Cyprinids are quantitatively the most important group of teleost fish cultivated around the world. The global fishery and aquaculture production of common carp reached approx. 3.3 million tons in 2009 (FAO, 2010). Throughout more than 2000 years of rearing common carp the cumulative founder effects, genetic drift, and natural or artificial selection have led to the formation of many distinct strain of this fish (Hulata, 1995; Bakos and Gorda, 1995). Israeli carp
To develop cost-effective and practical dietary formulations for target fishes, we must know their essential nutrient requirements. Dietary protein is the most important factor affecting the growth performance of fish and feed costs (NRC, 1993). Generally, fish growth can be improved by increasing dietary protein level. Protein utilization for growth may be improved by partially replacing dietary protein with lipid and/or carbohydrate to produce a protein-sparing effect. However, excessive energy in diets can lead to increased body lipid deposition and growth reduction of fish due to a lack of necessary nutrients for growth resulting from a reduction in feed consumption (Ali and Jauncey, 2005; Daniels and Robinson, 1986). On the contrary, insufficient non-protein energy in diet causes protein waste as the proportion of dietary protein used for energy increases, and ammonia excreted after amino acids are metabolized can reduce water quality (Yang et al., 2002). Therefore, it is important to increase dietary protein utilization for body protein synthesis rather than for energy purposes. Higher energy levels generally come from increased dietary lipid as lipid is an energy-dense nutrient and readily metabolized by fish. The present study, therefore, was conducted to investigate the effect of dietary protein and lipid levels on the growth and feed utilization of Israeli carp
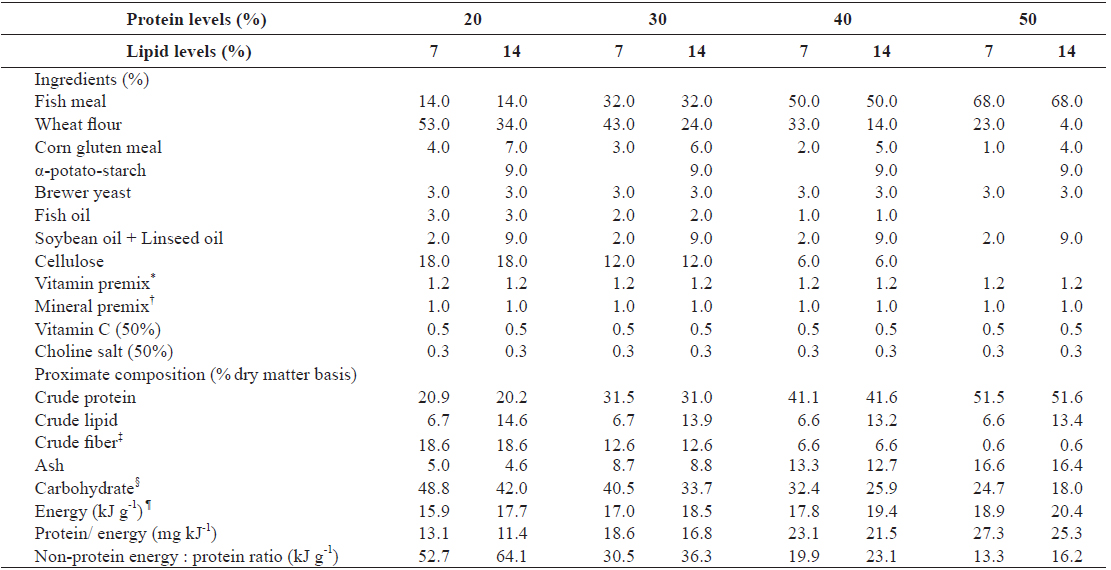
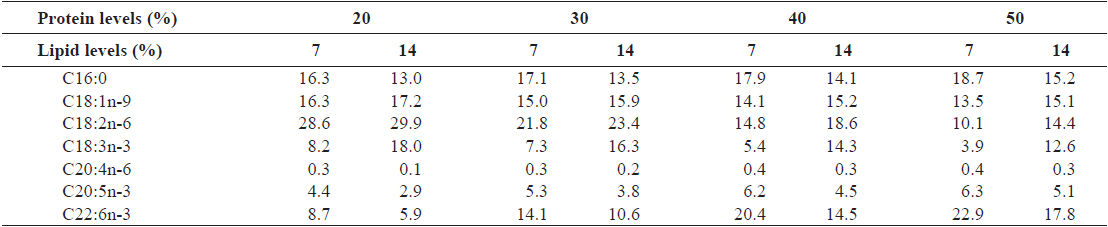
A 4 × 2 factorial design with three replicates was used. Eight experimental diets were formulated to contain four protein levels (20, 30, 40 and 50%) and two lipid levels (7 and 14%). Ingredient and proximate composition of the experimental diets are presented in Table 1. Fish meal as the primary protein source, fish oil and mixture of soybean and linseed oil as lipid sources and wheat flour as carbohydrate source were used. Proportion of fish oil decreased with an increase in dietary fish meal level. Fatty acid compositions of the experimental diets are shown in Table 2. The experimental diets were pelletized by a laboratory pellet machine after 400 g of water was mixed with 1 kg of ingredients and dried overnight at room temperature. All pellets were stored at -30°C until use.
