포가 60-80%정도 증식하면 SFM으로 교체하였다. 6시간 뒤에 PYE (10, 20 µg/mL)가 처리된 SFM으로 교체하고 4시간 후에 1 µg/mL LPS를 20시간 동안 처리한 후 PBS용액으로 세척하고 Lysis buffer [50 mM Tris-hydrochloride(Tris-HCl), pH 7.4, 150 mM sodium chloride (NaCl), 1mM ethylenediaminetetraacetic acid (EDTA), 1 mM sodium fluoride (NaF), 1% NP-40, 1 mM sodium orthovanadate(Na3VO4), 1 µg/mL aprotinin, 1 µg/mL leupeptin, 1 μg/mL pepstatin, 1 mM phenylsufonyl fluoride (PMSF), 0.25% Nadeoxycholate]를 radioimmunoprecipitation assay buffer [1% NP-40, 0.25% sodium deoxycholate, 1 mM ethylene glycol tetraacetic acid (EGTA), 150 mM NaCl, 50 mM Tris-HCl, pH 7.5]에 넣어 cell lysate를 회수하고 30분간 방치한 후 원심분리(12,000 rpm, 10 min, 4℃) 하여 단백질 농도를 동일하게 정량하여 SDS-PAGE에 전기영동한 다음 PVDF membrane (Millipore Co., Billerica, MA, USA)으로 이동시켰다. 이때 표준 분자량은 dual color marker를 사용하였다. 전기영동 시킨 membrane은 실온에서 1% bovine serum albumin/Tris-buffered saline-Tween 20 로 1시간 30분 동안 blocking 시킨 후 각각의 1차 antibody를 1:1,000으로 희석하여 4℃에서 16시간 반응시키고 2차 antibody를 1:10,000 비율로 희석하여 1시간 30분간 반응시킨 후 Super Signal West Pico Stable Peroxide Solution과 Super Signal West Pico Luminol/ Enhancer solution(Thermo Fisher Scientific Inc., Rockford, IL, USA)을 사용하여 KODAK X-ray film에 감광시켜 현상하여 시각화하였다.
RAW 264.7 세포를 5×104 cells/well 농도로 96-well plate에 배양한 후에 PYE를 0, 5, 10, 20, 40 µg/mL로 처리하고 4시간 반응시킨 후 LPS 1 µg/mL를 처리하여 24시간 배양한 뒤 세포 배양액 100 µL와 griess reagent (1% Sulfanilamide, 0.1% Naphthlethy-diamine-dihydro-chloride in 2.5% phosphoric acid) 100 µL를 혼합하여 실온에서 10분 동안 반응시킨 후 570 nm에서 ELISA plate reader (Benchmark microplate reader; Bio-Rad laboratories, Hercules, CA, USA)로 흡광도를 측정하였다.
세포내 활성 산소(ROS)를 측정하기 위하여 활성산소와 반응하면 형광을 발산하는 DCF-DA (2′7′-dichloro-fluorescein diacetate) (Sigma, St. Louis, MO, USA)을 이용하여 측정하였다. RAW 264.7 세포를 5×104 cells/well 농도로 96-well plate에 배양한 후에 PYE를 0, 5, 10, 20, 40 µg/mL로 4시간 처리한 다음 LPS 1 µg/mL농도로 처리하여 24시간 배양시킨 후 배지를 모두 제거하고 PBS에 DCF-DA 첨가, 37℃, 30분 반응한 뒤 excitation wavelength 485 nm, emission wavelength 530 nm에서 fluorescence microplate reader를 이용하여 활성산소를 확인하였다.
