The generation of transgenic organisms expressing living fluorescent protein reporters under the regulation of specific promoters has become an established technique in a variety of research fields, particularly in developmental and cellular biology (Gong et al., 2001; Chudakov et al., 2010). In addition, certain aquarium fish that have acquired vivid and faithful fluorescent colors in their external phenotypes through transgenesis have been developed as novel ornamental animals (Gong et al., 2003; Zeng et al., 2005; Stewart, 2006). Several fluorescent transgenic zebrafish Danio rerio and Japanese medaka Oryzias latipes strains have already been launched in the aquarium markets in the United States (Bratspies, 2004; http://www.glofish.com) and Taiwan (http://www.azoo.com.tw), respectively. The successes achieved in these pioneering works have encouraged the extension of fluorescent transgenic techniques to other fish species (Pan et al., 2008; Cho et al., 2011; Cho et al., 2013).
The previous studies described above have shown that transgenic expression, particularly in the intensity of fluorescent signals, could be largely governed by the strength of the promoter used, although genotype-dependent differences with a given transgene construct should also be considered (Wan et al., 2002; Gong et al., 2003; Zeng et al., 2005; Burket et al., 2008; Ge et al., 2012). However, in addition to promoter dependency, our literature-based survey indicated that the expression properties of transgenic fish (i.e., the visibility and brightness of fluorescent colors in their external phenotypes) could also be affected substantially by the different fluorescent protein genes used, even if they are under the control of the same kind of promoter (Finley et al., 2001; Wan et al., 2002). Each fluorescent protein (e.g., green, red, cyan, or yellow) has its own structural characteristics, often including modifications targeted to its wild-type progenitor. Unique photophysical properties such as maturation speed, perceived brightness, extinction coefficient, quantum yield, and/or photostability (Chalfie and Kain, 2005; Shaner et al., 2005) should be considered as important parameters in the aquarium display of fluorescent transgenic fish. To date, the fluorescent phenotypes of transgenic fish expressing green or red fluorescent protein (GFP or RFP, respectively) have been well documented in terms of ornamental applications (Hamada et al., 1998; Chou et al., 2001; Gong et al., 2001; Cho et al., 2011). However, the transgenic phenotypes developed using other fluorescent protein genes have been less extensively characterized in transgenic fish strains, with the exception of a few studies (Finley et al., 2001; Gong et al., 2003; Kinoshita, 2004; Ge et al., 2012). Moreover, the recent explosion in the diversity of newly available fluorescent proteins with improved photostability, folding efficiency, and brightness could offer a novel opportunity to develop more qualified ornamental transgenic fish strains with better visibility of their fluorescent colors.
AmCyan1 (excitation at 458 nm and emission at 489 nm) is the commercial brand name of the mutant version of the cyan fluorescent protein (CFP) amFP486, isolated from the non-bioluminescent Anthozoa species Anemonia majano (Matz et al., 1999). This coral reef organism-originating CFP (amFP486) exhibits a large structural difference from earlier CFP variants created from the jellyfish Aequorea victoria GFP, and several targeted modifications are known to have improved the brightness, solubility, and photostability compared to earlier versions of CFP (Clontech Laboratories Inc., Mountain View, CA, USA). As a result, this fluorescent protein (AmCyan1) is considered to be a valuable fluorescent reporter protein in various experiments that require multicolor detection. As a genetically distinct alternative reporter due to its unique spectra, the utility of AmCyan1 has been applied to protein localization and/or transgenic studies in plants (Wenck et al., 2003; Tang et al., 2006), mammals (Kawamata and Ochiya, 2010), insects (Sarkar et al., 2006), algae (Mikami et al., 2011), and zebrafish (Bertrand et al., 2008; Smith et al., 2010). However, most previous studies on cyan-transgenic fishes have focused on the functional utility of this CFP reporter in the developmental monitoring of a specific protein and/or cell lineage based on epifluorescence microscopy (see above references). However, in contrast to the abundant information on fluorescence microscopy data, the external phenotypes exhibited by the cfp-transgenic organisms during their life spans have been less characterized, although they are key chacracterists that should be considered for the ornamental application of transgenic fish. Furthermore, because the fluorescent appearances of certain fluorescent proteins perceived by human eyes may differ from those visualized with the specific filter sets implemented in fluorescence microscopy, the phenotypic attributes of the fluorescent colors achieved in transgenic fish should be empirically evaluated under aquarium conditions. However, currently, the ornamental evaluation of CFP in transgenic fish is still limited; only a transgenic zebrafish carrying an earlier version of the enhanced cfp (ecfp) transgene has been briefly described (Gong et al., 2001).