[Table 1.] Ingredients and proximate composition of experimental diets
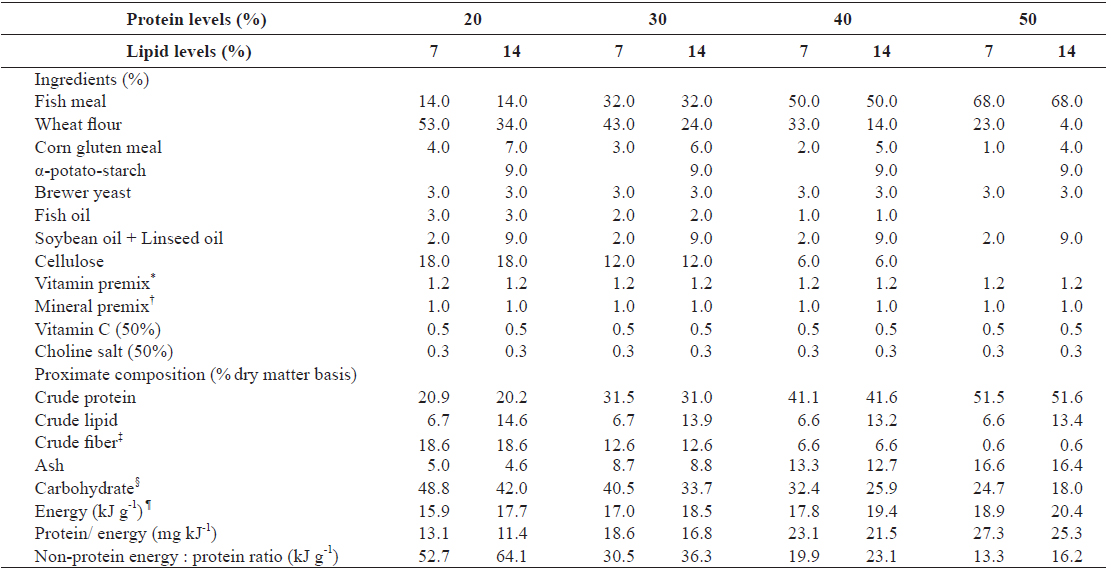
Ingredients and proximate composition of experimental diets
[Table 2.] Major fatty acid composition (% of the total fatty acids) of the experimental diets
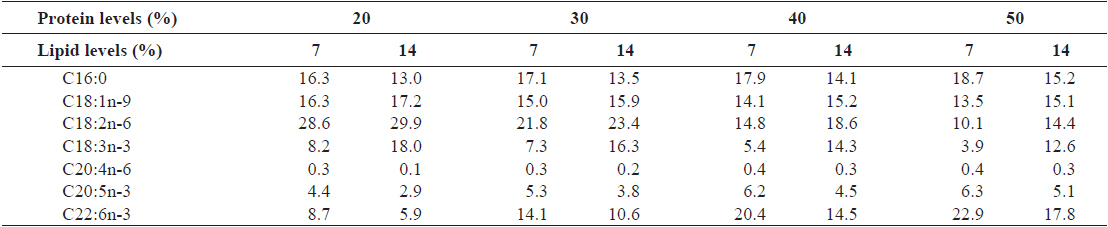
Major fatty acid composition (% of the total fatty acids) of the experimental diets
>
Experimental fish and feeding trial
Juvenile Israeli carp were obtained from the Inland Aquaculture Research Center, National Fisheries Research & Development Institute (Changwon, Korea). Fish were acclimated to experimental tank conditions and fed a standard commercial diet for 2 weeks prior to the start of the feeding trial. Fifty fish (initial mean weight, 1.3 ± 0.02 g) were then distributed randomly into 24 cylindrical plastic tanks (50 L water volume) for the feeding trial after being collectively weighed. Three replicate groups of fish were hand-fed to apparent satiation twice a day (09:00 and 17:00 for 6 days per week) for 9 weeks. Water temperature was maintained at 24.1 ± 0.82°C (mean ± SD), and the photoperiod was set to natural conditions during the feeding trail. Records were kept of daily feed consumption, mortalities, and feeding behavior in each tank.