세포의 SOD측정은 superoxide dismutase assay kit (Cayman chemical, Ann arbor, MI, USA)를 이용하여 측정하였다. PYE와 LPS가 처리된 RAW 264.7 세포를 extraction buffer (20 mM HEPES buffer, pH 7.2, 1 mM EGTA, 210 mM mannitol, 70 mM sucrose)를 이용하여 회수하여 원심분리(1,500
Catalase의 측정은 catalase assay kit (Cayman chemical, Ann arbor, MI, USA)를 사용하여 측정하였다. PYE와 LPS가 처리된 RAW 264.7 세포를 extraction buffer (50 mM potassium phosphate, pH 7.0, 1 mM EDTA)를 이용하여 회수한 뒤 원심분리 (1,500
모든 실험의 분석 결과는 각각의 군별로 평균과 표준편차(mean±S.D.)로 나타내었으며 실험군 간의 유의성은 SPSS 프로그램(Statistical Package for Social Science, SPSS Inc. Chicago, IL, USA)을 이용하여 나타내었다. 반복 측정에 의한 ANOVA Test로 검증한 후, Duncan's multiple range test를 통하여
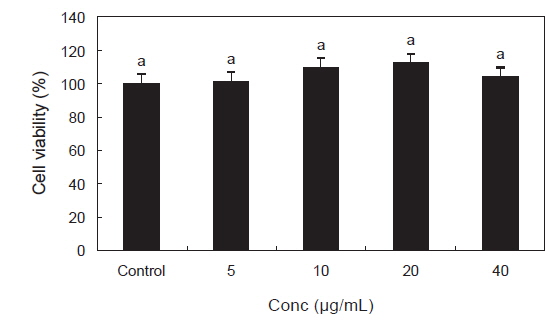
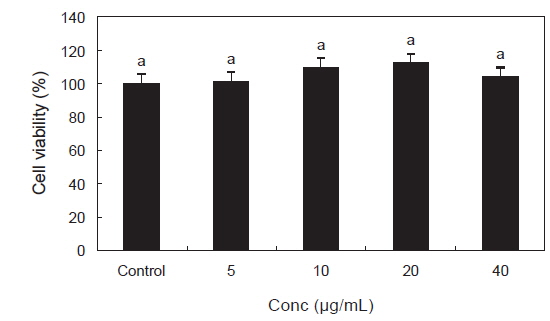
PYE가 RAW 264.7 대식세포의 세포생존율에 어떠한 영향을 미치는지 알아보기 위해 MTS assay를 이용하여 분석하였다. RAW 264.7 세포에 PYE를 5, 10, 20, 40 µg/mL의 농도로 처리한 후 24시간 동안 배양하였다. 그 결과 Fig. 1에서 나타난 바와 같이 PYE는 RAW 264.7 세포의 생존에 영향을 미치지 않는 것으로 나타났다.
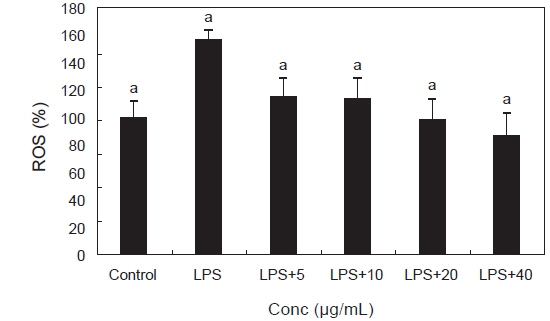
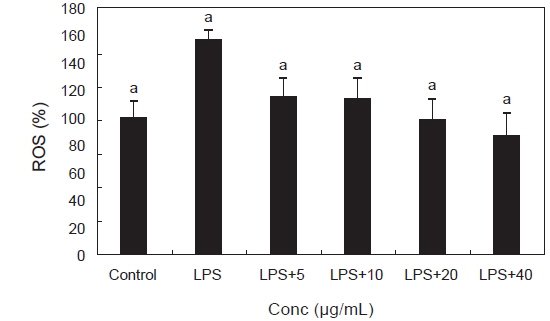
ROS는 인체 내 산소의 정상적인 대사작용에 의해서 자연스럽게 생기고 체내로 유입되는 세균이나 바이러스를 제거하는 면역체계로 세포신호와 항상성에 중요한 역할을 하지만 식품첨가물, 방사선, 알콜, 공해, 자외선이나 높은 열에 노출되는 것처럼 환경적인 스트레스 또는 지속적인 염증 반응에 의해 필요 이상으로 증가하여 질병과 노화의 주된 요인이 되고 있다(Devasagayam et al., 2004). PYE가 RAW 264.7 대식세포에 LPS를 처리하였을 때 생성되는 superoxide radical (O2-), hydrogen peroxide (H2O2), hydroxy radical (•OH) 등의 ROS 생성에 미치는 영향을 알아보았다. 대조군에 비해 LPS를 단독처리한 군에서 ROS가 유의적으로 증가하였고 PYE를 농도별로 처리한 군은 LPS 대조군에 비해 유의적으로 감소하는 경향을 나타내었다. 특히 PYE 농도가 높을수록 ROS 농도가 크게 감소되는 것을 볼 수 있었다(Fig. 2).