Marine medaka Oryzias dancena, a truly euryhaline teleost, has many advantageous merits as a candidate laboratory organism for various biological and ecotoxicological studies (Cho et al., 2010; Chen et al., 2011). Recently, we showed that the transgenesis of an RFP reporter driven by either the ubiquitous β-actin (actb) promoter or the fast-skeletal muscle specific myosin light chain 2 (mlc2f) promoter could faithfully visualize a red fluorescent phenotype in the transgenic marine medaka, in which the intensive fluorescent color could be easily presented due to the transparent body color of this species (Cho et al., 2011, 2013). Data from our previous studies strongly suggested that the fluorescent transgenic marine medaka held promising potential as novel ornamental fish strain to be displayed in both marine and freshwater aquaria. In line with our long-term goal of developing various fluorescent versions of transgenic marine medaka for novel ornamental varieties in future aquarium trade, the objective of this study was to evaluate the ornamental characteristics of a novel variety of stable transgenic marine medaka germlines harboring the AmCyan1 transgene driven by the marine medaka mlc2f promoter under different light-irradiation conditions.
The podmlc2AmCFP transgene was constructed by inserting a 2.95-kb O. dancena mlc2f promoter (Lee et al., 2013) in front of the ATG initiation codon of the cfp gene in the pAmCyan1-C1 plasmid (Clontech Laboratories Inc.) using the KpnI and AgeI sites. The resultant plasmid, podmlc2AmCFP (7.12 kb), was linearized by digestion with ClaI (New England BioLabs, Ipswich, MA, USA), gel-purified, and resuspended in an injection buffer (10 mM Tris–HCl, 0.1 mM EDTA, pH 8.0). One-celled embryos were microinjected with the ClaI-linearized podmlc2AmCFP construct (50 μg/mL) and transferred to an incubator at 26 ± 1°C until hatched. The salinity of the incubation water was adjusted to 5 ppt using synthetic sea salt (Kent Marine Aquarium Products, Acworth, GA, USA). During embryonic development, CFP-positive embryos were identified using fluorescence microscopy at the prehatching stage (~10 days postfertilization; dpf). The cyan fluorescent signal was analyzed with an AZ100 fluorescence microscope equipped with the NISElements BR image analysis software (Nikon Corporation Instruments Company, Tokyo, Japan). CFP expression was observed with the Nikon CFP filter (excitation filter wavelengths = 426–446 nm; dichromatic mirror wavelength cuton = 455 nm; barrier filter wavelengths: 460–500 nm), and the image was photographed using a digital camera (Nikon digital sight DS-Ri1) mounted on the AZ100 microscope. After hatching, the selected CFP-positive larvae were further grown until sexual maturity (~9 months). At sexual maturity, the presumed transgenic founders with CFP signals were subjected to the test for germline transmission to F1 offspring. Each adult CFP-positive transgenic female founder was crossed with a wild-type, non-transgenic marine male fish in a 1:1 mating manner. Transgenic male founders were crossed with four non-transgenic females. At least 100 F1 embryos from each mating were examined for CFP signals using fluorescence microscopy as described above. The F1 CFP-transgenic individuals from each transgenic line were mated with non-transgenic individuals to examine the transmission of the fluorescent transgene to the F2 generation following the Mendelian single gene inheritance pattern. The mating and CFP-typing for propagating subsequent generations were performed according to the procedures described above.
