자바리(
한국에는 남해안과 제주도 연안에 자바리, 붉바리(
양식산업은 한정된 사육 공간에서 대상어종을 고성장, 적정밀도로 건강하게 사육하여 단위면적당 생산량을 높이는 것이 경영면에서 중요하다. 이를 위해서는 양식대상 어종이 필요로 하는 적정 사육 환경을 유지시켜 주어야 한다. 특히 사육환경 중 수온은 수산동물의 성장, 면역, 성 성숙과 대사와 같은 생리학적 반응에 영향을 주는 중요한 요인이므로(Herzig and Winkler, 1986), 양식 대상 종의 치어를 생산하거나 양성하는데 있어서 우선적으로 적정 수온과 밀도 등 최적적응특성을 밝히는 것이 중요하다. 최근에도 수온과 밀도에 따른 치어의 성장(Sun and Chen, 2014)과 수온 등의 사육환경이 어류의 성장에 미치는 생리적 변화에 대한 연구가 진행되고 있으며(Qiang et al., 2012), 이러한 연구결과를 바탕으로 양식산업에 활용되고 있지만 국내 사육환경에 적합한 연구는 아직까지도 미흡한 실정이다.
따라서 본 연구는 새로운 고부가가치 양식대상 어종인 자바리의 상용화생산기술 개발을 목적으로 치어의 사육밀도 및 수온 등이 성장에 미치는 영향을 파악하여, 이 종의 최적 양식환경 조건을 알아보고자 하였다.
치어의 밀도 및 수온에 따른 특성조사의 실험어는 국립수산과학원 미래양식연구센터에서 생산한 치어(체중 6.9±1.1 g)를 이용하였다.
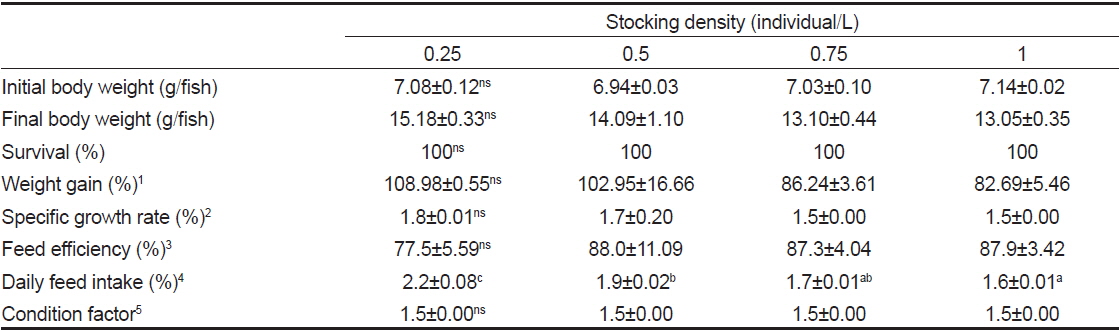
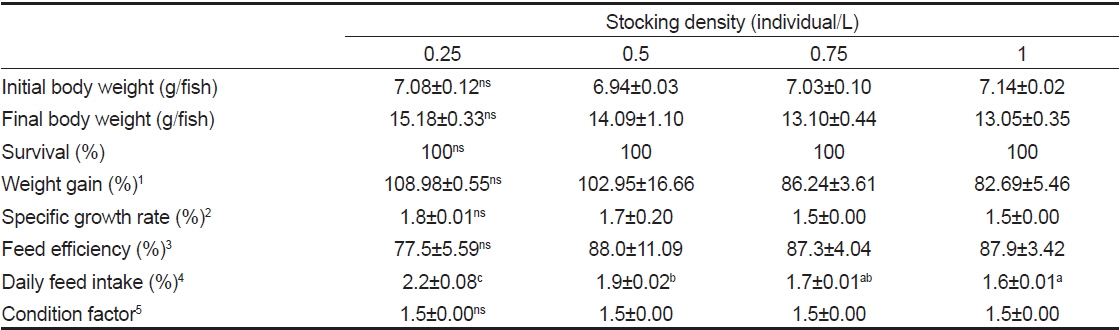
밀도에 따른 치어의 성장 특성 조사는 수량 70 L인 PP 수조에 각각 0.25, 0.5, 0.75 및 1마리/L 실험구를 설정하여 2반복으로 실시하였다. 실험기간 동안 수온은 22.7-23.8℃, 유수량은 5 L/min, 광조건은 조도 15 Lux에서 12시간 명기, 12시간 암기(L:D=12:12) 조건에서 6주간 사육하였다. 또한 액화산소를 이용하여 용존산소가 10.0-12.9 mg/L 범위가 되도록 유지하였다. 사육어의 사료공급은 상업용 EP 사료(유레카, 넙치 3호)를 1일 2회 만복이 되게 공급하였다.
