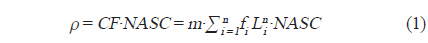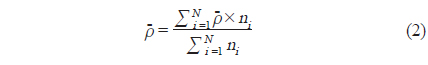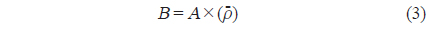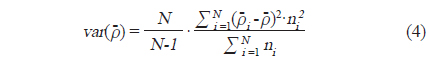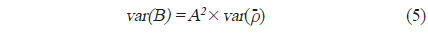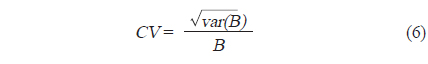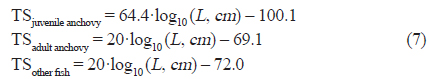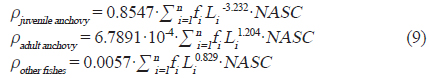The distribution and abundance of coastal fish species in Asan Bay, Korea, were estimated from hydroacoustic survey and net catches. Acoustic data were collected with 38 and 200 kHz from July to October of 2012, and converted to the nautical area scattering coefficient (
우리나라 황해 내만 및 연안역은 생물의 다양성이 높은 해역으로 어류의 산란장 및 보육장으로 이용된다(Kim et al., 1994; Lee and Hwang, 1995). 그 중 아산만은 갯벌이 넓게 발달되어있어 갑각류와 무척추 동물이 대량으로 서식하여 다양한 연안어류 및 회유성 어류에 좋은 서식처로 생태계에 매우 중요한 역할을 담당하는 해역이다(Cha et al., 2013). 하지만 1970년대 이후 방조제 건설 및 공업 단지의 조성 등의 지속적인 산업화 활동에 의한 환경 오염의 증가로 해양 생물의 서식처가 위협받고 있는 해역이다(Kim et al., 1994). 또한 아산만 및 남양만 인근해역은 삽교 및 아산방조제, 서해대교, 남양방조제, 화용방조제, 석문지구방조제 등의 건설과 석문공단, 인주공단 등 시설공사로 인한 해양 환경의 변동이 높아 어류의 분포 및 자원량에 영향을 미칠 가능성이 높은 해역으로 알려져 있다(Lee and Hwang, 1995).
국내에서 해양 및 육상 담수역에서 수산자원의 시·공간 분포 및 자원량 파악을 위한 과학적인 조사는 1960년 이후 현재까지 주로 어구 어업을 이용하였다(Choi, 1971; Huh, 1986; Cha et al., 2004). 어구 조사시 어란 및 유생은 봉고 네트로, 어류 유생 및 성체는 중층 및 저층 트롤과 같은 대형 어구로 채집 후 종조성 및 체장 특성을 파악하였다. 1990년 이후로는 어류 자원량추정 모델을 이용하는 간접적인 방법으로 자원량 변동을 추정하였다(Jang et al., 1992; Choi et al., 1999). 아산만 인근의 어류 자원 조사 또한 현존하는 어류의 종조성에 초점을 둔 어구 어법을 통해서 실시 되었다(Lee and Kim, 1992; Lee, 1993; Cha et al., 2013).
전통적인 어구 채집 방법은 정성적인 자료 측면에서는 많은 장점이 있지만 채집 면적 및 채집 수층의 제한성이라는 측면에서 정량적인 자료 도출에 오차를 만들어 낼 가능성이 크다. 또한 작업시간에 대한 비용 증가 요인이 크기 때문에 조사 효율성 측면에서 단점이 존재한다. 정성 분석에 치우친 기존 네트 채집방법의 문제점을 극복하기 위한 방법 중 하나로 1980년 초부터 제시되어 최근에 생물의 분포 및 자원량 조사에 활발히 적용하고 있는 생물 음향을 이용하는 방법이 대두되고 있다(MacLennan and Simmonds, 1992; Abad et al., 1998). 생물 음향 기법의 특징은 연구자의 주관을 배제시키고 사전에 설정된 음향 조사 정선으로부터 연속적이며 전 수층에 대한 객관적인 자료를 얻을 수 있는 장점이 있고, 우점 어종의 음향 반사강도(acoustic target strength, TS) 특성이 제공 된다면 조사 해역의 현존량까지 추정이 가능하다(Simmonds and MacLennan, 2005).
