Aquaculture continues to be the fastest growing animal food-producing sector. As aquaculture production becomes increasingly intensive, fish feed will be an important factor in increasing the productivity and profitability of the sector (Jamu and Ayinla, 2003). Considering that feed is the single costliest item in aquaculture practice, accounting for > 50% of the production cost, feed management determines aquaculture viability (Shang, 1992; Craig and Helfrich, 2002; Jamu and Ayinla, 2003). In this regard, nutrition research that reduces the cost of fish feeds without reducing their efficacy will be crucial to the successful development of aquaculture.
The use of crop and animal waste as animal feed is of considerable interest because it produces an environmental benefit besides reducing the cost of animal production (Samuels et al., 1991; Westendorf et al., 1998; Myer et al., 1999; Westendorf, 2000). In many tropical regions of Asia and Africa, waste from crops, agro-industry and animal production are commonly used in aquaculture. Such waste has been used as a feed ingredient, supplementary feed, or pond fertilizer (Ravishankar and Keshavanath, 1986; Wohlfarth and Hulata, 1987; Subosa, 1992; Tacon, 1993, 1994).
Coffee production for human consumption gives rise to a number of byproducts that can be used as feedstuff. These include leaves, pulp from the bean, coffee residues, coffee meal, and spent coffee grounds (CG). The coffee bean can be used to make one of the most popular beverages in the world, and considerable amounts of coffee bean are processed daily, leading to large quantities of byproducts that could be used as dietary ingredients (Heuzé and Tran, 2011). Coffee byproducts are rich, low-cost sources of carbohydrates, proteins and bioactive compounds. Roasted coffee contains ~10% protein, 11–17% lipid, 38–42% carbohydrates, and 4.5–4.7% minerals (Esquivel and Jiménez, 2012). In addition, coffee beans comprise a complex array of components with the potential to influence metabolic processes. The most influential compounds are alkaloids (caffeine and trigonelline), phenolic compounds (chlorogenic acids), and diterpenes (Oestreich-Janzen, 2010). These coffee components are believed to have potential as antioxidants and free radical scavengers. CG are the residue of soluble coffee processing. Although the volume of coffee residues has decreased due to improvements in soluble coffee processing, their disposal remains a matter of great concern. Hence, the development of rational uses for roasted coffee residues is necessary. Many attempts have been made to use roasted coffee residues as a fertilizer, animal feed, or a substitute for industrial materials.