>
Sample collection and analytical methods
At the end of the feeding trial, all fish in each tank were collectively weighed after being anesthetized with tricaine methanesulfonate (MS222, Sigma, St. Louis, MO, USA) at a concentration of 100 ppm after starvation for 24 h. Blood was drawn from the caudal vessel with 1 mL heparinized syringes from ten fish in each tank. The collected blood was centrifuged at 3,500 g for 10 min at 4°C, and the separated plasma was pooled and stored at -75°C for biochemical analysis. Total plasma protein, glucose, triglyceride, and cholesterol content were determined using an automatic analyzer (Toshiba-200FR, Tokyo, Japan). At the end of the feeding trials, fifteen fish from each tank were pooled and used for chemical composition analysis. Crude protein content was determined using the Auto Kjeldahl System (Buchi, Flawil, Switzerland), the crude lipid content was determined by the ether-extraction method, using a Soxhlet extractor (VELP Scientifica, Milano, Italy), the moisture content was determined with a dry oven (105°C for 6 h and the ash content was determined using a muffler furnace (600°C for 4 h). Lipid for fatty acid analyses was extracted by a mixture of chloroform and methanol (2:l v/v) according to the method of Folch et al. (1957), and fatty acid methyl esters were prepared by transesterification with 14% BF3-MeOH (Sigma, St. Louis, MO, USA). Fatty acid methyl esters were analyzed using a gas chromatography (PerkinElmer, Clarus 600, GC, USA) with a