우리나라에서 본격적으로 생물음향 기법을 사용한 것은 2000년대 초 연근해 바다목장 조성 해역에서 자원량 평가에 적용되었다(MOF, 2002; MOF, 2006; MOF, 2007). 2001년 통영 해역에서 시작되어 여수 남단의 다도해 지역 및 제주도 연안에서 진행되었고, 주로 어군의 공간 분포에 초점을 두고 계절 별로 실시되었다. 최근에는 동중국해, 통영 및 제주 연안에서 동물플랑크톤, 멸치 및 저서 어류의 자원량 추정에 활용하면서 생물음향기법의 활용도가 증가하고 있다(Kang et al., 2003; Kim et al., 2008; Lee et al., 2012).
본 연구는 경기도 화성시에 속해있는 아산만 연안 해역에서 생물음향 기법을 이용하여 하계−추계 시기에 출현하는 어류의 분포 특성을 음향 기법을 이용하여 파악하였고, 조사해역의 주요 어종인 멸치와 기타 어류의 자원량을 추정하였다. 이를 위하여 2012년7월부터 10월 사이 동일한 해역에서 음향 조사를 실시하였고, 음향 조사와 동시에 어구 어업을 병행하였다. 획득한 음향 자료와 어획 자료를 이용하여 어류의 시·공간적인 분포와 다중 주파수를 이용한 생물음향 처리 기법을 활용하여 조사 면적 내 멸치와 기타 어종의 현존량을 제시하였다.
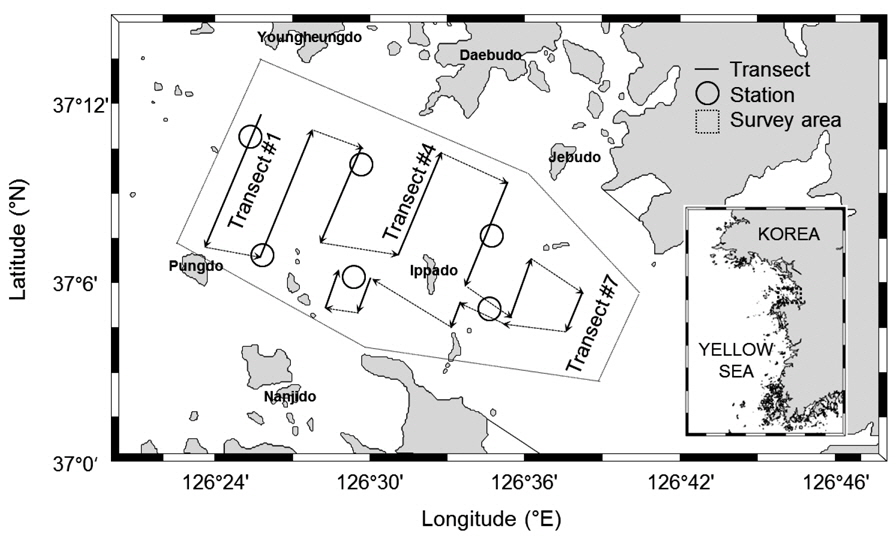
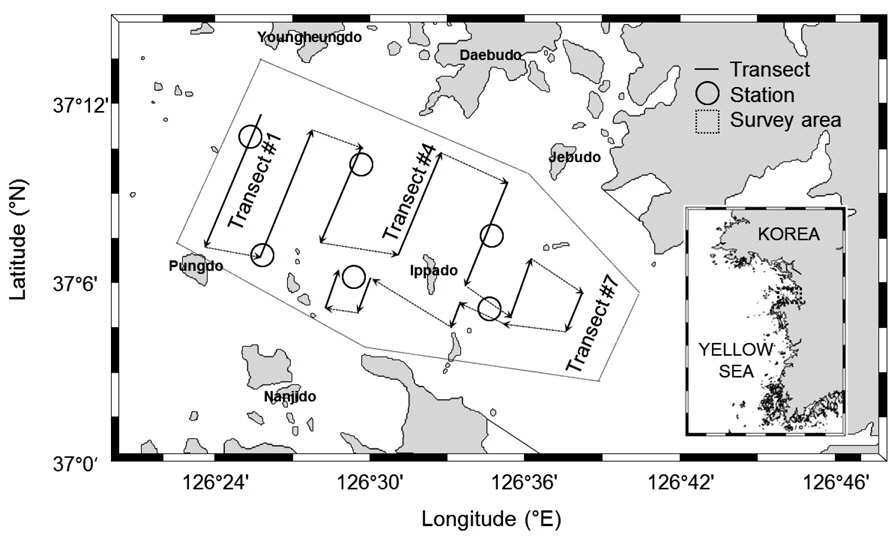
음향 조사가 실시된 해역은 경기도 아산만 내측의 “화옹 방조제−석문 방조제”와 외측의 “영흥도−풍도” 까지 구역으로 총 조사면적은 186 km2 (54 n·mile2)이다. 음향 조사는 2012년 하계부터 추계시기인7월부터 10월까지 총 4회 실시하였다. 