There are several reports on the use of coffee pulp as a feed ingredient for fish (Rojas, 2002; Moreau et al., 2003); however, to the best of our knowledge, no study on the use of spent CG in fish feed has been reported. Therefore, the present study aimed to evaluate CG as a potential feedstuff for olive flounder in terms of growth performance, feed utilization, body composition, and antioxidant enzyme activity.
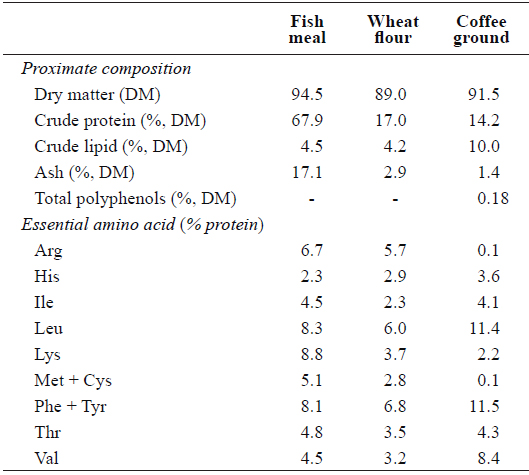
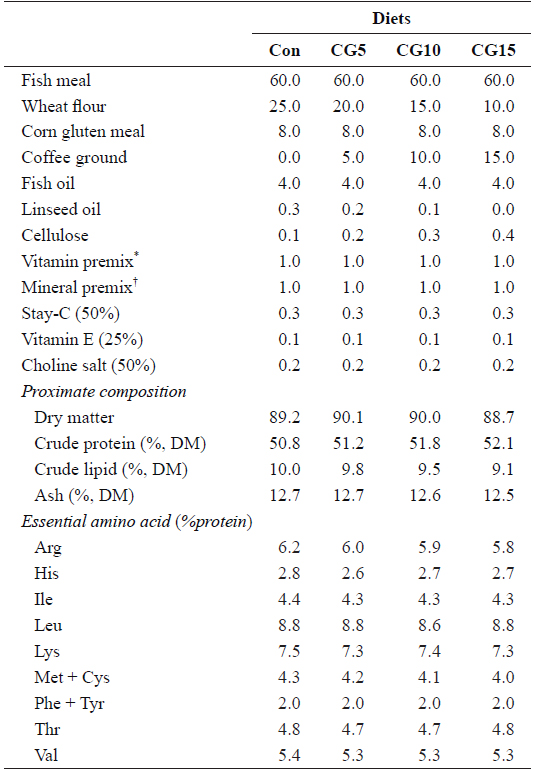
Proximate and essential amino acid compositions of the major dietary ingredients used in this study are shown in Table 1. A basal diet was formulated and used as a control and three other test diets were prepared to contain 5, 10 or 15% CG (designated as Con, CG5, CG10, and CG15, respectively) (Table 2). Roasted Arabica coffee residue was used as the source of CG in this study. Coffee beans were roasted at 180–210°C for 15 min, and the drip water temperature was set to 90°C. Fish meal was used as the primary protein source and fish oil was used as the lipid source. All ingredients were thoroughly mixed with 30% distilled water and pellets were prepared using a laboratory moist-pelleting machine. The pellets were dried at room temperature for 48 h, crushed into the desired particle size and stored at -30°C until use.
[Table 1.] Proximate composition and essential amino acid profile of the major dietary ingredients
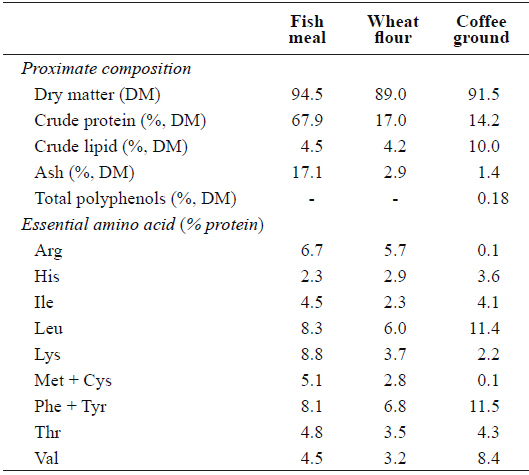
Proximate composition and essential amino acid profile of the major dietary ingredients
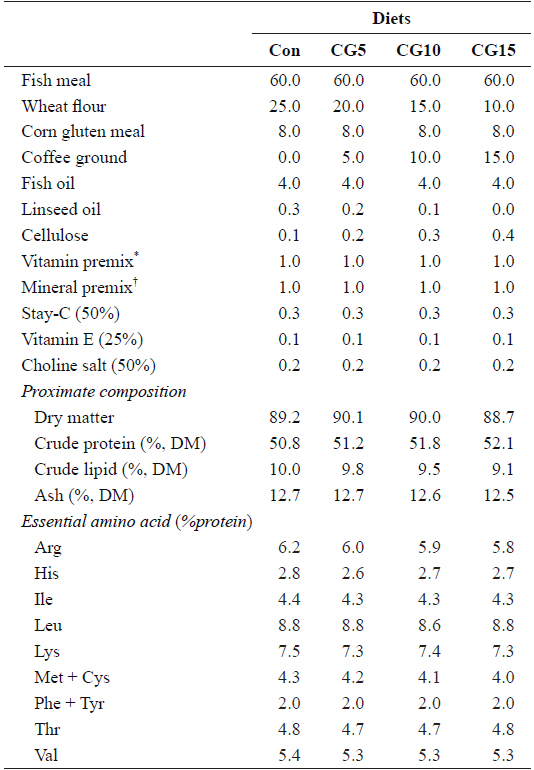
Formulation, proximate composition and essential amino acid profile of the experimental diets
Flounder were transported from a local hatchery (Namhae, Korea) to the Gangneung-Wonju National University, Korea. The fish were acclimated to laboratory conditions and fed a commercial diet twice daily for 2 weeks prior to starting the feeding trial. Two hundred forty fish (104 ± 0.7 g/fish) were randomly distributed into twelve 400 L polyvinyl circular tanks at a density of 20 fish per tank. Each experimental diet was fed to triplicate groups of fish to apparent satiation twice daily (09:00 and 17:00) for 10 weeks. A flow-through system was used, and each tank was supplied with filtered seawater at a flow rate of 4 L min-1 and aeration to maintain sufficient dissolved oxygen. The water temperature during the experiment was maintained at 21.5 ± 3.59°C and the photoperiod was maintained on a 12:12 h light:dark schedule.
