Balanced nutrients and enough calories in aquafeed are essential for growth and maintenance of farmed fish. Fish are known to utilize protein preferentially to lipid or carbohydrate as an energy source (Walton and Cowey, 1982). Dietary protein is the most important factor affecting both growth performance of fish and feed cost (Lovell, 1989). Accurate information on the protein requirements of fish is crucial for any aquaculture initiative owing to the high cost of protein ingredients which are usually required at high levels by most fishes (NRC, 1993).
Far eastern catfish is one of the most important commercial freshwater fish species, because this species has high consumer demand in Korea. This species is distributed in Korea, Japan, China and Taiwan (Chyoung, 1996). Aquaculture production of far eastern catfish has steadily increased during the last decade, and its production in 2012 reached 3,676 metric tons, which is second only to eel (4,259 metric tons) among freshwater fish aquaculture in Korea (Statistics Korea, 2013).
Recently, nutritional studies evaluating the effects of dietary protein and lipid levels, availability of dietary lipid sources, and substitution of fishmeal with soybean meal for growth in this species have been performed (Kim et al., 2009; Kim et al., 2010; Kim et al., 2012). Although the dietary protein requirement for 7 to 36 g juvenile far eastern catfish had been studied (Kang and Jeong, 1993), limited information about optimum dietary protein level is available for this fish species of different sizes. In an attempt to formulate an efficiently economical feed for the far eastern catfish, this study was conducted to determine the optimum protein level in the isoenergetic diets for attaining the maximal growth of young far eastern catfish.
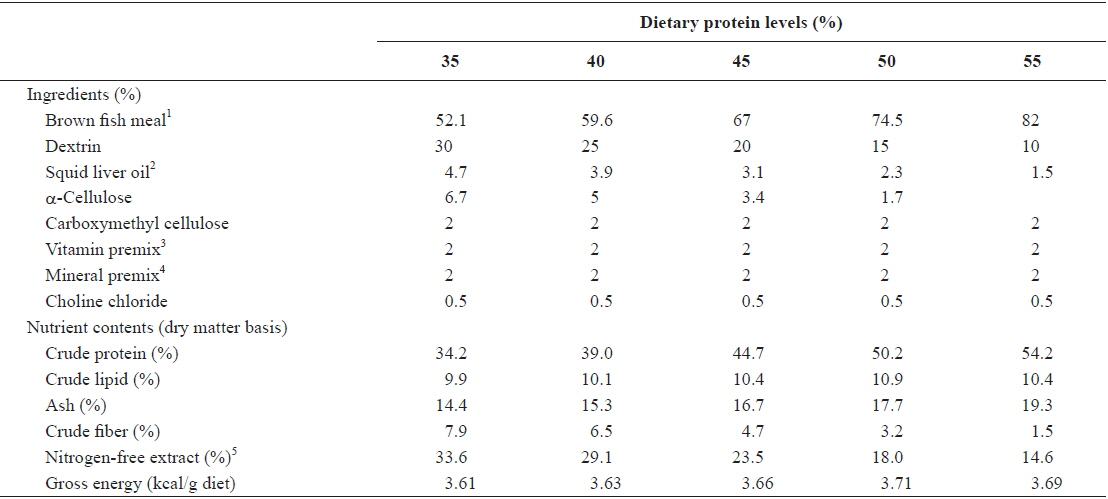
Ingredients and nutrient contents of the experimental diets are given in Table 1. Five experimental diets containing protein levels of 35, 40, 45, 50 and 55% were prepared. Gross energy levels of the diets were calculated based on 4, 9 and 4 kcal/g diets for protein, lipid and nitrogen-free extract, respectively (Garling and Wilson, 1976). Protein of fishmeal increased mainly at the expense of dextrin and α-cellulose. The dietary energy and lipid levels were designed to be isocaloric (3.7 kcal/g diet) and isolipidic (10%), respectively, by adjusting the levels of dextrin and squid liver oil. Diets were prepared with a laboratory pellet machine after 35−40 g water was mixed with 100 g mixture of ingredients, and dried over night at room temperature. The experimental diets were stored in a freezer at −24°C until use.