음향 정선은 아산만 중앙수로를 가로지르는 정선 1−7 번과 입화도와 당진 화력발전소 사이의 정선8−10번으로 10개를 선정하였고, 총 음향 정선 거리는 26.3−28.0 n·mile 이다(Fig. 1). 음향 정선은 조사선 안전을 확보하기 위하여 주변 어망과 조수 간만의 차이에 따른 5 m 미만의 수심에 대해서 조사시기 별로 부분적인 변동이 있었다. 정선 간 이동을 위한 거리는 음향 정선 계산시 제외하였다. 음향 조사 방식은 아산만 내 섬 주변과 연안 조사를 고려하여 정선 사이의 간격 변동이 있는 자유 정선(random parallel transect) 방식을 사용하였다.
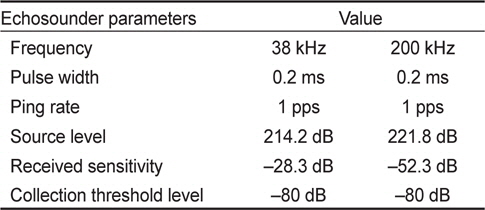
음향 조사를 위한 과학어군 탐지기(scientific echosounder)는 시스템 제어부, 센서부 및 예인체로 구성된 통합 시스템(Bio-Sonics DT-X series)을 사용하였다(BioSonics, 2005). 수신된 음향 신호로부터 종 분류를 위해서 38 kHz와 200 kHz의 다중주파수로 음향 신호를 획득하였다. 2개의 음향 센서는 예인체(towed body)에 설치하여, 조사선의 측면에 고정 후 해수면으로부터 약 0.5 m의 심도를 유지하며 음향 자료를 획득하였다. 수심이 얕은 천해 환경에서 조사선의 측면에 설치한 예인체의 음향 센서는 조사선의 움직임에 의해서 해저면 신호 및 어류 탐지 신호의 수신 값에 큰 영향을 준다. 본 연구에 사용된 디지털음향 센서는 조사선 방위 및 센서 기울기를 측정하는 독립적인 센서가 내부에 있어, 음향 시스템 운영시 실시간으로 수신되는 해저면 신호 세기와 센서 기울기 자료로부터 예인 속도를 조절하였다. 음향 조사 시 안정된 신호 수신을 위해서 센서의 pitch와 roll 허용 각도는 ±7°, 예인속도는 약 6노트 이내로 유지하며 자료를 획득하였다.