>
Sample collection and analyses
At the end of the feeding trial, all fish in each tank were counted and collectively weighed to calculate growth parameters and survival. Five fish per tank (15 fish per dietary treatment) were randomly captured and anesthetized with tricainmethanesulfonate (MS222, Sigma, St. Louis, MO, USA) (100 mg L-1). Blood samples were then collected from the caudal vein using heparinized syringes. Plasma was then separated by centrifugation at 7,500 rpm at 4°C for 10 min and stored at -70°C to determine alkyl radical and superoxide radical-scavenging activities and blood biochemical parameters. After collecting blood, dorsal muscle and liver were sampled to determine the radical-scavenging activities, liver total protein (TP), catalase activity (CAT), total antioxidant capacity (TAC), and glutathione peroxidase activity (GPx), as mentioned above. In addition, another set of five fish per tank was sampled and stored at -25°C for muscle and liver proximate composition analyses.
>
Proximate composition analysis
Proximate composition of the diet, and dorsal muscle and liver samples were analyzed according to standard methods (AOAC 1995). Crude protein content was determined by the Kjeldahl method using the Kjeldahl System (Buchi, Flawil, Switzerland). Crude lipid content was analyzed by ether extraction in a Soxhlet extractor (SER 148, VELP Scientifica, Milano, Italy). Moisture was determined by oven-drying at 105°C for 6 h, and ash content was determined after combustion at 550°C for 4 h in a muffle furnace. The amino acid composition of the major ingredients and experimental diets was determined using an automated amino acid analyzer (Hitachi, Tokyo, Japan) following acid hydrolysis with 6 N HCl (reflux for 23 h at 110°C).
>
Blood biochemical parameters
Plasma total protein, glucose, glutamate oxaloacetate transaminase (GOT), glutamate pyruvate transaminase (GPT), and total cholesterol concentrations were determined using a commercial clinical investigation kit (Asan Pharmaceutical Co., Seoul, Korea).
>
Antioxidant enzyme activities
Liver CAT and TAC activities were analyzed using commercially available kits (Bioassay Systems Hayward, CA, USA) following the manufacturer’s instructions.
GPx activity was assayed using a commercial kit (Bioassay Systems Hayward, CA, USA). In this assay, cumenehydroperoxide was used as a peroxide substrate (ROOH), and glutathione reductase (GSSG-R) and NADPH (β-nicotinamide adenine dinucleotide phosphate, reduced) were included in the reaction mixture. The change in absorbance at 340 nm due to NADPH oxidation was monitored to assess GPx activity. Briefly, 50 μL of plasma were added to 40 μL of the reaction mixture and incubated for 15 min; 10 μL of cumenehydroperoxide were then added and the OD1 was read at 340 nm. After 5 min of incubation, the OD2 at 340 nm was read using an enzyme-linked immunosorbent assay (ELISA) reader (Mannedorf, Zurich, Switzerland). GPx activity was calculated as U/L.
Dorsal muscle and liver samples were homogenized (Wiggenhauser, Berlin, Germany) with extract buffer in 5 mM Tris-HCl and 35 mM glycine (pH 8.4) followed by centrifugation at 13,000 g for 10 min at 4°C. The supernatant was then used to determine radical-scavenging activity.
Alkyl radicals were determined by the 2,2-azobiz-(2-amidinopropane)-hydrochloride (AAPH) method. The phosphate-buffered saline (pH 7.4) reaction mixtures included 10 mmol/L AAPH, 10 mmol/L 4-POBN, and known concentrations of sample (100 μg/mL), which were incubated at 37°C in a water bath for 30 min (Hiramoto et al., 1993) and then transferred to a capillary tube. The spin adduct was recorded using a spectrometer (JEOL LTD., Tokyo, Japan). The measurement conditions were as follows: modulation frequency, 100 kHz; microwave power, 10 mW; microwave frequency, 9,441 MHz; magnetic field, 336.5 ± 5 mT; sweep time, 30 s. Alkyl radical-scavenging activity (RSA) was calculated using the following equation:
RSA (%) = [(Acontrol– Asample) / Acontrol] × 100,
where Asample is the relative peak height of radical sample signals and Acontrol is the relative peak height of radical control signals.