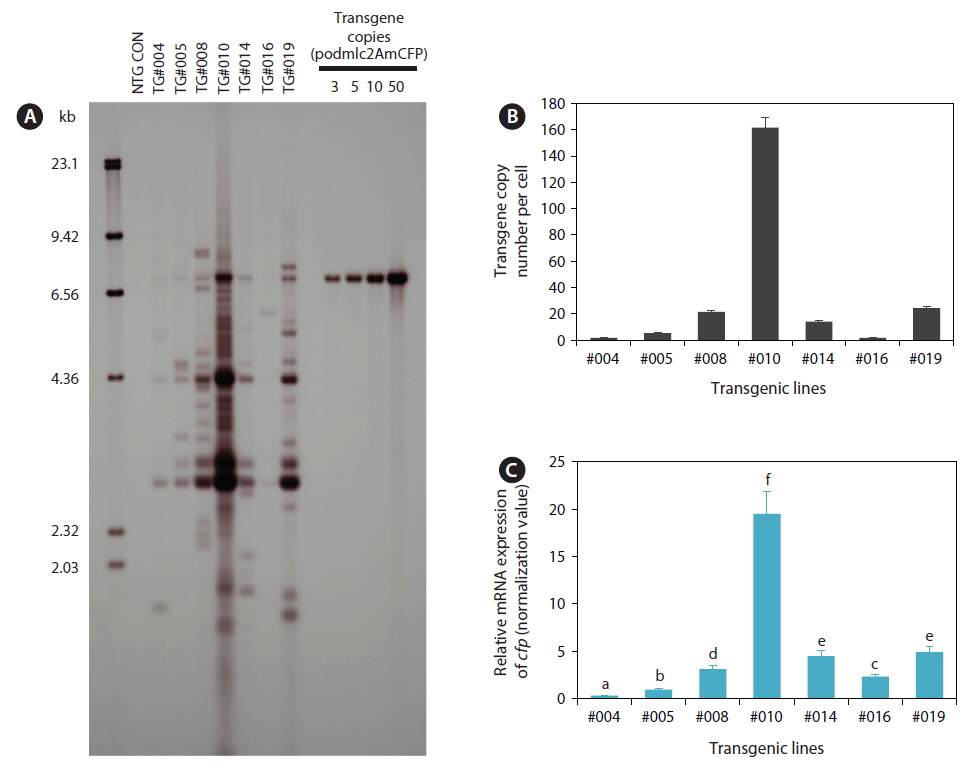
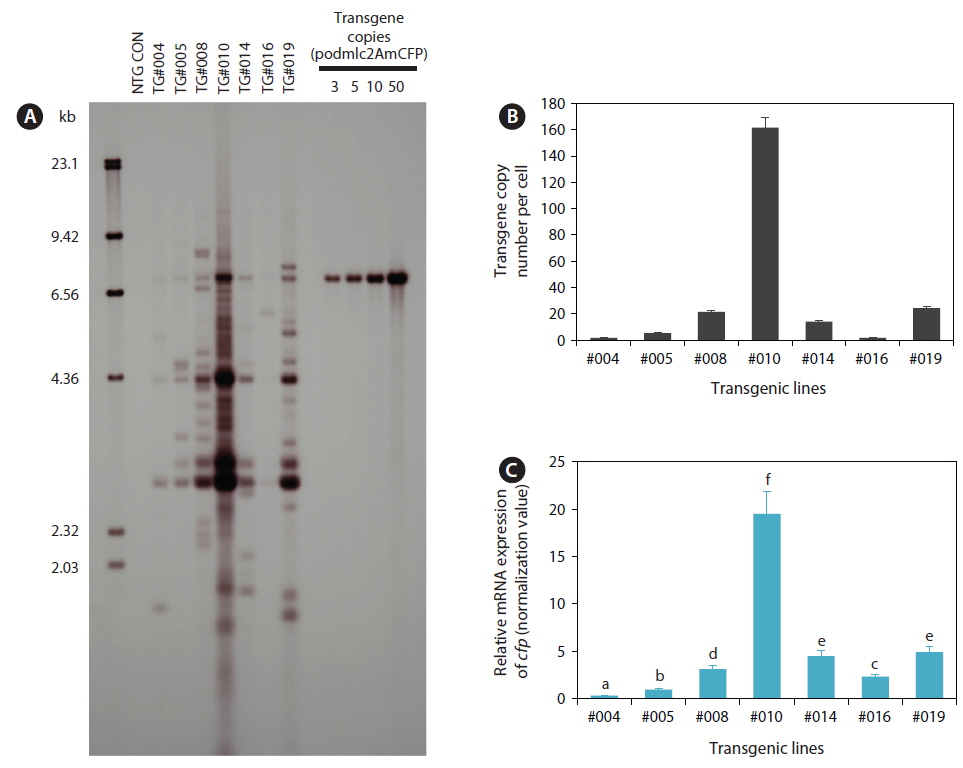
To confirm the presence of podmlc2AmCFP in the F1 fluorescent transgenic progeny, polymerase chain reaction (PCR) amplification was performed using the primer pair T-ODMLC2 FW (complementary to the mlc2f promoter; 5´-ACCATCACTTGATGGTCGACCA-3´) and ODAmCFP 1R (complementary to the cfp structural gene: 5´-TGCCGTACATGAACACGGTG-3´). PCR-positive, fluorescent transgenic individuals (F1) belonging to each transgenic line were subjected to genomic Southern blot hybridization analysis to confirm integration of the transgene into the host chromosome. Genomic DNA prepared from caudal fins using the conventional proteinase K/sodium dodecyl sulfate (SDS) method was digested with HindIII, and 5 μg of the digested DNA was separated on a 0.8% agarose gel, transferred to a nylon membrane, and hybridized with a digoxygenin (DIG)-11-dUTP labeled cfp probe (527 bp). All the procedures for Southern blot hybridization were performed according to the instructions provided in the DIG nonradioactive DNA labeling and detection kit (Roche Applied Science, Mannheim, Germany). The transgene copy number per cell was estimated with quantitative real-time PCR (qPCR). Spectrophotometrically measured genomic DNA(10 ng) was subjected to a thermal cycling reaction in a reaction mixture that included 2× iQ™ SYBR® Green Supermix (Bio-Rad, Hercules, CA, USA) and a primer pair (qAmCFP 1F: 5´-CTACAGATGCCAGTTCCACA-3´ and qAmCFP 1R: 5´-GAGATCTGAGTCCGGAGAAG-3´) designed to amplify a 182-bp internal fragment of the cfp gene. Thermal cycling was performed on an iCycler® Real-Time Optical Module (Bio-Rad) with the default settings. Before the qPCR assay of the transgene copy numbers, each genomic DNA sample was confirmed to have a uniform cycle threshold (Ct) number (Ct value variation <0.3) in the control amplification of the endogenous β-actin gene actb (GenBank accession number HM347346; data not shown). Based on the standard curves prepared with 4-log dilutions (1‒1000 copies) of positive plasmids, the transgene copy number in each DNA sample was determined. PCR efficiency ≥90% was confirmed for each amplification reaction. Triplicate assays were performed for each DNA sample from individual fish (three individuals per transgenic line) in an independent fashion.