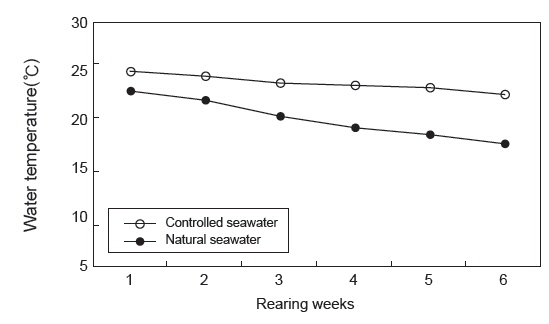
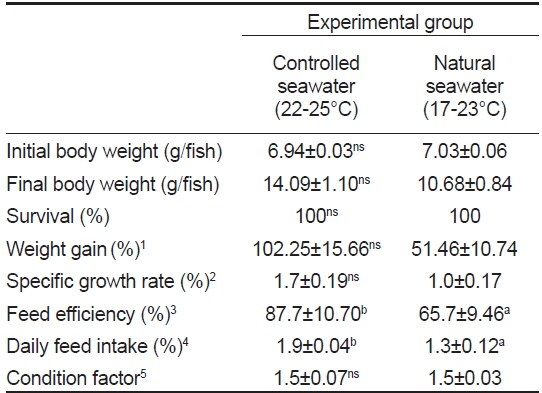
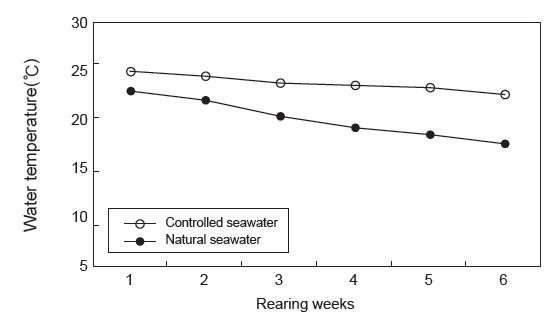
수온에 따른 치어의 성장 특성 조사는 수량 70 L인 PP 수조에 가온 및 자연해수 실험구에 각각 35마리씩 실험구별로 2반복으로 가온에 따른 성장 특성을 6주간 조사하였다. 실험기간 동안 사육수온은 가온해수 처리구(22-25℃) 및 자연해수 처리구(17-23℃)로 구분하고(Fig. 1), 유수량, 광주조건, 용존산소 및 먹이 공급은 밀도 실험과 동일한 방법으로 수행하였다.