flame ionization detector, equipped with SPTM-2560 capillary column (100 m × 0.25 mm i. d. film thickness 0.20 μm; Supelco, Bellefonte, PA, USA). Injector and detector temperatures were both set to 240°C. The column temperature was programmed to increase from 140 to 240°C at a rate of 5°C min-1. Helium was used as the carrier gas. Fatty acids were identified by comparison with retention times of the standard fatty acid methyl esters (PUFA 37 component FAME Mix; Supelco).
Data were subjected to one-way and/or two-way analysis of variance (ANOVA) to test the effect of dietary protein and lipid levels on fish performance. When significant (
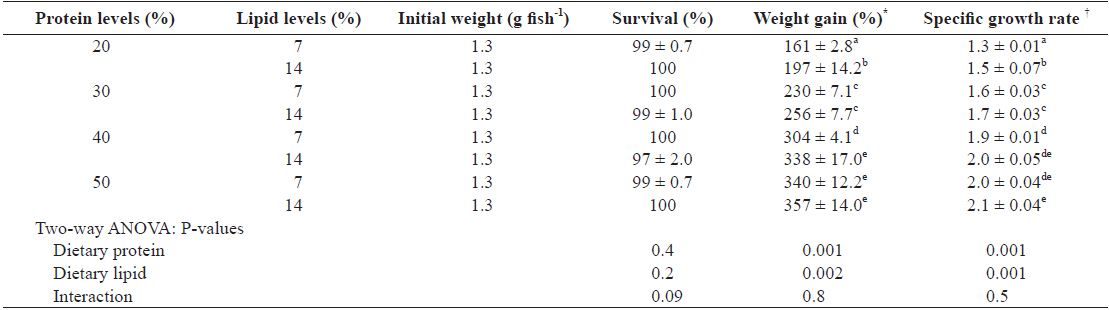
The growth performance data of juvenile Israeli carp fed the diets containing various protein and lipid levels for 9 weeks are presented in Table 3. Survival of each group was over than 97% and there was no significant difference among treatments (
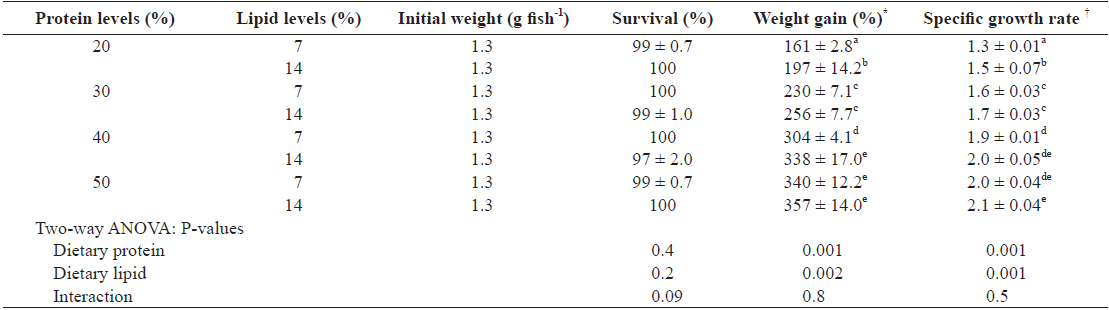
Growth performances of juvenile Israeli carp Cyprinus carpio fed the experimental diets containing various levels of protein and lipid
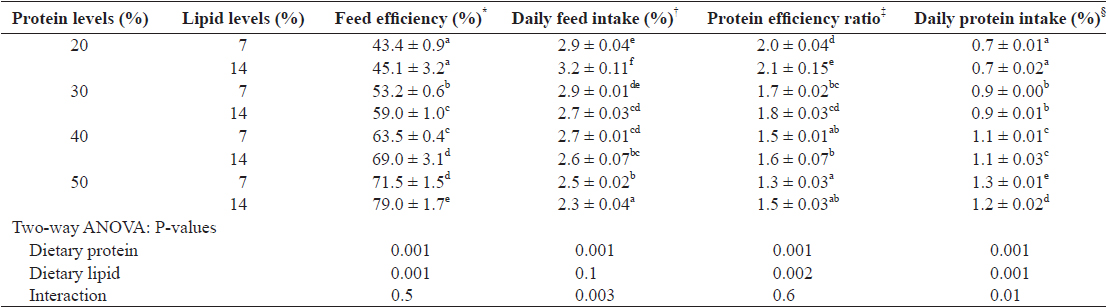
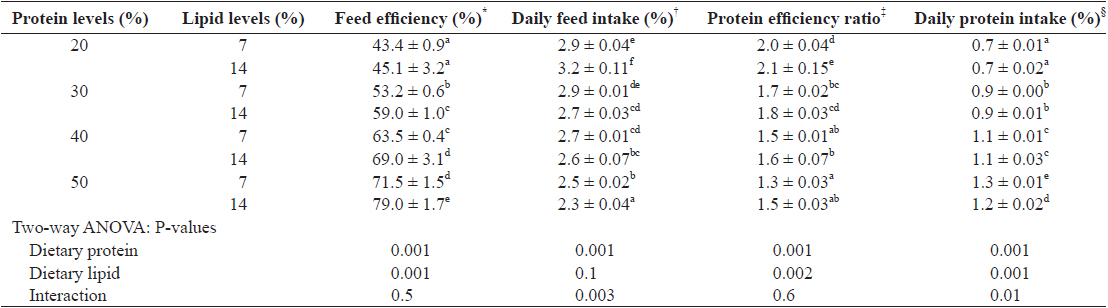
Feed utilization of juvenile Israeli carp Cyprinus carpio fed the experimental diets containing various levels of protein and lipid
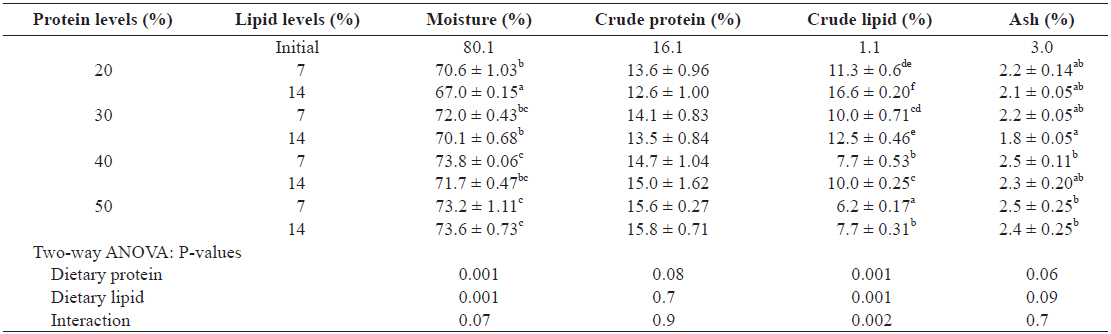
The proximate composition of whole body is given in Table 5. The moisture and crude lipid contents of whole bodies were significantly affected by dietary protein and lipid levels (