To examine any positive relationships between the fluorescence intensity of transgenic CFP signals and expression levels of cfp mRNAs in the skeletal muscles of transgenic fish (F2 generations), reverse transcription qPCR (qRT-PCR) assays were performed. Three transgenic individuals (8 or 9 months old) from each selected transgenic line were photographed with fluorescence microscopy, and then the skeletal muscle of the each individual was subjected to total RNA isolation for qRT-PCR. The total RNA sample was purified using the RNeasy Plus Mini Kit (Qiagen, Hilden, Germany), and a 2-μg aliquot was reverse-transcribed into cDNA using the Omniscript Reverse Transcription Kit (Qiagen) and oligo(dT)20 primers. During the RT reaction, an O. dancena 18S rRNA gene reverse primer (OD18S RV; GenBank accession number HM347347) was also included in the RT reaction at a final concentration of 0.01 μM to normalize the amount of input total RNA across samples (Cho et al., 2011). The synthesized cDNA sample was diluted twofold with sterile water, and 2 μL of the diluted cDNA was used as the template for qPCR. The PCR primers for the cfp transgene (qAmCFP 1F and qAmCFP 1R) were the same as those used for the transgene copy number assay above, while the normalization control 188-bp internal fragment of the 18S rRNA gene was amplified using the primers qOD18S 1F (5´-AAGCTCGTAGTTGGATCTCG-3´) and qOD18S 1R (5´-CCTAGCTGCGGTATTCAGGC-3´). The relative expression level of the cfp transcripts in each transgenic line was normalized to the level of 18S rRNA using the following formula: relative expression = [(1 + Ecfp)Ct cfp]−1/[(1 + E18SrRNA)Ct18S rRNA]−1, where E is the PCR efficiency (E = 10−1/slope − 1) and Ct is the cycle threshold number. Triplicate independent assays per cDNA sample were performed. Differences among the samples were assessed with ANOVA, followed by Duncan’s multiple range test at P = 0.05.
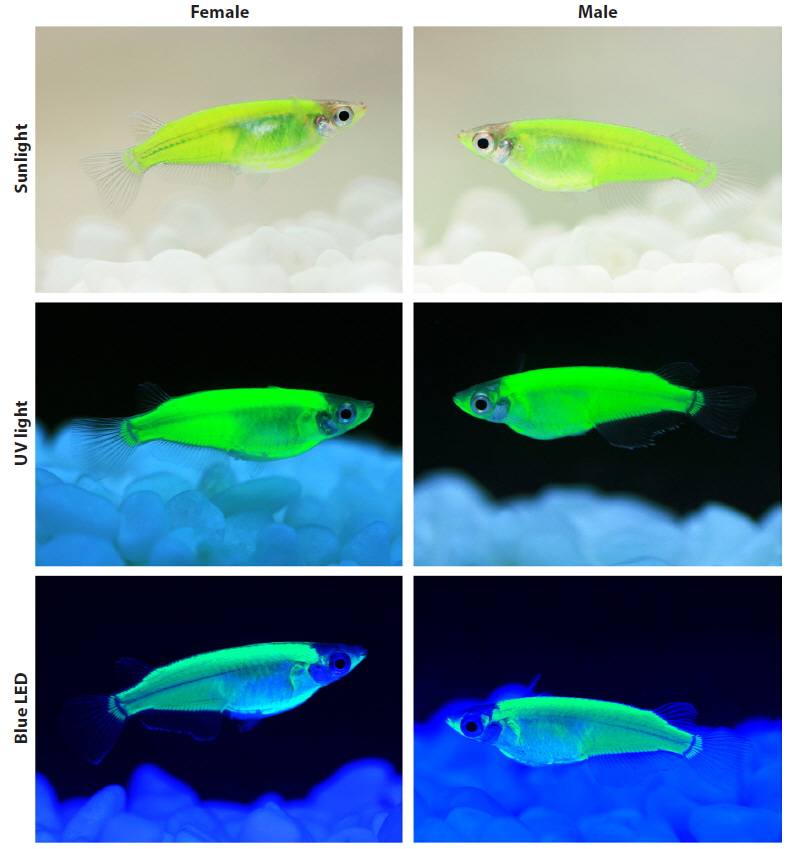
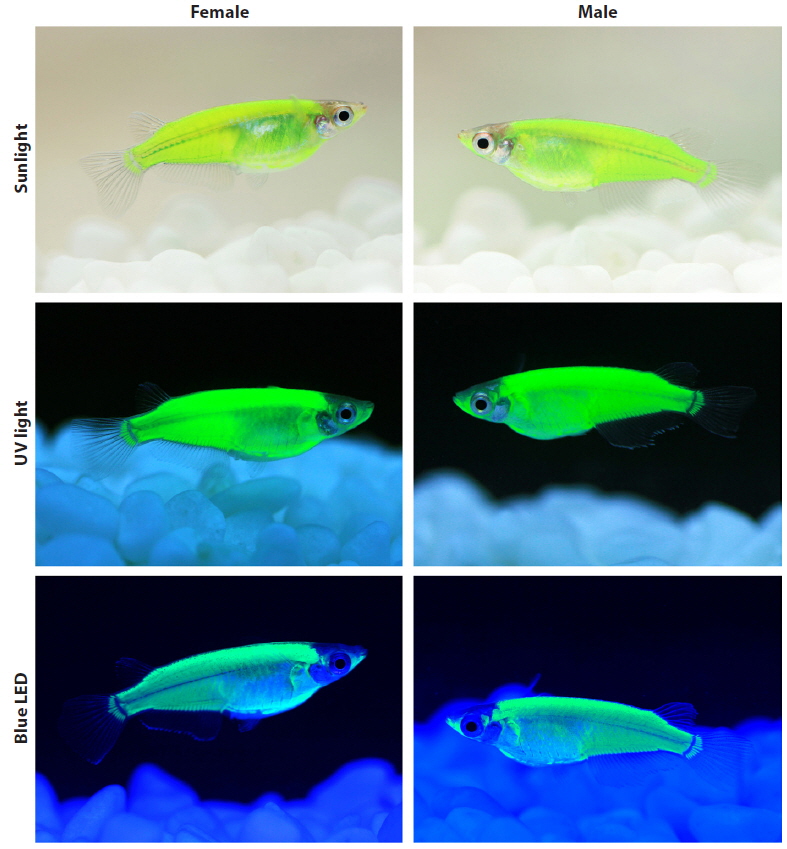
After growing to the adult stage (~9 months of age), the external phenotypes of the CFP-transgenics were examined under different light-illumination conditions, including daylight (sunlight), normal white fluorescent room light (14 W: ERI-Su1125-6007; ERI, Beijing, China), ultraviolet (UV) light (black light lamp: 352 nm, 15 W; Sankyo Denki Co., Ltd., Tokyo, Japan), aquarium blue light (FHF 14STEX-D blue lamp, 450–495 nm, 14 W; Leedarson Lighting Co., Ltd., Fujian, China), and multiple colors of light-emitting diode (LED) lights. The adjustable LED illumination control system (LED-Lighting ZigBeeControl Program) was designated by the LED-Marine Convergence Technology R&BD Center (Pukyong National University, Busan, Republic of Korea) to emit blue (454 nm), green (517 nm), and red (628 nm) light in an independent or mixed fashion. The external CFP phenotype was photographed using a macrolens (EF 100 mm; 1:2.8L USM) connected to an EOD 5D Mark II digital camera (Canon, Tokyo, Japan).