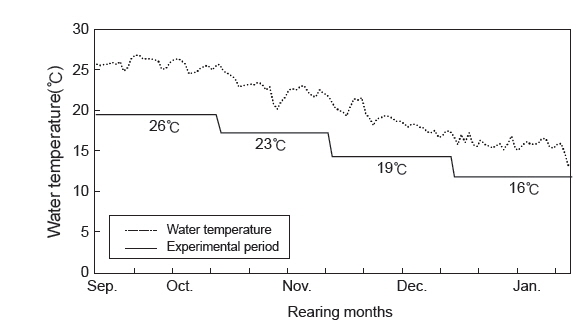
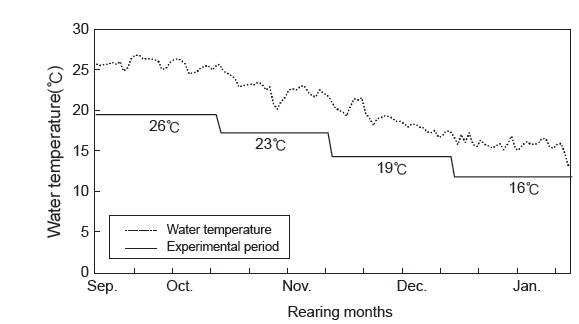
자연해수를 이용하여 사육 시기별 수온 변화에 따른 치어의 성장 특성을 조사하기 위해 실험어의 평균 체중이 19.7±5.2 g (당년어,
성장률 조사를 위해 실험어는 MS-222 (Sigma-Aldrich, St. Louis, MO, USA)를 이용하여 마취 후 개체별 체중을 측정하였다. 밀도 및 가온에 따른 성장 특성은 2주 간격, 시기별 수온변화에 따른 성장 특성은 1개월 간격으로 전수조사를 하였다. 실험 종료 후 각 실험구별 증체률(weight gain, WG), 일간성장률(specific growth rate, SGR), 사료효율(feed efficiency, FE), 사료섭취률(daily feed intake, DFI), 생존율(survival) 및 비만도(condition factor, CF) 값을 계산하였다.
모든 결과의 통계처리는 SPSS-통계패키지(version 10.0)를 이용하여 One-way ANOVA-test을 실시하여 Duncan's multiple range test 평균값 간의 유의성을 검정(
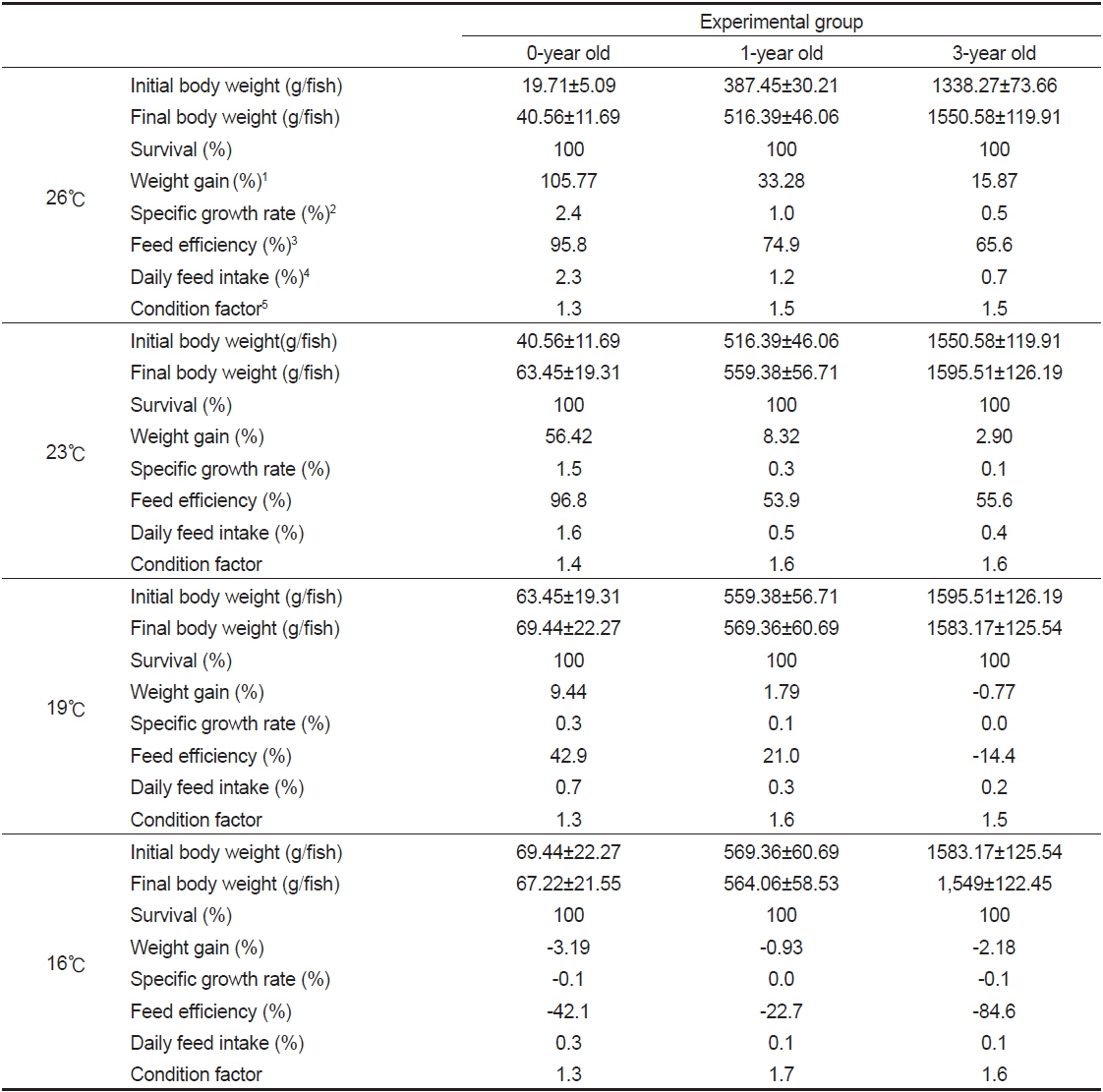
밀도에 따른 치어의 성장 특성을 조사한 결과(Table 1)에서, 생존율 및 비만도(CF)는 모든 실험구간에서 유사하였다(