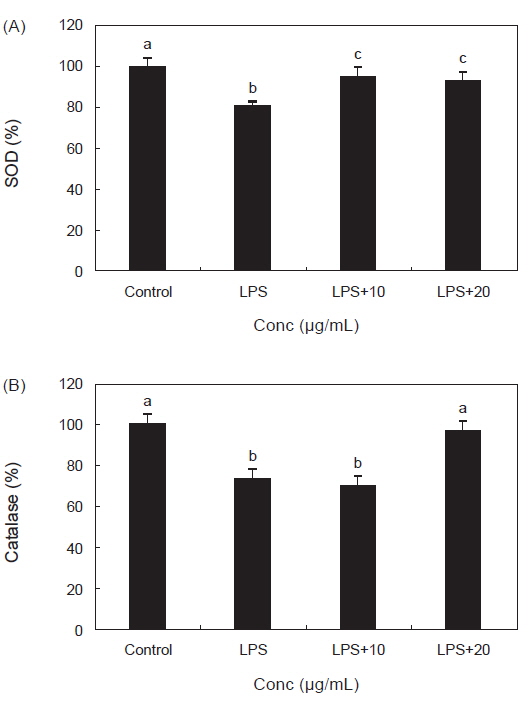
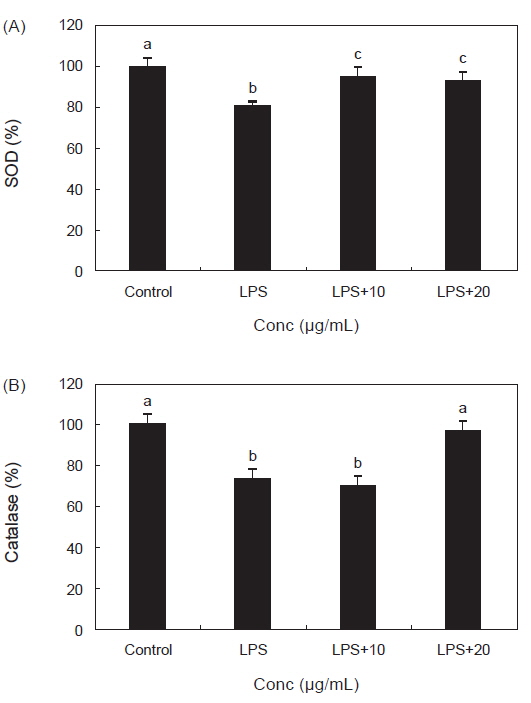
RAW 264.7 대식세포에서 LPS 처리 후 PYE를 농도별로 처리한 결과 SOD와 catalase의 활성에 미치는 효과는 Fig. 3에서 보는 바와 같다. Fig. 3 (A)를 보면 LPS 단독 처리 후 대조군보다 SOD활성이 유의적으로 감소 하였고 PYE를 10, 20 µg/mL 농도로 처리한 군에서는 LPS단독 처리군보다 유의적으로 SOD 활성이 증가하는 것이 확인되었다.