Superoxide radicals were generated using an ultraviolet (UV)-irradiated riboflavin/EDTA system (Guo et al., 1999). The reaction mixture containing 0.3 mM riboflavin, 1.6 mM EDTA, 800 mM DMPO, and the indicated sample concentration was irradiated for 1 min under a UV lamp at 365 nm. The reaction mixture was then transferred to a 100 μL quartz capillary tube for measurement by ESR spectrometry. The experimental conditions were: magnetic field, 336.5 ± 5 mT; power, 10 mW; modulation frequency, 9.41 GHz; amplitude, 1 × 1,000; sweep time, 1 min. The superoxide radical-scavenging ability of each sample was calculated as follows:
Superoxide radical scavenging activity (%) = (1-A/A0) × 100,
where A and A0 are the relative peak heights of radical signals with and without sample, respectively.
Data were subjected to one-way analysis of variance (ANOVA) and if significant (
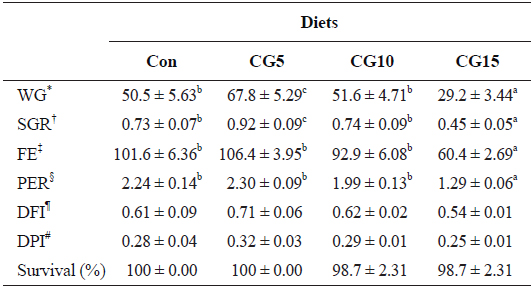
Growth performance and feed utilization of olive flounder fed the experimental diets are shown in Table 3. The results showed significant (
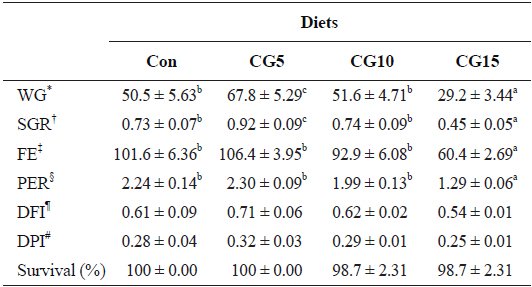
Growth performance and feed utilization of olive flounder Paralichthys olivaceus (104 ± 0.7 g/fish) fed the experimental diets for 10 weeks
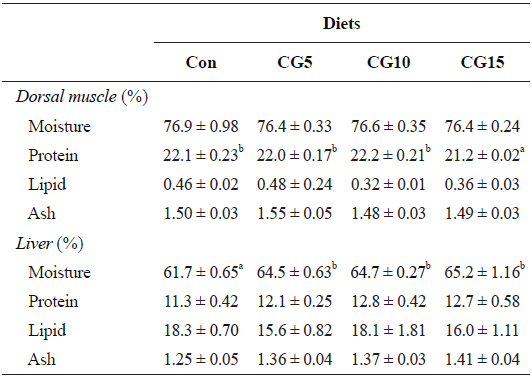
Dorsal muscle and liver compositions were significantly influenced by dietary CG (Table 4). Significantly lower muscle protein content was found in fish fed CG15 diet compared the other dietary treatments. However, muscle lipid, moisture and ash contents did not significantly differ among treatments. Also, the results showed significant increase of liver moisture content in all CG fed fish compared to the control.