[Table 1.] Ingredients and nutrient contents of the experimental diets
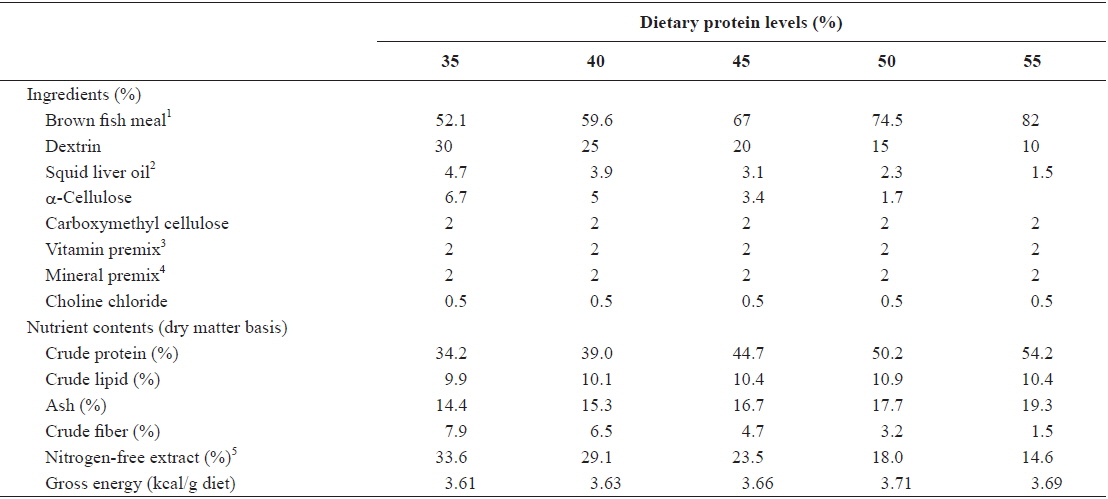
Ingredients and nutrient contents of the experimental diets
Far eastern catfish (
>
Sample collections and chemical analysis
At the end of the feeding trial, five fish in each tank were collected and stored in a freezer at -70°C for chemical analysis. Crude protein was determined by the Kjeldahl method using Auto Kjeldahl System (Gerhardt VAP50OT/TT125, KG, Germany). Crude lipid was determined by ether extraction using a Soxhlet extractor (Velp SER 148, Usmate, Italy). Moisture was determined by oven drying at 135°C for 2 h. Ash was determined by muffle furnace at 550°C for 6 h. Crude fiber was determined by an automatic analyzer (Fibertec, Tecator, Hoganas, Sweden). Nitrogen free extract was calculated by difference.
The data were subjected to one-way analysis of variance (ANOVA) using the SPSS program Version 11.5 (SPSS, Chicago, Illinois, USA). Orthogonal-polynomial contrasts were used to determine linear and quadratic effects of increasing dietary protein levels. Significant differences (
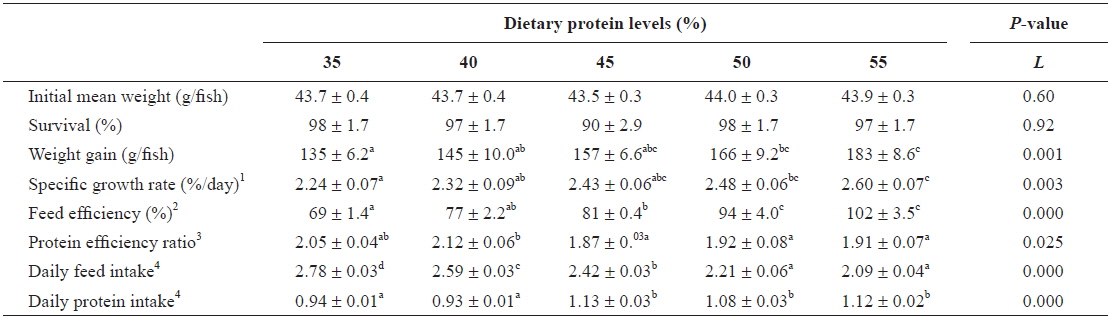
Survival of far eastern catfish in response to diets containing 35-55% protein for 9 weeks was over 90%, and there were no significant difference among the groups (
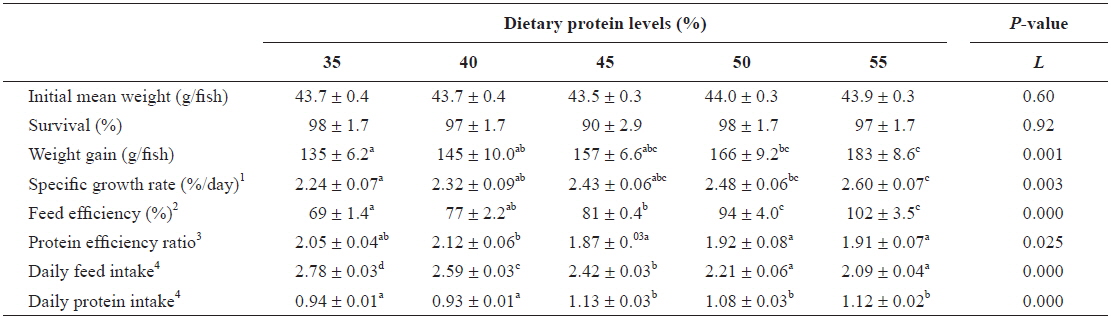
Growth performance of young far eastern catfish Silurus asotus fed diets containing the different protein levels for 9 weeks
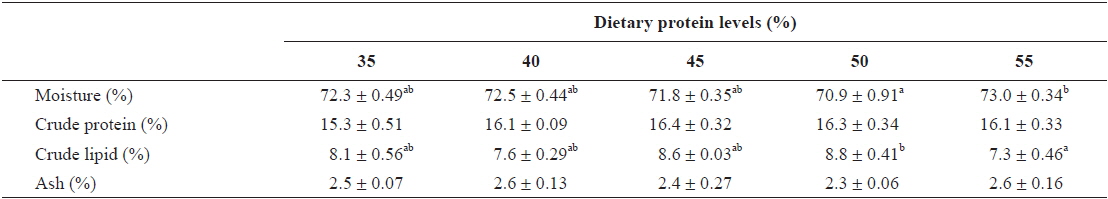
The contents of moisture and crude lipid in the whole body were significantly affected by dietary protein level (
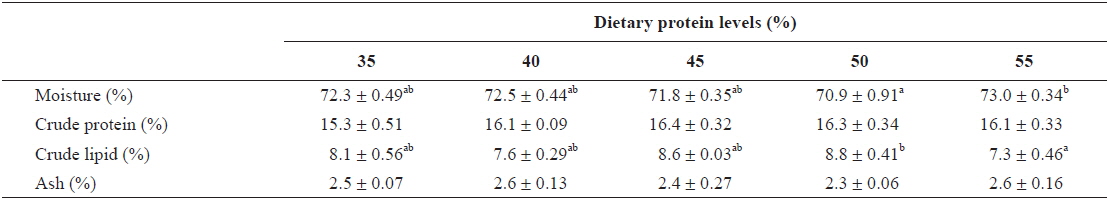
Whole body composition of young far eastern catfish Silurus asotus fed diets containing the different protein levels for 9 weeks
In the present study, growth of young far eastern catfish was increased with increasing levels of dietary protein, but weight gain and specific growth rate were no significant difference among diets containing 45 to 55% protein. The result indicates that 45% dietary protein is the optimal level for maximum growth of young far eastern catfish. The dietary protein requirement for young far eastern catfish determined in this study is comparable to protein requirements (42-44%) estimated for bagrid catfish
Protein requirements of fish are affected by fish species, fish size, water temperature, protein quality, and utilization of non-protein energy (NRC, 1993). Dietary protein requirement of 45% for young fish of 44 to 227 g determined in this study is similar with that of 45% reported for 7 to 36 g juvenile of this species (Kang and Jeong, 1993). Protein requirements, as a proportion of the diet, decrease as fish approach maturity (NRC, 1993). Protein levels of 25% was adequate in the diet of channel catfish of 114 to 500 g, but 35% protein produced improved faster gains than did 25% protein in fish weighing between 14 to 100 g (Page and Andrews, 1973). These differences among studies may be due to differences in fish species or fish size used in the studies.
