Rapeseed (
Rapeseed is mainly cultivated for the production of house hold oil as nutritional oil and for the biodiesel production in the industry. The seed of rapeseed is rich in protein (36–40%) compared to stem and leaf, therefore, it is used in the cattle form as animal feed (Fenwick
Individual GSLs of rapeseed has become an interest for breeders because GSLs are bioactive compounds as anti-cancer or anti-pathogen agents. In the present study, rapeseed sprouts were cultured because matured rapeseeds are relatively taken long term cultivation (6–7 months) from autumn to the following spring, and their GSLs were quantified in seven rapeseed varieties. Moreover, whether GSL contents are affected or not by light and dark conditions, seed sprouts were cultured with/without fluorescent lamp in the growth chamber. Therefore, efforts have been made to select the best variety in terms of food contents thereby to increase value in local food products which have been used for many generations.
HPLC-grade acetonitrile (CH3CN), methanol (CH3OH), and sodium acetate trihydrate (NaC2H3O2 • 3H2O, 98.5%) were obtained from J.T Baker chemical Co. (Phillipsburg, NJ, USA) and Samchun Pure Chemical Co., Ltd. (Seoul, Korea). Sinigrin (2-propenyl GSL), aryl sulfatase (Type H-1, EC 3.1.6.1), and DEAE Sephadex A-25 were purchased from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Ultra-pure water used in this study was made by PURELAB Option-Q system (ELGA Lab Water, VWS Ltd., UK).
Seven varieties of
Seven varieties of rapeseed seeds (2 g) were soaked in tap water for 3 h, after that, the seeds were sown on sponge plastic pot (7 × 7 × 12 cm) and cultivated both in dark and light conditions for fourteen days in the growth chamber (temperature, 25℃; humidity, 70%). However, in the case of light cultivation condition, initially for three days the seeds were cultivated under dark condition after that from 4th day seeds were grown under 16 h light (129 μmol/m2s1) by white fluorescent lamps (32 W × 8) and 8 h dark till the end of the cultivation periods for 11 days. After cultivation, the seed sprouts were lyophilized, ground with mortar and pestle, and individually stored in a plastic bottle with a lid in desiccator until GSL analysis.
>
Extraction and desulfation of crude GSLs
Desulfo (DS)-GSLs were extracted according to the procedure of Kim
>
LC-ESI-MS/MS analysis for identification of DS-GSLs
An API 4000 Q TRAP tandem mass spectrometer (Applied Biosystems, Foster City, CA), equipped with an Agilent 1200 series HPLC system (Agilent Technologies) and an electrospray ionization tandem mass spectrometry (ESI-MS/MS) source in positive ion mode ([M+H]+), was used for the identification of the individual DS-GSLs. The MS operating conditions were as follows: ion spray voltage, 5.5 kV; curtain gas (20 psi), nebulizing gas (50 psi) and heating gas (50 psi), high purity nitrogen (N2); heating gas temperature, 550℃; declustering potential (100 V); entrance potential (10 V); spectra scanning range,
>
Separation and identification of desulfo (DS)-GSLs using HPLC
DS-GSLs obtained from different lines of cabbage were analyzed by 1200 series HPLC system (Agilent Technologies, CA, USA) equipped with an Inertsil ODS-3 (C18) column 150 × 3.0 mm i.d., particle size 3 μm) (GL Science, Tokyo, Japan). The HPLC analysis was carried out with a flow rate of 0.2 ml/min at a column oven temperature of 40℃ and a wavelength of 227 nm. The solvent system employed was ultra-pure water (A) and 100% acetonitrile (B). The gradient program used as follows: 0–18 min, 7 → 24% B; 18–32 min, keep 24% B; 32.1–40 min, keep 7% B. The individual GSLs were quantified with the sinigrin with their HPLC areas and response factors (ISO 9167-1, 1992). In this study, all the samples were designated as GSLs even though DS-GSLs were determined.