ROS인 O2- 는 산소호흡을 하는 호기적 세포에서 생성되며 산화적 스트레스로 인한 과다 생성시 암, 염증 및 여러 질병의 밀접한 관련이 있다(Sun et al., 2004). 이렇게 생성된 O2- 는 생물체 내에 가지고 있는 SOD에 의해 제거 된다. 대식세포의 LPS처리는 SOD의 활성을 감소시켰으나 PYE를 LPS와 동시 투여함으로서 SOD의 활성이 대조군 수준으로 회복시켰다. 이것은 PYE 투여가 LPS처리로 인하여 생긴 내독소의 산화적 스트레스에 대한 항산화 작용이 있는 것으로 사료 된다. Fig. 3 (B)에서 catalase의 효소 활성은 LPS만 처리 했을 때는 유의적으로 감소하였고 PYE 20 µg/mL처리시 유의적으로 증가하였다. 이 결과 PYE가 LPS를 처리한 대식세포에 대해 항산화작용을 하여 catalase 활성을 증가시켜 ROS 생성을 저해시켰음을 확인할 수 있었다. catalase는 조직 내의 SOD가 효소 반응에 의해 O2- 를 제거 한 후 TNF-α, IL-1β, IL-6 과 같은 염증성 인자인 cytokine 생산의 2차 messenger 역할을 하는 H2O2를 물로 분해 시켜 생체를 방어하는 항산화 효소이다(Yasui and Baba., 2006). 본 연구의 결과에서 LPS로 인한 산화적 스트레스에 대해서 PYE가 방어작용으로 SOD, catalase 등의 항산화 효소를 활성화 시켜 세포를 보호하는 효과를 가지는 것으로 사료된다.
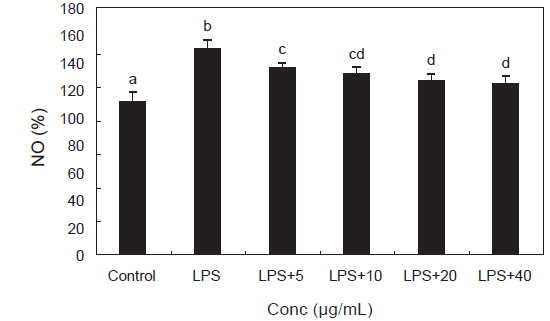
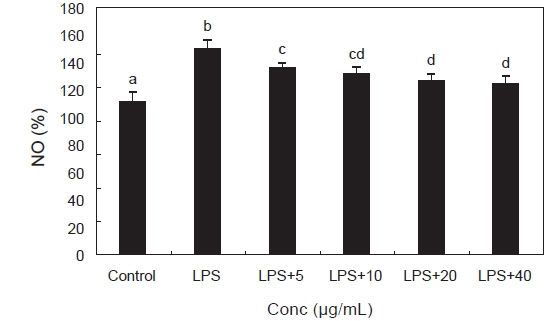
체내염증과정에서 과량의 NO 및 PGE2등의 염증인자는 유도형인 iNOS 및 COX-2에 의해 형성된다. NO를 형성하는 NOS (NO synthase)는 L-arginine을 L-citrulline으로 전환시키면서 NO를 만들어 내며 NO는 iNOS에 의해 생성된다(Schmidt and Walter., 1994). 이렇게 만들어진 NO는 염증반응을 매개하는 역할을 하고 생체 내에서 빠르게 superoxide와 반응하여 강력한 산화제인 peroxynitrite를 형성하여 염증 자극제로서 대식세포를 활성화 시켜(Ippouchi et al., 2002; Kang et al., 2000) NO의 생성을 현저히 증가시키고 다른 염증 물질들과 함께 과도한 염증을 유발시켜 조직의 손상을 일으킨다(Nathan, 1992; Pan et al., 2011). LPS로 자극된 RAW 264.7 대식세포에서 생성되는 NO에 대한 PYE의 억제효과를 확인하였으며 그 결과는 Fig. 4에서 보는 바와 같다. LPS 처리군은 대조군에 비하여 NO생성이 유의적으로 증가하였고 LPS와 PYE를 병행 처리한 군에서는 NO생성이 LPS 처리군에 비해 농도 유의적으로 감소하는 효과가 관찰되었다.
