Disturbance is defined as “any relatively discrete event in time that disrupts ecosystem, community, or population structure and changes resource, substrate availability, or the physical environment” (White and Pickett 1985). Its impact depends upon the type, scale, intensity, and frequency. Two types of disturbances have been identified: 1) largescale, occurring over a wide area and causing great destruction; and 2) small-scale, in which branches or stems are broken and trees either snap or are uprooted (Kang and Choi 2000). Although the frequency of largescale disturbances is generally very low, some events, such as hurricanes or typhoons, can destroy an entire forest (Lorimer 1977, Bormann and Likens 1979, Canham and Loucks 1984, Boerner and Cho 1987). In contrast, small-scale disturbances occur more frequently, and are characterized by falling trees that can create a mosaic of various vegetation types and ages (Runkle 1982, Cho 1989, Cho and Boerner 1991).
A forest gap forms in the canopy when individual or multiple trees fall, or when large branches are broken on a single tree (Cho 1992). Strong winds are associated with approximately 97% of all gaps formed in mature forests within the southeastern United States and 80% of all new openings in Costa Rican rain forests (Barden 1981, Lawton and Putz 1988). Species with shallow root systems are especially vulnerable to high winds (Ruel et al. 2001). When large trees are uprooted, the forest floor is significantly altered because seeds lying within the soil become exposed and germinate, and the physical and chemical nature of the soil is changed by penetrating light (Greenberg and McNab 1998). In addition, soil gaps such as pits and mounds (Peterson and Pickett 1990) may lead to new microsites that can support the development of vegetation, ultimately changing the overall forest structure (Clinton and Baker 2000).
Odaesan National Park in Korea is a mosaic of various mature forests. The largest area is dominated by pure stands of
In October of 2006, forests within this park were subjected to strong winds and heavy rainfall. Throughout the eastern coast of Gangwon-do, including Sokcho, Gangneung, Donghae, and Pyeongchang (where Odaesan is situated), this storm inflicted damage to humans, buildings, fishing boats, and roads (Shin and Park 2006). A maximum gust of 63 m/s was measured at Sokcho, along with sustained winds of 31 m/s, and precipitation during this event totaled 232 mm. Throughout that month, Yangyang, Gangneung, and Donghae received record high precipitation, as well as winds in the typhoon category that exceeded 17 m/s. The nearest weather station, 22 km away in Daegwallyeong, indicated gusts of 22.5 m/s and a historically high 171.5 mm of rain during a single day (Korea Meteorological Administration 2008). Although weather conditions were not recorded at Odaesan during this storm, the largest old-growth tree, an
Fortunately, we have been monitoring several types of forests in the Odaesan National Park since 2005. The information of these reference forests provides a great opportunity to understand the structure and dynamics of the forests at the stand and species level (Kim 1996). Therefore, we investigated how the combination of strong winds and torrential rain, which are typical for Odaesan National Park, affected the development of forest gaps, influenced representative canopy tree species, and finally selected susceptible tree species to windthrow in that location.
Odaesan National Park represents a typical forested area populated by deciduous broadleaf tree species in central Korea. We examined the impact of wind damage on the structure of two types of forests there: 1) a mixture of the conifer
The
Our research area has a typical continental climate. The average annual temperature at the Daegwallyeong weather station is 6.4°C, and the average monthly temperatures range from –12.5°C (January) to 22.9°C (August). The average annual precipitation is 1,717.2 mm, with most rain falling in the summer (Korea Meteorological Administration 2008).
>
Measurement of forest gaps and identification of gap-maker tree species
The total number of forest gaps that formed following the storms on October 23rd and 24th of 2006 was determined for the mixed and
Gap-maker trees (DBH of ≥5 cm) were defined as those that contributed to the creation of gaps because they had either been uprooted or broken directly by the wind, or were struck by neighboring trees that had fell. The species of windthrown trees were identified, their DBHs were measured with the direction in which they fell, and they were labeled as either uprooted or snapped. The elevation, aspect, and slope of each gap were determined with an altimeter, compass, and clinometer, respectively. The soil depth at each site was measured by poking a 5-mm-diameter steel pipe into the ground six times.