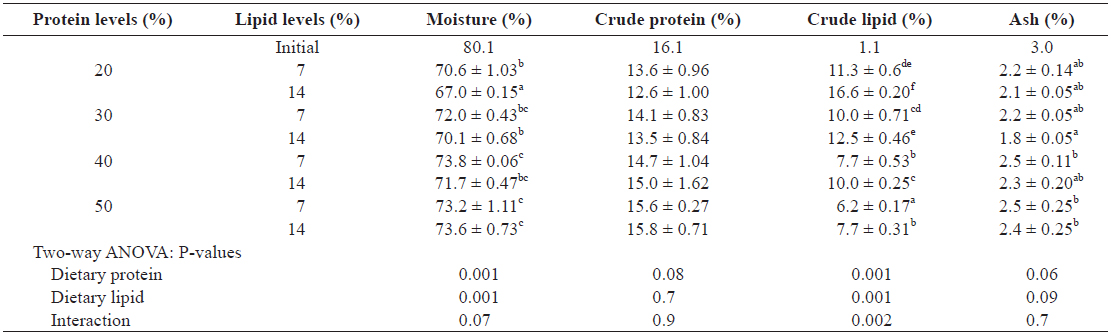
Proximate composition of the whole body of juvenile Israeli carp Cyprinus carpio fed the experimental diets containing various levels of protein and lipid
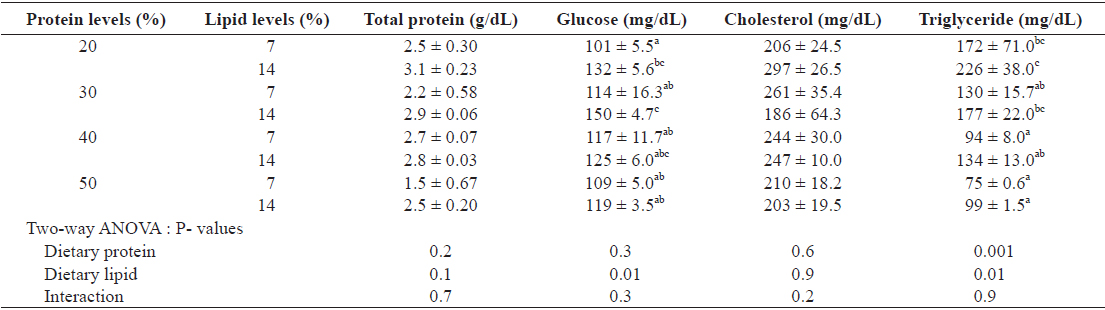
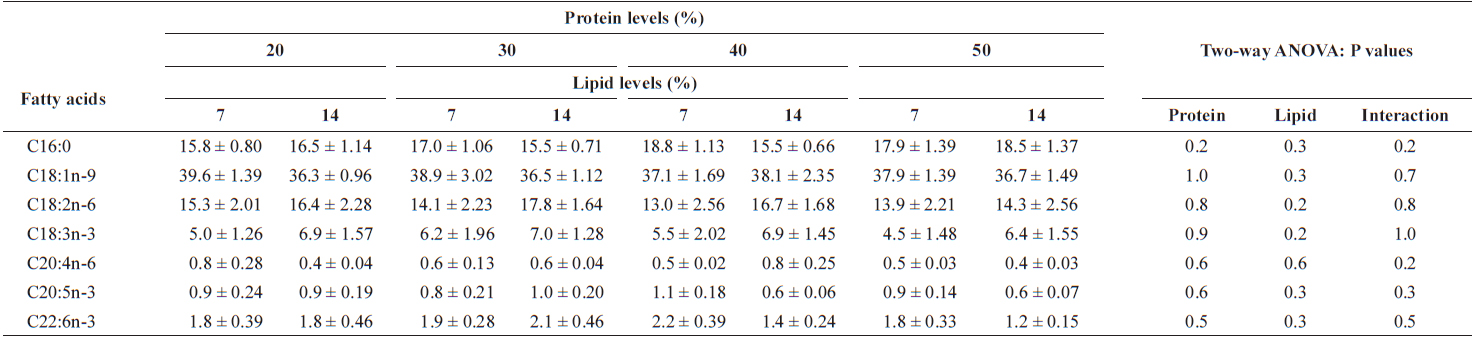
Hematological changes in the plasma of juvenile Israeli carp fed the diets containing various protein and lipid levels for 9 weeks are presented in Table 6. Total protein and cholesterol of the plasma of fish were not affected by either dietary protein or lipid levels. Glucose content of the plasma was affected by dietary lipid, but not by dietary protein. The increased dietary lipid resulted in higher glucose in plasma at 20 and 30% protein diet. Triglyceride of plasma of fish decreased with increasing dietary protein. Major fatty acid compositions (Table 7) of whole body in juvenile carp were not affected by dietary protein and lipid levels (
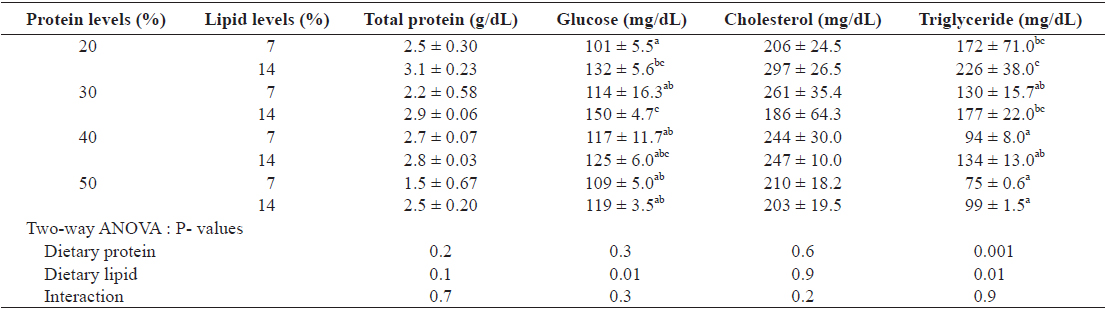
Hematological change of the plasma of juvenile Israeli carp Cyprinus carpio fed the experimental diets containing various levels of protein and lipid
Major fatty acid composition (% of the total fatty acids) of whole body of juvenile Israeli carp Cyprinus carpio fed the experimental diets containing various levels of protein and lipid
The results of this study showed that the 40% protein in the diet with 14% lipid is sufficient for growth of juvenile Israeli carp. This value is somewhat higher compared with the dietary protein requirement reported for some other cyprinids fish such as 30% for fingerling Indian major carp
A positive correlation was found between the dietary protein to energy (P/E) ratio and fish weight gain (r = 0.89;
Several studies have demonstrated that feed intake is regulated by the dietary available energy (Lee and Putnam, 1973; Jobling and Wandsvik, 1983), probably because the fish eat to satisfy their energy requirements. In this study, maximum daily feed intake was shown in fish fed 20% protein diet with 14% lipid, which corresponded to the lowest P/E ratio (11.4 mg kJ-1) and the highest level of non-protein energy/protein (64.1 kJ g-1) in the diet. This finding suggests that feed intake in juvenile Israeli carp is directly related to dietary protein and non-protein energy levels and ratios. It has been reported that feed intake is affected by dietary protein or lipid levels in channel catfish and grass carp (Page and Andrews, 1973; Du et al., 2005). Feed intake could be increased to compensate for the available essential amino acids or energy when inadequate amount of protein or energy in the diet is provided to fish.
The body moisture content in this study was positively correlated with dietary protein level (r = 0.74;
The results of this study indicate that an increase in the dietary lipid level has a protein-sparing effect, and a diet containing 40% protein with 14% lipid would be optimal for the growth and effective protein utilization of juvenile Israeli carp.