Specific growth rate is a good indicator of protein quality (Martínez-Palacios et al., 2007), and decrease as fish increase in size (Lee and Kim, 2009). In this study, young far eastern catfish showed similar or slightly lower specific growth rates (2.2-2.6%/day) compared with the juveniles of the same species (2.1-3.0%/day) (Kang and Jeong, 1993), which indicated that the diets used in the present study were well utilized by young far eastern catfish. Similar findings have been reported for rockfish
In this study, feed efficiency increased with increasing dietary protein level, which can be attributed to enhanced weight gain because of the increased protein level of diet. Similar results of improved feed efficiency with increasing dietary protein level was reported for juvenile far eastern catfish (Kang and Jeong, 1993), and have also been noted for other catfish species such as bagrid catfish and Asian red-tailed cat-fish (Kim and Lee, 2005; Deng et al., 2011). Feed efficiency may also be influenced by the size of fish. Feed efficiency of 69-102% for young fish fed with the various levels of dietary protein in this study is lower than that of 86-113% reported for juvenile of this species (Kang and Jeong, 1993). In comparison to our earlier study (Kim et al., 2012), feed efficiency, protein efficiency, and daily feed intake of far eastern catfish at early life stage (initial weight 7.6 g) showed better performances than fish (initial weight 44 g) at later life stage. This result is consistent with previous studies that small fish tend to use feed more efficiently for growth than do larger conspecifics (Jobling, 1994; Lee and Kim, 2009).
This study showed decreased daily feed intake with increasing dietary protein level (r = -0.90,
Dietary protein levels could effect protein efficiency rate of fish. In this study, the higher protein efficiency rate was observed in fish fed a 40% protein diet compared to fish fed 45-55% protein diets. Similar results of decreased protein efficiency rate with increasing dietary protein level were reported in other studies (Lee et al., 2002; Kim and Lee, 2005).
In this study, dietary protein levels significantly affected whole body lipid and moisture contents, and body lipid content showed an inverse relationship with body moisture content. Similar results of an inverse relationship between body lipid and moisture contents was observed in a previous catfish study (Kim et al., 2012). Crude protein content of fish was not significantly affected by dietary protein level in this study. Similar results of no significant changes in fish body protein content when fish were fed with various levels of dietary protein were observed in previous catfish studies as well as in the studies of other fish species (Kim et al., 2012; Moore et al., 1988). However, several studies showed body protein content increased with an increase in dietary protein levels (Cowey et al., 1972; Shiau and Huang, 1989). Shearer (1994) pointed out that the proximate composition of fish is influenced by both endogenous factors such as fish size and sex as well as exogenous factors such as diet composition and the culture environment.
The results of this study suggest that the 45% protein in the diet is optimal for maximum growth of young far eastern catfish weighing in the range of 44 to 227 g.