The topographical characteristics of the canopy gaps in the two forest types are shown in Table 1. A total of 26 gaps were confirmed in the mixed forest. They appeared on the steep northeast-facing slope (average 37°) at an elevation of 990 to 1160 m, where the soil was very shallow, i.e., 13.8 cm. Because that area was a scree, it was difficult to determine the depth to which the trees had rooted. In contrast, the
[Table 1.] Topographical characteristics associated with canopy gaps formed in two forest types

Topographical characteristics associated with canopy gaps formed in two forest types
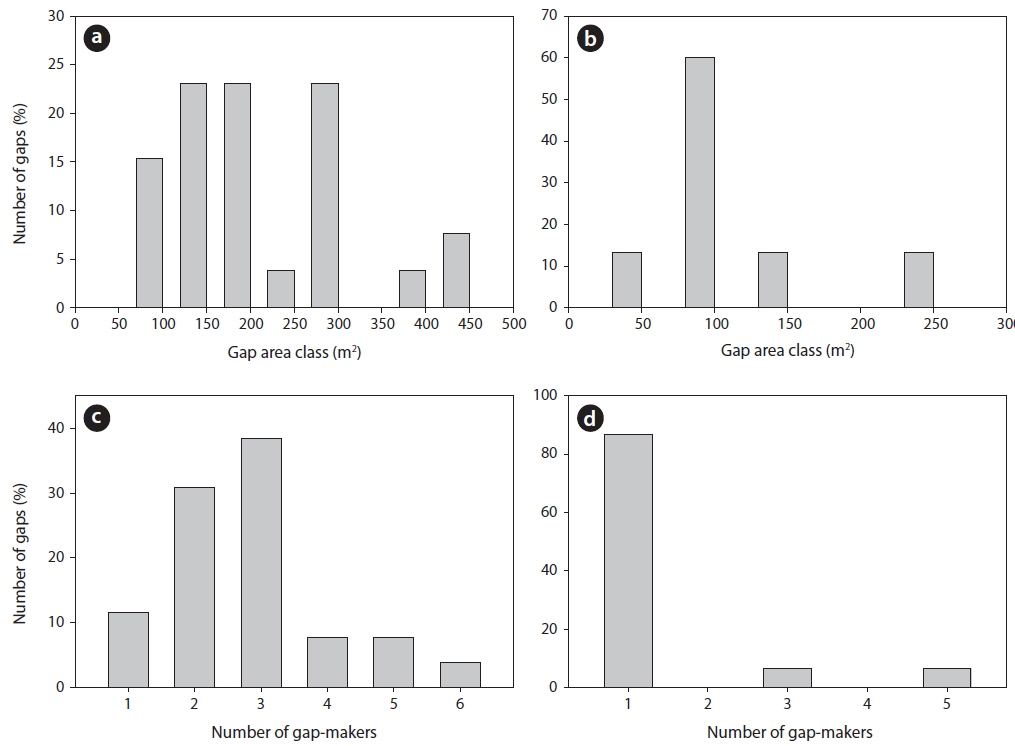
The 26 gaps in the mixed forest covered an average area of 205 m2 (75-442 m2), with 73% of them being 100 to 300 m2 in size (Fig. 2a). In contrast, the 15 gaps in the
A gap develops when a tree or trees in the upper canopy fall. Here, the average number of gap-maker trees was 2.8 and 1.4 in the mixed forest and
>
Characteristics of gap-maker species
The 26 gaps in the mixed forest resulted from the fall of 73 trees. Among these, 46 (63%) were downed by wind, while the remaining 27 were struck by neighboring trees as they fell (Table 2). The average diameter of trees directly damaged by wind was 41.5 cm compared with 25.4 cm for those that were indirectly damaged by falling trees. Most of the windthrown trees (73.9%) had thick trunks and, thus were uprooted, while those hit by neighboring trees (67.8%) had narrower trunks and, instead, were primarily snapped (Fig. 3). The 15 gaps in the
[Table 2.] Causes of tree-fall and death of trees in two forest types

Causes of tree-fall and death of trees in two forest types
In the mixed forest, the gap-maker trees were members of 18 species. The largest proportion (27.3%) was the
Of the gap-maker trees in the mixed forest, 60% of
To investigate whether particular tree species are more vulnerable to wind damage, we compared species composition between our designated windthrown forests and nearby undamaged reference plots. The relative density of

Species composition of gap-maker trees directly damaged by wind in mixed forest compared with population in nearby undisturbed reference forest (DBH ≥5 cm)
In the reference plots for the

Species composition of gap-maker trees directly damaged by wind in Quercus forest compared with population in nearby undisturbed reference forest (DBH ≥5 cm)
The types of vegetation that become established in particular regions depend upon numerous factors, such as parental rock materials or topography, as well as the opportunities that make species distribution amenable. Even under similar environmental conditions, forests in a certain area can comprise of a mosaic of stands with different successional stages and vegetation types. This results from not only natural or artificial methods of regeneration, but also from large- or small-scale disturbances, the frequencies of those events, and the ways in which sites recover from such challenges.