음향 자료는 어군 전체의 분포 특성 및 강도를 나타내는 체적후방 산란강도(Volume backscattering strength, SV)를 위치 자료와 함께 획득하였다. 음향 자료 처리에 필수적인 음향 수신 신호의 위치 정보는 조사 주변의 섬과 육지에 근접하여 음향 조사를 실시하고, 또한 조사 면적이 원양이 아닌 좁은 해역임을 고려하여 기존의 GPS보다 정확한 DGPS (differential global positioning system)를 사용하였다.
음향 조사에 사용한 송신 신호의 길이를 나타내는 펄스 폭(pulse length)는 얕은 수심과 상대적으로 고분해능의 수층 자료를 고려하여 0.2 ms로, 각각의 ping 간격(ping interval)은 1초로 설정하여, 38 kHz와 200 kHz 주파수 동시에 음향 신호를 송·수신하였다. 음향 조사시 해수의 계절적인 변동에 의한 환경요소(수온, 염분)를 제외한 시스템 변수는 동일하게 설정하였다(Table 1).
[Table 1.] System parameters of scientific echosounder for acoustic survey
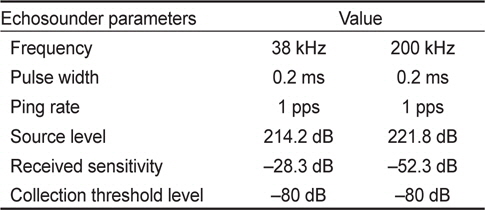
System parameters of scientific echosounder for acoustic survey
음향 자료는 가상 에코그램 기법을 이용하여 동일한 시간 및 수심에 대한 복수의 주파수에서 획득한 자료 분석을 실시하였다(Myriax, 2008). 자료 처리는 두 주파수에서 획득한 음향 자료(raw data)을 가상 에코그램 기법을 활용하여 자료의 질이 불량한 영역에 대한 잡음 제거(Mask bad data), 거리 및 수심에 대한 자료의 압축(Resample by Time & Depth), 종 분리를 위한 음향 산란강도의 차이 에코그램 생성(dB difference) 과정을 거쳐 최종적으로 새로운 에코그램(New echogram & SV)을 생성하였다.