Growth performance of juvenile longtooth grouper Epinephelus bruneus with different density
가온에 따른 치어의 성장 특성을 조사한 결과(Table 2)는 가온과 자연해수 실험구에서의 증체율(WG)은 각각 102.3±15.7, 51.5±10.7%, 일간성장률(SGR)은 각각 1.7±0.2, 1.0±0.7%로 가온해수가 자연해수 보다 2배 정도 높았다(
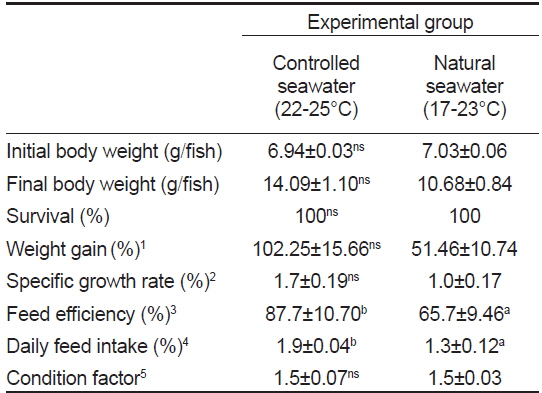
Growth performance of juvenile longtooth grouper Epinephelus bruneus at controlled seawater and natural seawater
자연해수의 시기별 수온 변화에 따른 치어의 성장 특성을 조사한 결과(Table 3)에서 생존율은 모든 실험구에서 100%이었다. 치어 단계인 0년어는 사육수온 26, 23, 19 및 16℃에서 증체율(WG)은 각각 105.8, 56.4, 9.4 및 -3.2%으로 수온이 낮을수록 낮아지는 경향이 뚜렷하였고, 0년어가 대조구인 1년어와 3년어에 비해 높았다. 특히 사육수온 26℃에서 1년어와 3년어가 각각 33.3와 15.9%에 비해 0년어가 105.8%로 높았다.