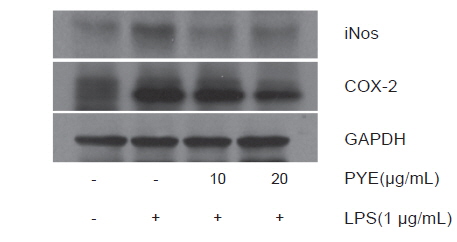
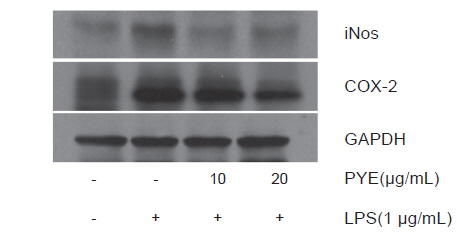
LPS로 자극된 RAW 264.7 대식세포에서 생성되는 COX-2, iNOS의 생성에 대한 PYE의 효과를 Western blot으로 분석하였다. Fig. 5에서 보는바와같이 iNOS의 단백질 발현은 LPS처리군에 비해 PYE를 처리한 군에서 억제되는 것을 확인하였다. 이것은 PYE 처리에 의한 NO 생성의 감소가 iNOS 발현 억제에 의한 것임을 의미한다. COX-2의 단백질 발현도 iNOS와 마찬가지로 LPS 처리군에 비해 PYE를 처리한 군에서 COX-2의 단백질 발현 역시 억제되는 것을 확인할 수 있었다. 염증은 생리적인 보호기전으로 방어기전을 활성화 시키거나 손상을 제한하지만 활성산소의 파괴적 성향으로 인하여 과다 생성되면 세포와 조직의 손상을 유도하고 더 나아가 만성질환 및 노화를 일으킨다(Azard et al., 2008). 여기서 생성되는 많은 양의 염증 유도 cytokine은 대식세포의 식세포작용을 과도하게 작용시켜 염증반응과 산화적 스트레스를 증가시킨다(Blatteis et al., 2004; Fullerton et al., 2013). NOS는 type Ⅰ, Ⅱ, Ⅲ의 3 종류로 구분되며 type Ⅲ인 iNOS는 평소에는 세포내에 존재하지 않으나 LPS, cytokine 및 박테리아 독소 같은 자극이나 NF-κB활성에 의해 유도되어 장시간 동안 다량의 NO를 생성한다(Nathan., 1992). 다른 염증인자인 COX-2는 arachidonic acid를 prostaglandins로 전환하는 cytokine, 자외선, 세균성 내독소 및 TNF등과 같은 여러 종류의 proinflammatory agent에 의해 과다하게 발현하여 염증 및 각종 퇴행성 질환에 관여한다(Botting, 2006; Wu et al., 2009).
>
PYE의 염증성 Cytokines의 생성 억제 효과
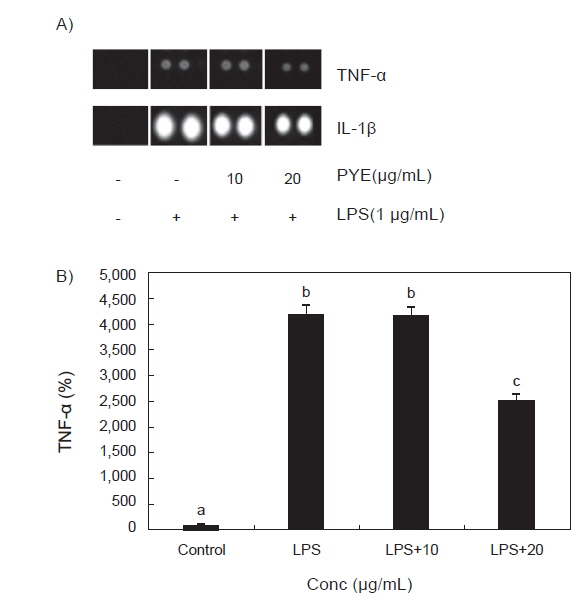
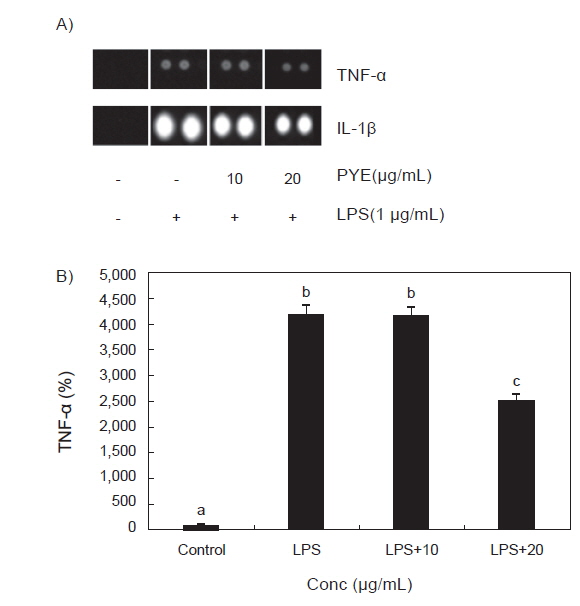
RAW 264.7 대식세포에 LPS를 처리한 후 PYE를 농도별로 처리하고 mouse cytokines array를 이용하여 각 군별 TNF-α와 IL-1β의 발현정도를 비교하였다.