Of 1650 embryos microinjected in eight trials, 697 fish reached sexual maturity, and 62 fish possessed CFP signals in their external bodies as determined by fluorescence microscopy. All of the CFP-positive founder fish exhibited a mosaic distribution of the transgenic CFP signals in their bodies, suggesting that their transgenic status would be mosaic, as commonly observed in founder generations of transgenic fish generated by microinjection (Nam et al., 1999; Figueiredo et al., 2007; Hartmann and Englert, 2012). Based on the intensity and expression areas of CFP signals observed in their external bodies, 19 transgenic founders (8 females and 11 males) were selected for the testing of germline transmission to F1 offspring. Of the 19 founders tested, seven individuals (six males and one female) were demonstrated to pass the fluorescent transgene to their F1 progeny. All founders transmitted their transgenes to offspring with a germline transmission frequency below 50%, indicating that they were mosaic for their germ cells as well. Despite severe mosaicism in the founder generation, the transgenic hemizygous status was stabilized from the F1 generation in each transgenic line. All of the tested F1 fish, irrespective of their transgenic lines, passed the fluorescent transgene to their F2 progeny at a frequency close to 50% (now up to F4). PCR typing of a random sample of CFP-positive or CFP-negative larvae also showed that the CFP signal clearly coincided with the presence of the podmlc2AmCFP transgene (data not shown). Obligate tetramerization of overexpressed fluorescent proteins has been known to potentially exhibit apparent toxicity to animal embryos (murine embryos in particular), as evidenced by DsRed, a popular RFP isolated from Discosoma sp. (Long et al., 2005; Yang et al., 2009). A certain transgenic fish line overexpressing the RFP protein (DsRed2) ubiquitously due to high transgene integration reportedly suffered from physiological abnormalities and reproductive impairment (Cho et al., 2011). However, data on the germline transmission in this study undoubtedly indicate that expression of the AmCyan1 CFP protein exclusively in skeletal muscles had no adverse effect on the normal development and viability of this species. As yet, no significant differences have been observed between CFP-positive and non-transgenic individuals for other physiological attributes, such as growth and reproduction.