Data were analyzed by application of the Tukey’s multiple range test at P ≤ 0.05, using SPSS statistical software (version 21 for Windows, SPPS Inc., Chicago, IL, USA). The data shown in all the Tables are the means of three replicates. For comparisons of continuous parameters between groups and within a group over time, repeated measures ANOVA was used.
>
Identification of GSLs by LC-ESI-MS/MS analysis
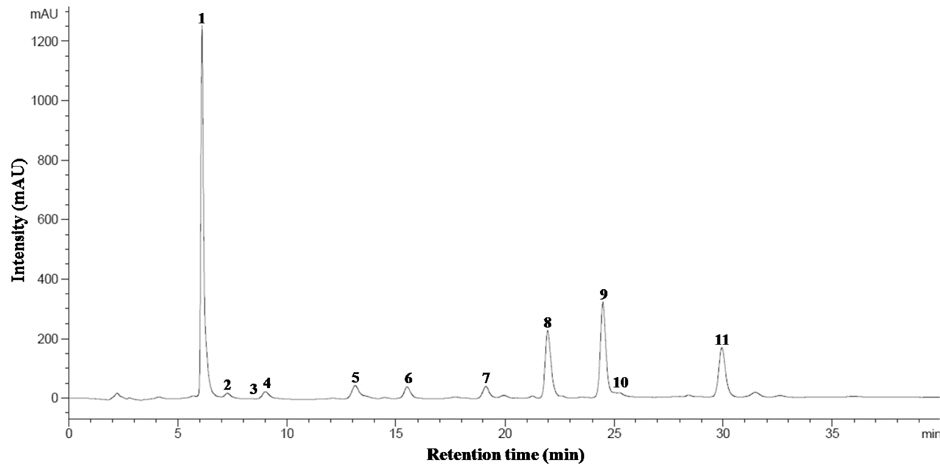
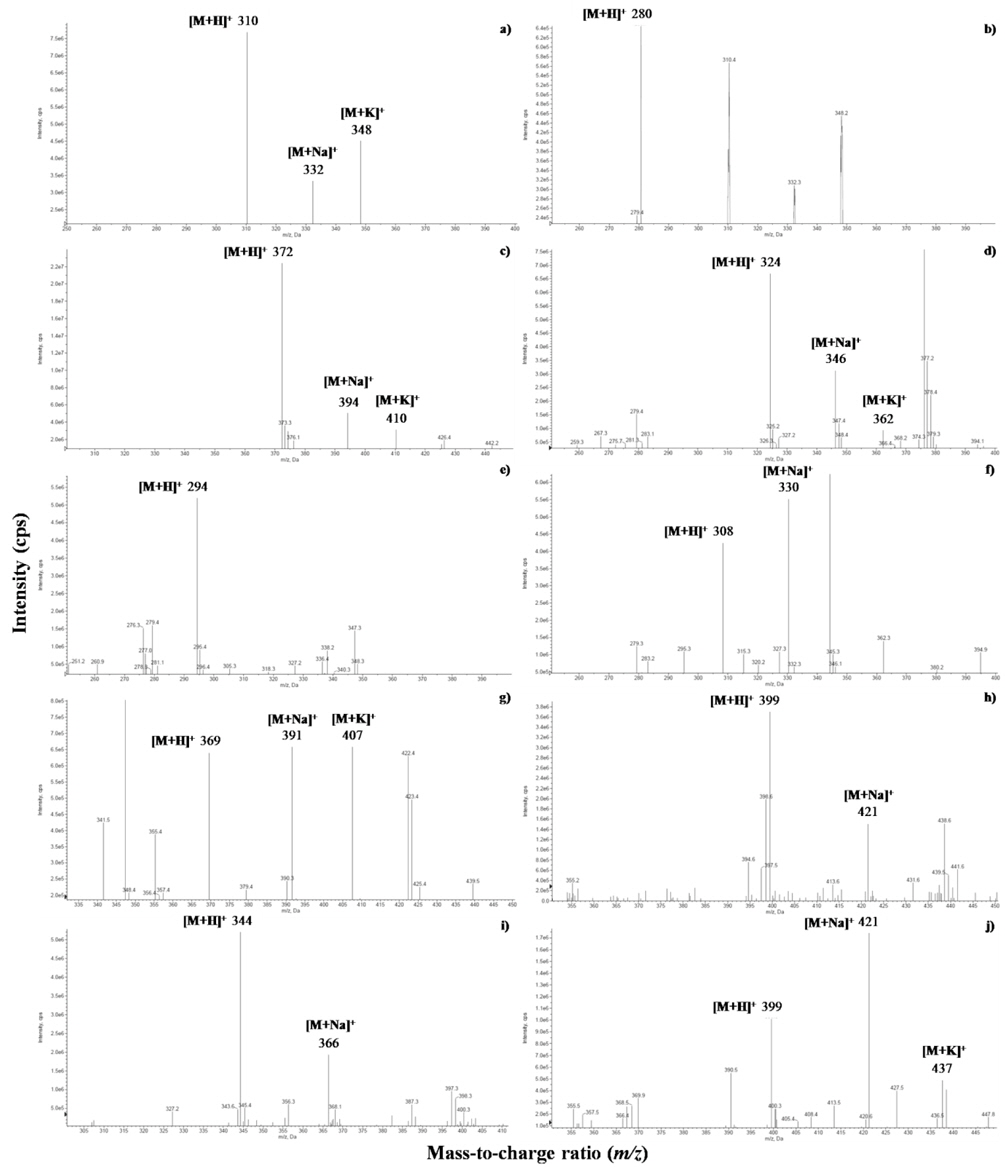
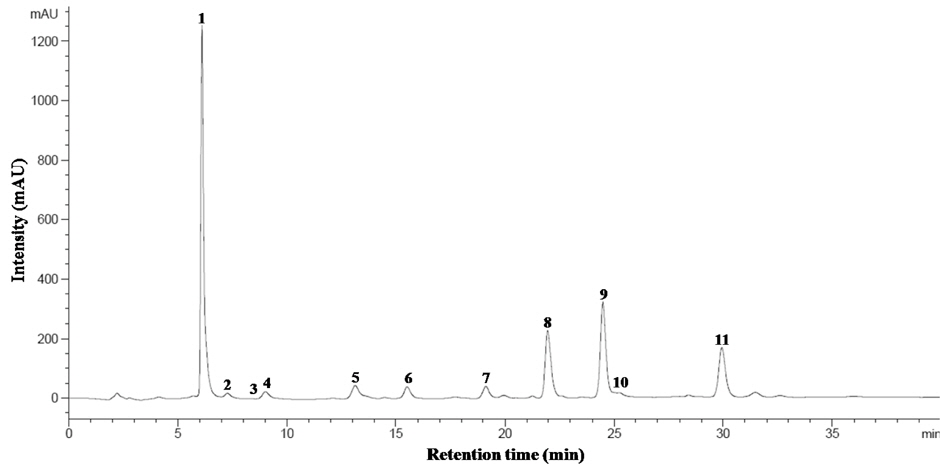
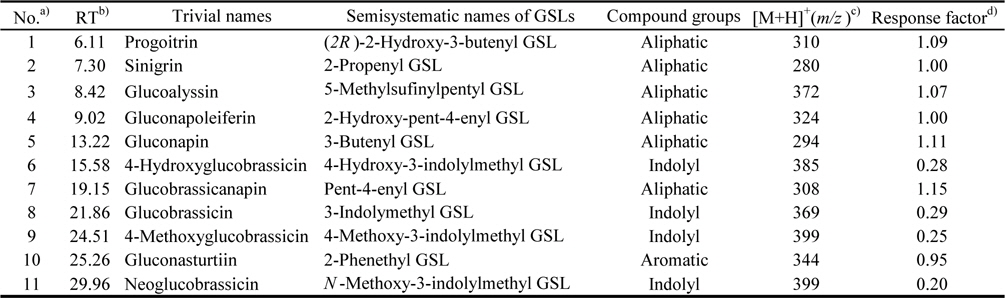
The identified compounds by LC-ESI-MS/MS analysis in positive ion mode [M+H]+, including systematic and common names and the principal ions are listed in Table 1. The identified GSL in seven varieties of rapeseeds were very similar, and eleven GSLs were detected in all extracts. Each GSLs were identified based on their protonated molecular ions [M+H]+ and corresponding product ions such as sodium [M+Na]+ and potassium [M+K]+ adducts. Six aliphatic (progoitrin, sinigrin, glucoalyssin, gluconapoleiferin, gluconapin, and glucobrassicanapin), four indolyl (4-hydroxyglucobrassicin, glucobrassicin, 4-methoxyglucobrassicin, and neoglucobrassicin), and one aromatic GSLs (gluconasturtiin) were identified based on fragmentation patterns of MS spectra and quantified based on the peak areas of HPLC chromatogram (Fig. 1 and Fig. 2). Peaks 1 and 5 were identified as progoitrin and gluconapin, respectively. MS data revealed that these two GSLs contained 3-butenyl group as R-groups backbone with variation in the hydroxyl moiety in their structure (
[Table 1.] Glucosinolates (GSLs) identified by LC-ESI/MS in rapeseed sprouts
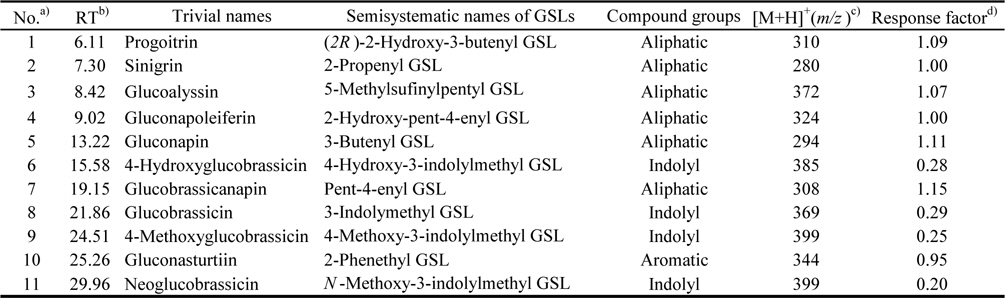
Glucosinolates (GSLs) identified by LC-ESI/MS in rapeseed sprouts
>
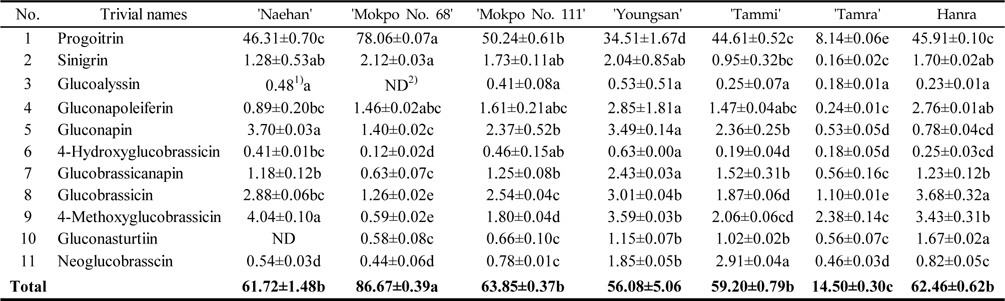
Variation of GSLs under dark condition
The total GSL contents of the rapeseed sprouts cultivated under dark condition were ranged from 14.50–86.67 μmol/g DW (Table 2). Based on various reports, the major GSLs in rapeseeds are progoitrin, gluconapin, glucobrassicanapin, and glucobrassicin (Font
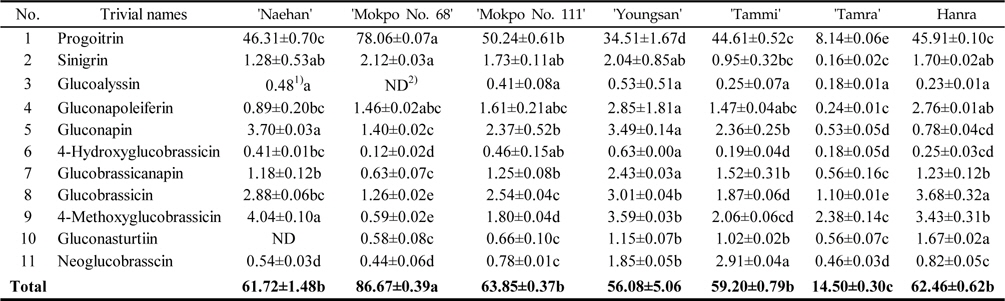
Glucosinolate contents (μ mol/g DW) in rapeseed sprouts cultivated under dark condition (n=3)
>
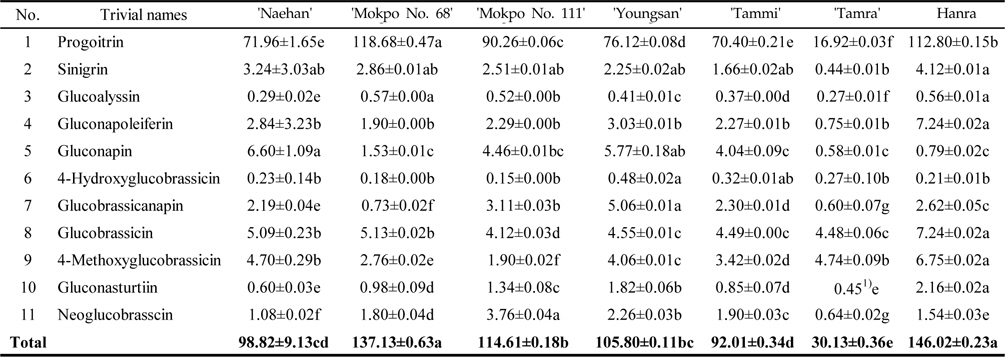
Variation of GSLs under light condition