Fig. 6에서 보는 바와 같이 TNF-α의 경우 LPS 단독 처리군에 비해 PYE를 20 µg/mL농도로 처리하였을 때 발현이 감소하였고 IL-1β 역시 LPS 단독 처리군에 비해 PYE를 처리하였을 때 농도유의적으로 감소하는 것이 관찰되었다.
LPS에 의해 자극된 대식세포는 TNF-α와 IL-1β의 생성을 유도하는데(Beutler and Ceramin., 1989) TNF-α의 과잉생산은 T cell의 수를 감소시켜 면역을 떨어뜨리고(Abul et al., 2007) 혈액과 조직 사이의 내피세포를 자극하여 여러 백혈구들을 염증부위로 이동시켜 각종 cytokine을 지속적으로 분비하여 미생물과 암세포에 대한 독성을 유지하거나 염증반응을 일으켜 조직을 손상시키기도 한다(Tracy et al., 1987). 따라서 TNF-α분비를 조절하는 것은 염증 반응 조절에 중요한 부분이다. Fig. 6에서 LPS처리 후 TNF-α의 발현이 LPS를 처리하지 않은 군보다 급격히 증가하였고 Fig. 6과 동일한 결과로 TNF-α의 발현이 PYE를 20 µg/mL로 처리하였을 때 유의적으로 낮은 수치를 보였으며, 이는 PYE가 TNF-α 발현에 관여하는 것을 보여주고 있다.
IL-1β는 대표적인 전 염증성 cytokine으로 면역 생물학적으로 볼 때 다른 cytokine보다 TNF-α와 밀접한 연관이 있다(Zhang et al., 1997). 급성기 염증 반응에서 TNF-α와 IL-1β의 발현은 급격히 증가하며 상호 상가효과를 나타낸다(Mathiak et al., 2000). Fig. 6에서도 대조군에 비하여 LPS처리 후 TNF-α와 IL-1β이 급격히 증가한 것을 알 수 있다.
NF-κB는 cytokine, LPS 와 산화적 스트레스등과 같은 여러 외부 자극에 의하여 활성화 되어 핵으로 이동한 뒤 면역과 염증반응에 관여한다. NF-κB는 세포질 속에서 불활성형으로 존재하다가 LPS, cytokine, ROS등에 의하여 IκB kinase가 활성화되고 IκB가 분해되면 활성화되어 핵으로 이동한다. 핵으로 이동한 NF-κB는 cytokine, iNOS, COX-2와 같은 염증반응 매개 물질들의 발현을 촉진하게 된다(Lawrence et al., 2001; Riehemann et al., 1999; Allenm and Tresini, 2000).