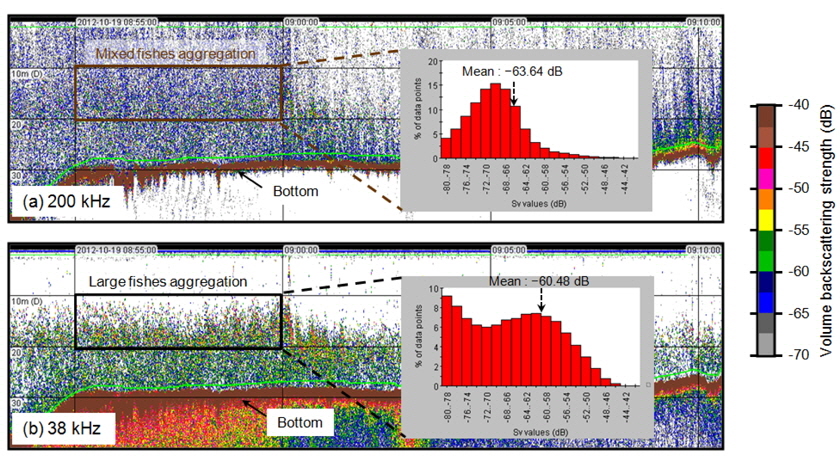
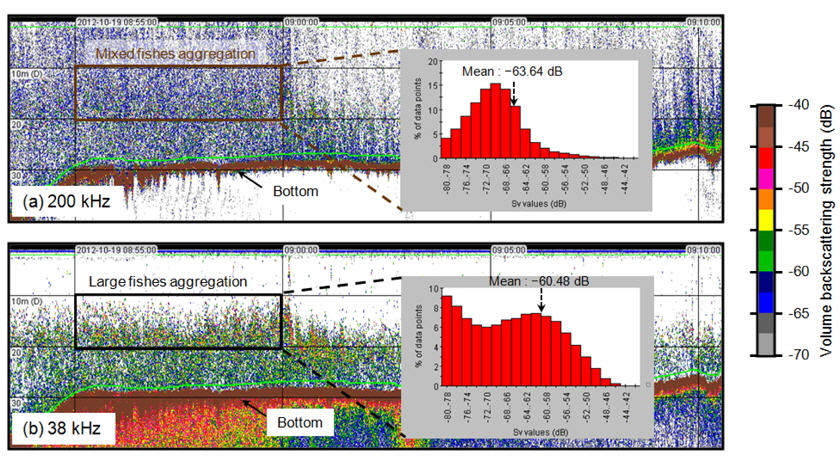
Fig. 2는 2012년 하계시기(7월)에 가장 외측인 정선 1에서 취득한 38 kHz와200 kHz 에코그램의 예이다. 동일한 음향 산란체가 존재하지만 200 kHz에서는 전층에 걸쳐 소형 어류에 판단되는 푸른색 계열의 –65 ~ –60 dB의 범위가 높게 나타나며, 대형 어류에 의한 높은 산란강도(>–60 dB)의 신호도 혼재되어 수신되었다. 반면에 상대적으로 낮은 주파수인 38 kHz 신호에서는 강한 어류의 신호를 의미하는 중층의 빨간 계열 신호(>–50 dB)가 뚜렷하게 나타나며, 해저면 부근에서도 녹색 계열의 –60 ~ –55 dB 의 어류 신호가 존재하였다. 동일한 시기, 동일한 위치, 동일한 음향 산란체가 존재함에도 불구하고 SV는 주파수에 따라 분포 종의 음향 반사 특성이 다름을 의미한다. 동물플랑크톤이나 체내에 공기나 부레가 없는 생물체는 일반적으로 저주파에서 약한 반사강도 특성을 가진다(Miyashita et al., 1997). 그러나 체내에 부레가 있는 어류는 저주파에서도 상대적으로 강한 음향 반사를 일으킨다(Simmonds and MacLennan, 2005). 따라서 종 별 신호 분리를 위해서 복수의 주파수로 수신된 각각의 음향 신호로부터 음향 산란강도의 차이법(Difference of Mean Volume Backscattering Strength, △MVBS)을 적용하였다(Madureira et al., 1993; Watkins et al., 2002; Kang et al., 2003).