The total GSL contents of the rapeseed sprouts cultivated under light condition were ranged from 30.13–146.02 μmol/g DW (Table 3). Under light condition the total GSL contents of the sprouts of ‘Hanra’ were higher (146.02 μmol/g DW) followed by ‘Mokpo No. 111’, (137.13 μmol/g DW). It is noted that rapeseed cultivated under light condition exhibited comparatively 1.83 fold higher GSL than under dark condition. Similar to dark condition, progoitrin identified as the major GSLs with higher contents in all the varieties together with gluconapin and shared 74.3% and 3.45% of total GSLs. In ʻHanraʼ, the predominant GSLs were progoitrin (112.80 μmol/g DW) followed by gluconapoleiferin (7.24 μmol/g DW), glucobrassicin (7.24 μmol/g DW), 4-methoxyglucobrassicin (6.75 μmol/g DW), sinigirin (4.12 μmol/g DW), glucobrassicanapin (2.62 μmol/g DW), gluconasturtiin (2.16 μmol/g DW), and neoglucobrasscin (1.54 μmol/g DW), whereas, glucoalyssin, gluconapin and 4-hydroxyglucobrassicin documented less than 1.0 μmol/g DW. Under light cultivation condition the contents of indolyl GSLs varied significantly. Glucobrassicin is the most abundant indolyl GSLs, representing nearly 44.8% of the indolyls and about average 5.92% of the total GSLs (Table 3). The highest level of glucobrassicin in rapeseed sprout was found in the ‘Hanra’ (7.24 μmol/g DW) and ‘Mokpo No. 68’ (5.13 μmol/g DW), while