Genomic Southern blot hybridization analysis with seven selected transgenic lines showed that multiple transgene copies were integrated into the host genomes in all the transgenic lines examined. Each transgenic line was represented by its own unique hybridization pattern in the HindIII blot, and the copy numbers of integrants varied greatly among the transgenic lines (Fig. 1A), indicating the random integration of microinjected transgene constructs, similar to previous reports on other transgenic fish strains (Tewari et al., 1992; Chou et al., 2001; Cho et al., 2011). Some of the transgenic lines, particularly lines TG#008, TG#010, and TG#019, exhibited signs of possible concatemerization of multiple transgene copies prior to integration, which reportedly is a typical phenomenon observed in microinjection-based gene transfer in fishes (Alam et al., 1996; Nam et al., 1999; Grabher and Wittbrodt, 2007). The results of transgene copy numbers per cell determined based on the qPCR assay also agreed well with those of the Southern blot analysis. Average transgene copy numbers were 1.76 (TG#004), 5.44 (TG#005), 21.60 (TG#008), 161.60 (TG#010), 14.23 (TG#014), 1.66 (TG#016), and 24.40 (TG#019) (Fig. 1B). Considering the germline transmission frequency of the fluorescent transgene from F1 to F2, which followed the Mendelian single gene inheritance pattern in all transgenic lines, these multiple transgene copies might integrate into a single chromosomal site or very closely neighboring regions in a chromosome (Kinoshita, 2004; Cho et al., 2011).
Based on the qRT-PCR assay of transgenic mRNA in the skeletal muscles, transgenic lines with higher transgene copy numbers tended to exhibit greater expression levels of cfp transcripts than those with lower transgene copy numbers (Fig. 1C). However, the relationship between transgene copy numbers and mRNA expression levels was not directly proportional, as the efficiency of transgenic mRNA expression per transgene copy was lower in some high-copy number transgenic lines (e.g., TG#010 and TG#019), although they did display the greatest amount of cfp transcripts in an absolute manner. Transgenic loci containing long, repetitive concatemers are often prone to rearrangement or modification, leading to unwanted transgene silencing in transgenic animals (Davis and MacDonald, 1988; Wolffe, 1997; Geurts et al., 2006). However in our transgenic lines, the fluorescent phenotype acquired in the F1 generation has been shown to be stable, currently up to F3 or F4, with no notable sign of loss or reduced intensity of the transgenic CFP phenotype. Further exploration of the transgene loci, including the organization of transgene copies, integration sites, and neighboring sequences, will be necessary to obtain deeper insight into the copy number-dependent or -independent expression of the transgene in these transgenic lines.