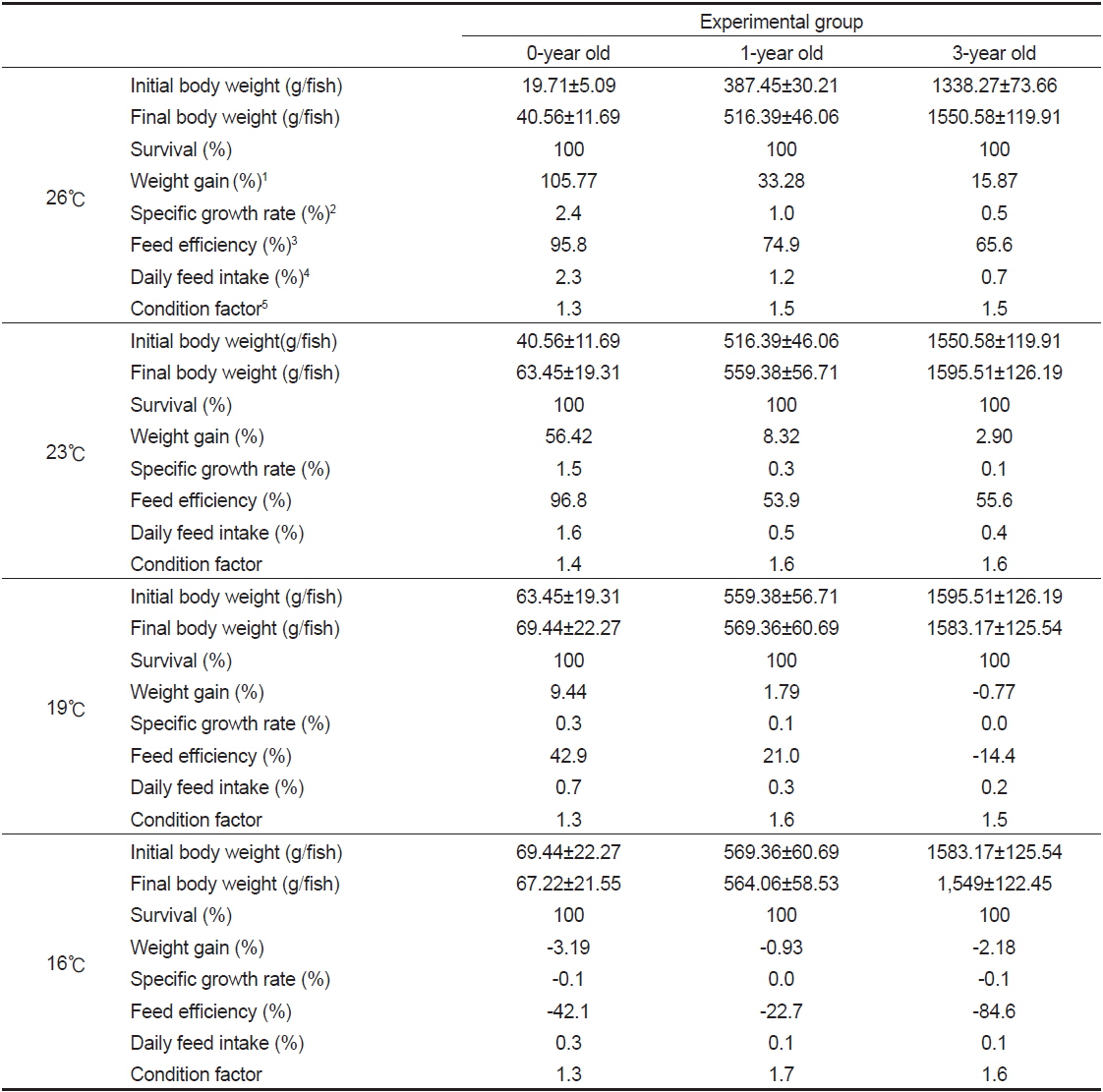
Growth performance of juvenile Longtooh grouper Epinephelus bruneus with body size and water temperature
일간성장률(SGR)은 0년어가 각각 2.4, 1.5, 0.3 및 −0.1%으로 수온이 낮을수록 낮아지는 경향이 뚜렷하였고, 1년어와 3년어에 보다 높았다. 특히 사육수온 26℃에서 1년어와 3년어가 각각 1.0, 0.5%에 비해 0년어가 2.4%로 높았다.