the lowest concentration was found in ‘Mokpo No. 111’ (4.12 μmol/g DW). The sum of residual indolyl GSLs (4‐hydroxyglucobrassicin, 4‐methoxyglucobrassicin, and neoglucobrassicin) was less and exhibited comparatively significant differences in their contents among the rapeseed varieties, except in ‘Hanra’ which contained a significant amount of 4‐methoxyglucobrassicin (6.75 μmol/g DW). In contrast to indolyl GSLs, aliphatic GSLs such as progoitrin, sinigrin, glucoalyssin and gluconapoleiferin are mainly synthesized under the control of genetic materials (Magrath
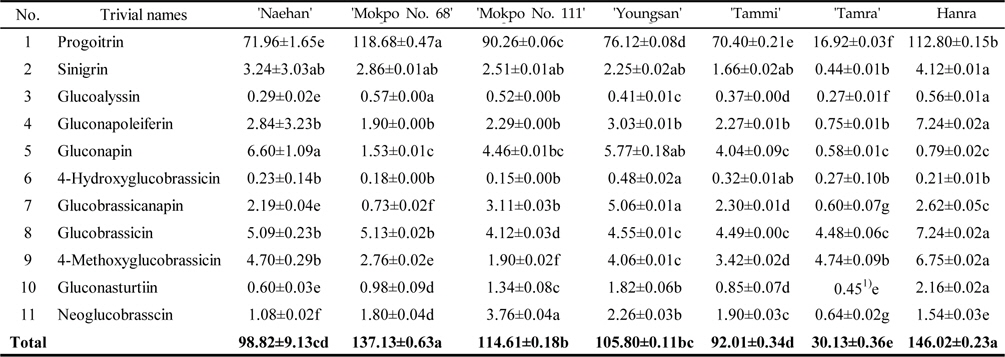
Glucosinolate contents (μ mol/g DW) in rapeseed sprouts cultivated under light condition (n=3 )
Rapeseed sprouts have similar GSL profiles to irrespective of their cultivation condition, and progoitrin is the most abundant GSLs. The total contents of GSLs in light cultivated rapeseed sprouts are 1.55–2.33 times higher than those in dark cultivated sprouts. Total GSL contents were higher in ‘Hanra’ and ‘Mokpo No. 68’ (146.02 and 137.13 μmol/g DW, respectively). The glucobrassicin contents of sprouts grown under light are much higher than those of the dark, but the total GSL contents in other indolyl groups are comparatively similar. In both dark and light cultivation conditions, the total contents of progoitrin are much higher than those of other individual GSLs. From this study it is confirmed that dark growing condition is better if our interest to reduce the content of GSLs, whereas the light grown condition is preferable if research is interested in biodiesel production. In the rape crop breeding program, continuing efforts have sought to decrease the contents of individual GSLs, for food and safety reasons. To decrease the content of antinutritional components in rapeseeds, a new variety has to be made through pollination and breeding methods, which attracts the rapeseed crop growers and the animal feed industries for the production of cattle feed.

![Mass spectra of desulfoglucosinolates in rapeseed sprouts (‘Mokpo No. 68’). 1, progoitrin (m/z 310 [M+H]+); 2, sinigrin (280); 3, glucoalyssin (372); 4, gluconapoleiferin (324); 5, gluconapin (294); 7, glucobrassicanapin (308); 8, glucobrassicin (369); 9, 4methoxyglucobrassicin (399); 10, gluconasturtiin (344); 11, neoglucobrassicin (399). Peak 6, 4hydroxyglucobrassicin was identified based on our data base and HPLC retention time.](http://oak.go.kr/repository/journal/15156/HGNHB8_2013_v32n4_315_f001.jpg)