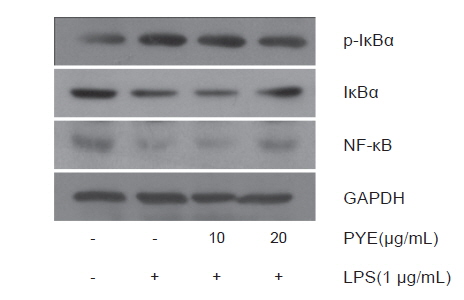
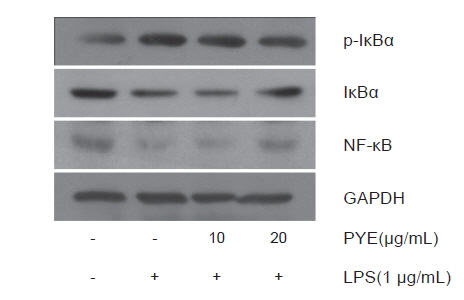
본 연구에서는 Fig. 7에서 보는 바와 같이 LPS처리에 의해 NF-κB와 IκB는 감소하지만 PYE 농도에 따라 증가하는 것을 확인하였고 LPS 처리에 의해 증가되는 IκB 인산화는 PYE처리로 감소하였다. 이것은 PYE 처리로 인하여 IκB 인산화가 억제되는 것으로 볼 수 있다. 따라서 LPS에 의해서 발생하는 염증 발현에서 PYE가 NF-κB 신호경로에 단계적으로 작용하여 저해효과를 나타내는 것으로 생각된다.
MAPK는 ERK, JNK, p38 MAPK로 구성되어 있으며 MAPK 신호경로들은 LPS로 유도되어 활성화된다. 활성화된 MAPK는 NF-κB 활성화에 관여하여 iNOS 발현 및 전염증성 물질에 발현에 관여한다(Celec, 2004). 그리고 Akt 또한 NF-κB 활성화뿐만 아니라 대식세포와 혈관세포에서 IκB활성화에 중요한 역할을 하며 여러 염증 유전자 발현에 관여하는 것으로 알려져 있다(MacMicking et al., 1997; Sheu et al., 2005).
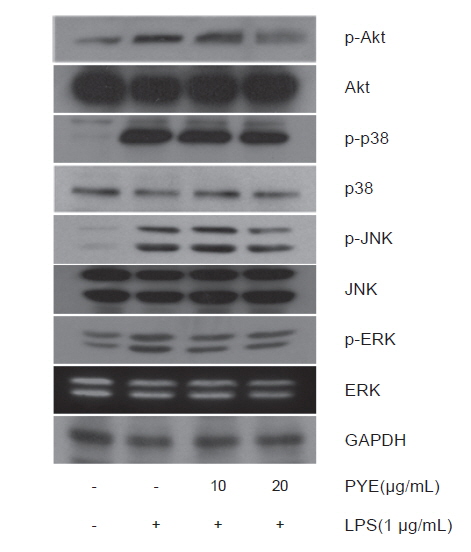
본 연구에서는 PYE가 NF-κB 활성화의 상위 신호 전달 경로인 MAPK 및 Akt 인산화에도 영향을 미치는지를 알아보기 위해 MAPK 및 Akt 신호경로에 관련된 단백질의 발현 수준을 western blot을 통해 확인하였다. Fig. 8에서 보는 바와 같이 Akt, p38, JNK, ERK의 활성화 형태인 p-Akt, p-p38, p-JNK, p-ERK는 LPS 처리시 증가하였으나 PYE 처리에 의하여 농도 유의적으로 감소하는 것이 관찰되었다. 따라서 PYE가 NF-κB pathway와 MAPK 및 Akt 경로에도 관여함으로써 NF-κB 활성화에 영향을 주는 것으로 보인다.
이상의 결과에서 PYE는 IκB 인산화를 억제시켜 NF-κB 활성을 저해하고 또한 NF-κB 활성화의 상위 신호 전달 경로인 MAPK 및 Akt 인산화에도 영향을 주어 NF- κB 활성을 저해하는 것으로 나타났다. 이를 통해 iNOS단백질과 COX-2 단백질 발현을 억제하여 염증성 매개체인 NO생성 감소와 염증촉진성 cytokines 생성 억제에 연관되어 있는 것으로 나타났으며 항산화 효소인 SOD, catalase의 활성을 증가시켜 산화적 스트레스를 감소시킴으로써 PYE가 LPS로 자극한 RAW 264.7 대식세포의 염증발현을 억제하는 것을 확인하였다. 따라서 PYE가 생체의 염증작용을 억제하는 효능을 가지고 있음을 알 수 있다.