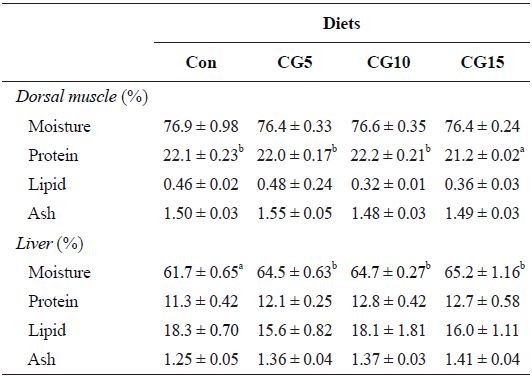
Proximate composition of dorsal muscle and liver of olive flounder Paralichthys olivaceus fed the experimental diets for 10 weeks
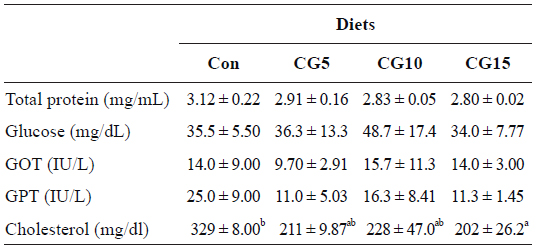
Results of biochemical analysis of blood showed the significant decrease of plasma total cholesterol concentration in fish fed CG15 diet in comparison to control group (Table 5). However, plasma total protein, glucose, GOT and GPT concentrations were not significantly influenced by dietary CG.
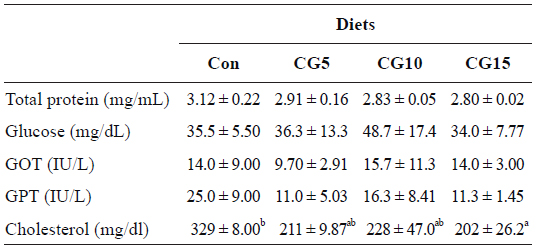
Plasma biochemical parameters of olive flounder Paralichthys olivaceus fed the experimental diets for 10 weeks
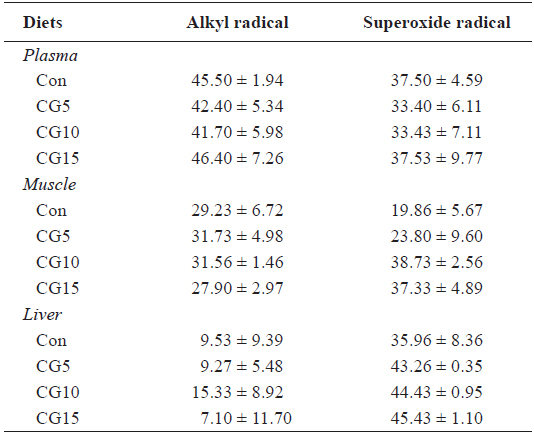
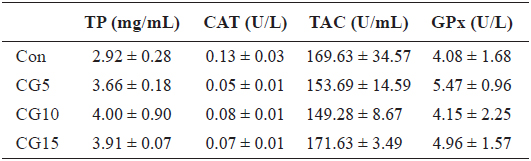
Radical scavenging activity of plasma, dorsal muscle and liver is provided in Table 6. The results showed that inclusion of CG in diets for olive flounder has no significant effects on alkyl and superoxide radicals scavenging activity. Also, no significant changes were found in liver total protein content, TAC or CAT and GPx activities among dietary treatments (Table 7).
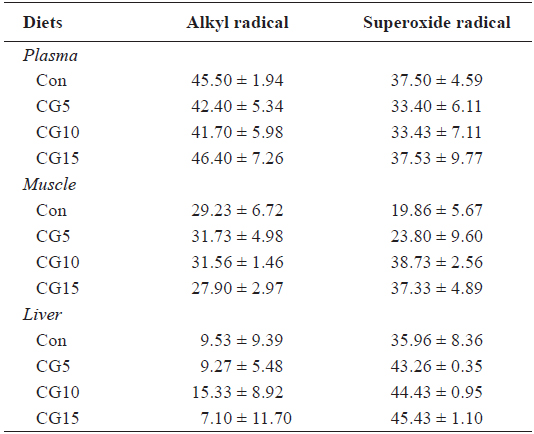
Radical scavenging activity (%) of the muscle, plasma and liver in olive flounder Paralichthys olivaceus fed the experimental diets for 10 weeks
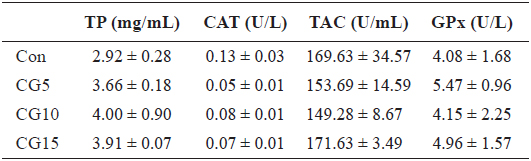
Liver total protein (TP), catalase activity (CAT), total antioxidant capacity (TAC) and glutathione peroxidase activity (GPx) of olive flounder Paralichthys olivaceus fed the experimental diets for 10 weeks
In the present study significant enhancement in growth performance of olive flounder was observed by dietary inclusion of 5% CG. It has been reported that dietary supplementation of polyphenol-rich plant products improves growth performance by influencing the gut microbiota (Fiesel et al., 2014). Dueñas et al. (2015) suggested that dietary polyphenols exert prebiotic-like effects by stimulating the growth of beneficial bacteria (i.e., lactobacilli and bifidobacteria) and inhibiting the pathogenic bacteria. The findings of this study indicate that up to 10% CG can be included in the diet of olive flounder without adverse effects on growth performance and feed utilization. To the best of our knowledge, no study on the utilization of CG in fish feed has been reported. However, several studies have examined other coffee byproducts (
In the present study, whole-body protein content was significantly reduced in fish fed the CG15 diet. This is in correspondence with the results obtained for PER. Indeed, the use of coffee byproducts in animal feed has been reported to interfere with nutrient availability and absorption in the gastrointestinal tract (Cabezas et al., 1987; Aregheore, 1998; Rathinavelu and Graziosi, 2005). As mentioned above, coffee byproducts contain high levels of tannins, which affect feed digestibility and nitrogen utilization in animals (Marcel et al., 2011). Tannins have been reported to reduce protein digestibility and assimilation either through interaction with dietary protein or through inhibition of digestive proteases (Reed, 1995; Bravo, 1998). Furthermore, evidence suggests that nitrogen availability can be affected by the formation of protein complexes by tannins.