The forests within Odaesan National Park are heterogeneous, but trees of at least 100 years old are widely distributed here, suggesting that large-scale harvests or forest fires have not taken place for the time being. This assumption is based on signs of selective logging for certain species and scattered clumps that have regenerated via sprouting. Fires might have been a threat in the past; however, at present it probably is not true in that park because the most common species on that mountain have proven to be highly resistant (Choung et al. 2004). Furthermore, these forests have been protected by rigorous forest management policies since the 1970s (Odaesan National Park Service 2008). Therefore, natural mortality and occasional high winds are the major disturbances that now form canopy gaps. The development of such gaps is largely a function of forest type, the prevailing disturbance regime, and geographical location (Foster 1988). Daegwallyeong, an area that borders this park, is known in Korea for having very strong winds (average speed of 5.8 m/ s), a velocity that is second to Jeju Island’s 8.9 m/s (Kim 2005). Because of that, the region is now the site of a wind farm. The high frequency and strength of these winds mean they have the potential to also cause major disturbances. Odaesan National Park is topographically vulnerable to wind damage because of its high elevation, steep slopes, and shallow soils on numerous scree sites. Moreover, when strong winds are accompanied by heavy rainfall, as it happened in October of 2006, the saturated soil becomes unstable, causing tall trees to fall, as they act as masts (Clinton and Baker 2000).
In both of our forest types, canopy gaps tended to form on the slopes, suggesting that such locations are more affected by strong winds than mountain ridges or flat areas (Kim 1996). The direction in which gap-makers fall is influenced by topography and the type and extent of the disturbance (Cho 1992). In particular, because trees on flat land are considerably affected by wind direction, they tend to fall in a certain direction that makes it possible to estimate the severity of a particular disturbance (Falinski 1978). When no specific disturbance has occurred, however, the fall direction is largely influenced by topography. Cho (1992) showed that in the neighboring area of Gwangneung Natural Forest at Soribong, most of the dead trees fell along the slope aspect in the absence of any disturbance. However, we found that trees growing on northeast-facing slopes in our mixed and
Gap sizes in the mixed forest averaged 205.0 m2 (75-442 m2). This is approximately similar to the 246 m2 reported from a study of deciduous broadleaf trees in a mature forest at Gwangneung (Cho 1992), where the vegetation structure was almost the same as what we recorded in our current survey. In contrast, our
In the mixed forest, the majority of windthrown gapmakers belonged to
The undamaged sites within the
A singularly strong wind accompanied by a blast of rainfall selectively uprooted trees of
Such small-scale disturbances alter site conditions and change the levels of resources, including light and moisture available for the remaining trees to grow. These scenarios also lead to the rapid regeneration of the existing canopy species or saplings and seedlings that are found in the understory layer according to the gap size.
Overall, these findings suggest that