During development, the onset of CFP signals was first detectable from the completion of somitogenesis (~3.5 dpf) to the formation of the tubular heart, and the expression level gradually increased as development proceeded. Overall, the temporal patterns of CFP expression observed in transgenic marine medaka embryos were in accordance with previous findings on the endogenous expression of mlc2f and transgenic expression of other fluorescent genes driven by the mlc2f promoter (Xu et al., 1999; Gong et al., 2003; Ju et al., 2003; Cho et al., 2013). After reaching adulthood, transgenic fish belonging to each transgenic line were easily distinguishable from non-transgenic individuals with the unaided eye under normal daylight conditions owing to the authentic fluorescent signals achieved in their external bodies. The distribution pattern of the transgenic CFP signal was the same for all transgenic lines examined, although the intensity of the CFP signal varied among the lines. Transgenic expression was highly dominant in dorsal parts, the caudal trunk, and lower jaw-borne muscles, and moderate in the belly region. The fin buds, especially the region connected to the hypural bone, were also clearly CFP-positive (see Fig. 2). In an external view, however, the CFP signal was not detected in the cranial region, eyes, or fins (Xu et al., 1999; Ju et al., 2003; Cho et al., 2013).
The fluorescent colors were affected considerably by the light-irradiation sources (Fig. 2). Under normal daylight (i.e., sunlight) conditions, the CFP-transgenic fish exhibited only a yellowish-green phenotype. A similar greenish color was observable under conventional white fluorescent room lighting. The green glow was highly intensified when the transgenic fish were irradiated under UV light (352 nm), and a very bright green fluorescence was visualized. Unlike UV illumination, which resulted only in green fluorescence, the blue LED light (454 nm) caused the CFP-transgenic fish to appear bluer. Then under the blue LED lighting, the CFP-transgenic fish exhibited more cyan-like appearances as a bluish-green phenotype, although the true cyan fluorescent glow observed under fluorescence microscopy was not achieved. As expected, the CFP-transgenic fish responded to neither green (510 nm) nor red (650 nm) LED light alone. Irradiation with mixed LED lights (e.g., blue together with green or red) only altered the fluorescent colors of the transgenic fish slightly (photographs not shown). No differences in fluorescent external phenotypes were observed between male and female transgenics.
In this study, the AmCyan1-transgenic marine medaka did not present a true cyan fluorescence-emitting color under the aquarium light conditions. Instead, the transgenics were generally green-glowing, although they presented a more blueshifted phenotype in response to blue LED light. However, even under blue LED light conditions, the phenotype color of the transgenics did not appear as true cyan to the human eye. A similar observation was reported for the cestrum virus promoter-driven AmCyan1-transgenic rice (Oryza sativa) callus, which exhibited a slight green–yellow-glowing color without the use of fluorescence optics (Wenck et al., 2003). Some mechanisms of spectrophotometric tuning have recently been reported, although most postmortem studies have based their results on epifluorescence microscopy, and the effects on the external phenotype of a whole organism in vivo remain unknown. Although, we have yet to clarify the mechanism responsible for the generally green fluorescent phenotype of AmCyan1-transgenics, one possible explanation is the structural similarity of the chromophore-forming tripeptides (X–Y–G; X = hypervariable, Y = Tyr and G = Gly) between AmCyan1 and GFPs, even though AmCyan1 has a quite different overall polypeptide structure, including two key residues (Ala-165 and His-199) that make an important contribution to the blueshift (Henderson and Remington, 2005). The importance of the chromophore structure to the optical characteristics of fluorescent proteins, rather than the amino acid environments over the entire polypeptide, has been previously reported (Kiseleva et al., 2008). Consequently, similar chromophore structures may result in similar fluorescent appearances when perceived by human eyes, even though the specific filter sets used in fluorescence microscopy are able to distinguish the colors. Meanwhile, although AmCyan1 was not designated to have a photo-switching function, the mutant CFP generated from the mutagenesis of Aequorea coerulescens GFP has been reported to be highly capable of photoconversion from a cyan to green fluorescent form in response to 405-nm UV light irradiation (so termed photo-switchable CFP; PS-CFP) (Chudakov et al., 2004, 2010).
In sum, our results suggest that the ornamental evaluation of fluorescent color acquired by transgenic fish should be empirically performed independently of optic data generated with fluorescence microscopy, and that fine-tuning of light-irradiation apparatuses might be necessary to optimize the visibility of ornamental transgenic fish. Further studies should be performed to pinpoint the effects of specific light wavelengths on transgenic fluorescent phenotypes under aquarium display conditions.