사료효율(FE)은 0년어가 사육수온 26℃와 23℃에서 95.8와 96.8%로 비슷한 값을 나타낸 반면, 19℃에서는 26℃와 23℃에 비해 절반 정도로 낮은 42.9%였고, 16℃에서는 −42.1%로 가장 낮았다. 0년어가 1년어와 3년어에 비해 19℃ 이상에서 높은 값을 보인 반면, 16℃에서는 0년어가 1년어에 비해 낮은 값을 보였다. 사육수온 26℃에서는 1년어와 3년어가 각각 74.9, 65.6%에 비해 0년어가 95.8%로 높았다.
사료섭취률(DFI)은 0년어가 각각 2.3, 1.6, 0.7 및 0.3%로 수온이 낮을수록 낮아지는 경향이 뚜렷하였고, 1년어와 3년어보다는 높은 값을 보였다. 특히 사육수온 26℃에서 1년어와 3년어가 각각 1.2, 0.7%에 비해 0년어가 2.3%로 2배 정도 높았다.
비만도(CF)는 0년어가 각각 1.3, 1.4, 1.3 및 1.3으로 수온에 따른 차이는 없었고, 1년어와 3년어에서는 각각 1.5-1.7, 1.5-1.6 범위로 유사하였다.
1년어와 3년어의 증체율(WG), 일간성장률(SGR), 사료섭취률(DFI) 및 사료효율(FE)의 변화는 0년어와 비슷한 결과로 수온이 낮아질수록 낮아지는 경향이었다. 특히 증체율(WG), 일간성장률(SGR), 사료섭취률(DFI) 및 사료효율(FE)의 값은 어체 크기가 작을수록 높았다.
어류 양식에서 최적사양관리는 사육수온과 밀도에 따른 성장과 사료효율 등 사육환경의 적응특성을 파악하는 것이 우선 되어야 한다. 어류 사육에서 밀도가 높아짐에 따라 성장이 낮아지고 폐사율이 높아지는 현상은 대부분 공통된 현상이다(Trzebiatowski et al., 1981; Morrissy, 1992). 쥐노래미(
사육환경 중 수온은 어류의 신진대사와 성장 등에 영향을 미치고, 어류의 생리적 기능은 수온의 영향에 의한 효소활성으로 조절된다(Tandler et al., 1989; Iwata et al., 1994). 자바리 치어를 수온이 23℃ 이하로 내려가는 10월 하순부터 6주 동안에 자연수온구 보다 약 4℃ 정도 높은 가온해수로 사육하였을 때 자연해수에 비해 증체율, 일간성장률, 사료효율 및 사료섭취률이 높았다. 특히 본 연구에서 가온해수가 수온이 내려가는 대조구에 비해 성장이 높게 나타나 성장만을 고려한다면, 가온에 의한 치어의 중간육성이 필요한 조건이라고 할 수 있다.
난류성 어종인 붉은 쏨뱅이(
홍민어의 일간성장률은 26, 23 및 20℃ 순으로 수온이 높을수록 빠른 성장을 하였고, 일간섭식률에서도 26℃에서 가장 높았다(Choi et al., 2002). Orange-spotted grouper
자바리 양식은 상용화 크기까지 양성기간이 길어 양식기간을 단축하지 않으면 다른 어종의 양식에 비해 경쟁력이 없는 실정이다. 본 연구에서 자바리는 치어 단계부터는 공식, 질병 등에 대한 감모가 없었서, 앞으로 적정 사육수온 조건으로 양성기간을 단축하고 열이용 히트펌프 시스템과 산소발생기 등의 사육 시스템 구축 등으로 양식 경영개선이 선행된다면 육상수조의 적정 양식이 가능할 것으로 전망이 된다.