In this study dietary inclusion of CG resulted in a reduction in plasma cholesterol concentration and a significant difference was observed between groups fed the control and CG15 diets. It has been shown that some dietary polyphenols, such as coffee polyphenols exert hypocholesterolemic effects. Meguro et al. (2013) reported that the addition of coffee polyphenols to a high-cholesterol diet for zebrafish
Antioxidant enzymes include superoxide dismutase (SOD), CAT and GPx, and constitute the first line of antioxidant enzymatic defense. SOD catalyzes the dismutation of superoxide radicals to hydrogen peroxide and oxygen, and CAT catalyzes the breakdown of hydrogen peroxide to water and molecular oxygen. GPX, a selenium-dependent enzyme, decomposes peroxides using the peptide glutathione as a co-substrate (Halliwell, 2006). Caffeine, chlorogenic acid (CGA), cafestol, trigonelline, and kahweol, all found in coffee, are believed to have significant potential as antioxidants and free radical scavengers. The roasting process influences the antioxidant properties of coffee as total antioxidant activity decreases with increased roasting temperature and duration (del Castillo et al., 2005). Polyphenol levels are reduced by 10% and those of CGA from 15% to 5% in dark versus lighter roasts (Cammerer and Kroh, 2006). Two low-molecular-mass products, CGA and caffeine, remain in dark roasts and have a protective effect (Daglia et al., 2000). Many researchers believe that Maillard reaction products, which are also strong antioxidants, are formed during the roasting process (Nicoli et al., 1997; Del Castillo et al., 2002; Nebesny and Budryn, 2003). Yen et al. (2005) evaluated the antioxidant properties of roasted coffee residues and found them to have excellent potential for use as a natural antioxidant source. Accordingly, in the present study, the changes in antioxidant enzyme activities following CG administration were evaluated. However, we found no significant change in TAC, CAT, GPx activity, or radical-scavenging activity among the diets. Del Castillo et al. (2002) reported that the degree of roasting affects the decrease in the antioxidant activity of coffee, which is associated mainly with the degradation of CGA. The antioxidant activity of roasted coffee is dependent on the roasting temperature and duration as well as the roasting method used (Nebesny and Budryn, 2003). In addition, the concentrations of coffee constituent compounds, such as CGA and polyphenols, which contribute to its antioxidant activity, vary geographically (Mullen et al., 2013). Therefore, further studies are required to examine the antioxidant activity of CG in other fishes.
In conclusion, the findings of this study indicate that CG can be used as a feedstuff to enhance the growth of olive flounder. However, CG at > 10% of the diet dry weight should not be used, as inclusion of higher levels resulted in negative effects on growth performance and feed utilization.