조사해역의 주요 우점종은 당해년도에 산란되어 성장한 멸치로(Cha et al., 2013), 멸치 자치어의 경우 주간시기 부레에서 공기를 배출하고, 야간시기에 부레에 공기를 채우는 것으로 알려져 있다(Miyashita et al., 2003). 본 음향 조사는 조사선의 안전을 고려하여 주간시기에 조사가 진행되었다. 따라서 멸치 자치어와 다른 산란체 (중층어류 및 동물플랑크톤)를 구별 하는 기준을 고려하여 고주파와 저주파 사이의 체적 산란강도 차이의 범위는 “2 dB < △MVBS200-38 kHz < 20 dB” 로 설정하였고, 이 설정 값을 사용하여 전제 에코그램에서 멸치 치어 크기의 신호만을 분리하여 SV를 생성하였다(KIOST, 2014). 멸치 치어와 달리 체내에 부레를 가지고 있는 기타 어류에 대한 신호는 일반적인 어류의 체적 산란강도 차이 범위인 “2 dB < △MVBS38-200 kHz < 20 dB” 로 설정하여 SV를 생성하였다(Simmonds and MacLennan, 2005). 생성된 비선형 형태의 SV 자료는 어군의 시·공간적인 분포 특성 및 현존량 분석을 위해 선형 형태인 면적 산란계수(Nautical Area Scattering Coefficient,
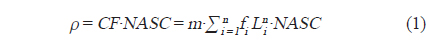
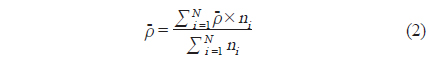
식(2) 에서 는
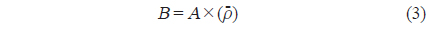
각각의 정선 평균 밀도와 전체 조사지역의 밀도로부터 전체 조사지역에서 평균 어군 밀도의 변동(variance,
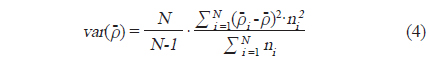
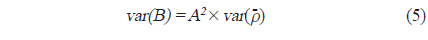
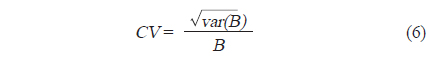
아산만 내 어군의 음향 밀도 및 자원량 추정을 위한 보조자료로 조사 구역 내 6개의 정점에서 어류를 채집하였다(Fig. 1). 어획 조사는 2012년 7−10월 음향 조사 시기와 동일 시기에 실시하였다. 채집시 사용한 어구는 안강망(stow net, 24 m (망고)×35 m (망폭)×50 m (길이), 망목 0.85 mm)으로 각 정점에서 투망하여 1시간 동안 설치 후 양망하였다. 채집 된 어류는 현장에서 얼음과 함께 아이스박스와 함께 냉장 보관한 뒤, 실험실로 운반 후 어종 별 체장 및 무게를 측정하였고, 종조성을 분석하였다. 어류의 동정은 Kim et al. (2005)의 기준을 따랐고, 채집량이 많은 어종에 대해서 부표본하여 측정 후, 전량으로 환산하였다.
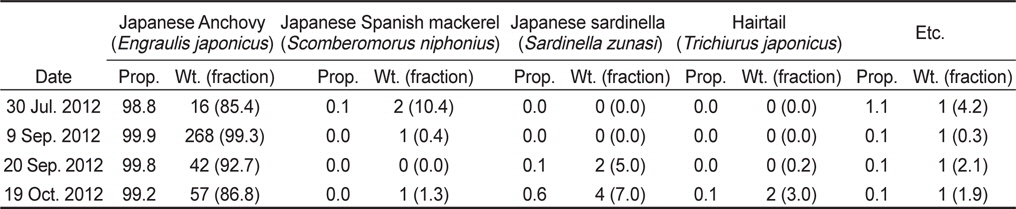
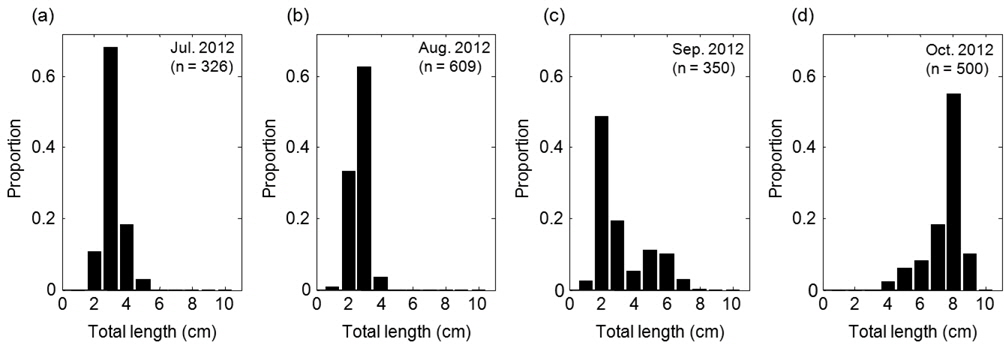
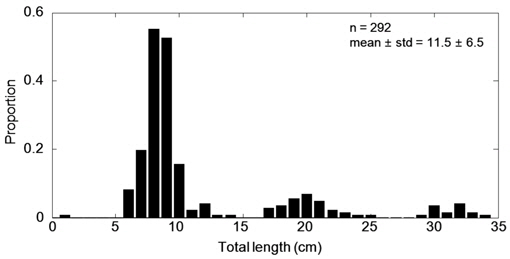
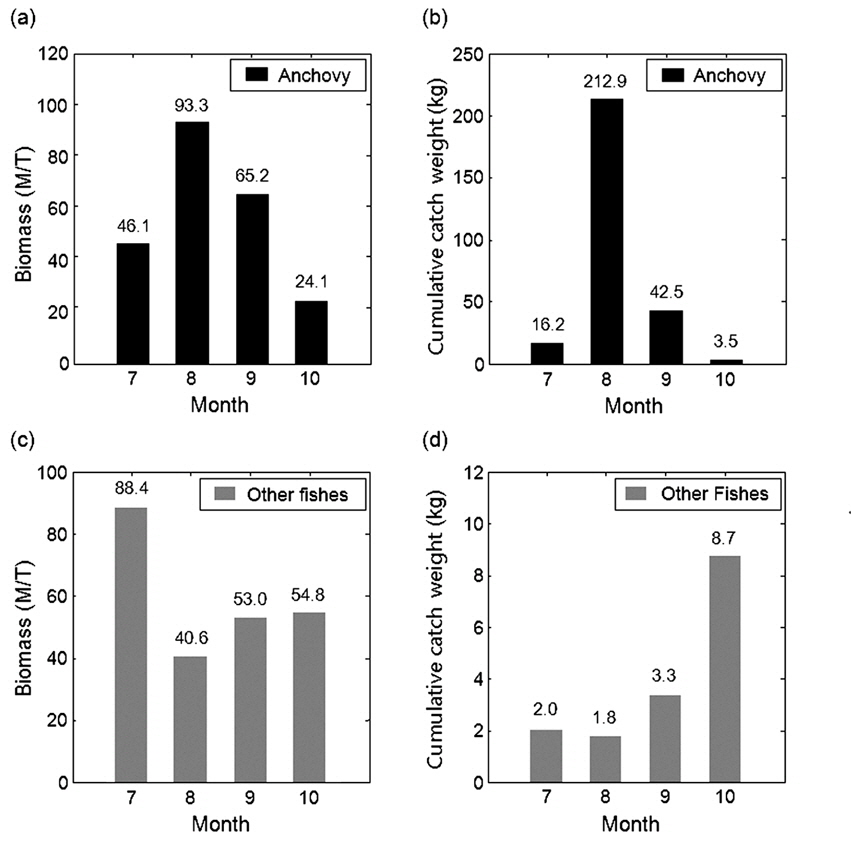
아산만 인근해역에서 2012년 7월부터 10월 조사기간 동안 채집된 어류는 총 개체수 3,487,395마리이고, 총 생체량은 400,782 g이었다(Table 2). 조사 해역의 최우점종은 멸치(
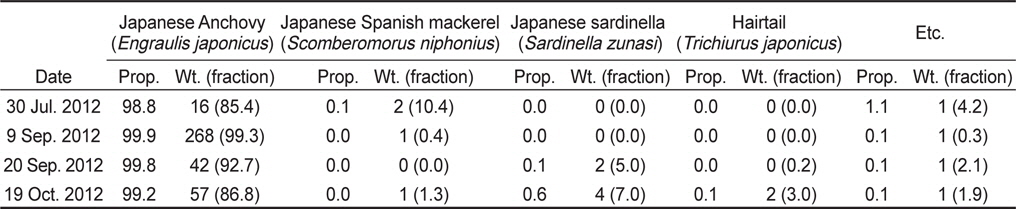
Number (No.) of stow net catches with each fish species; their relative proportion of individuals (Prop.; %) in catches species; total cumulative catch weight (Wt.; kg); and fraction of the combine species catch weight
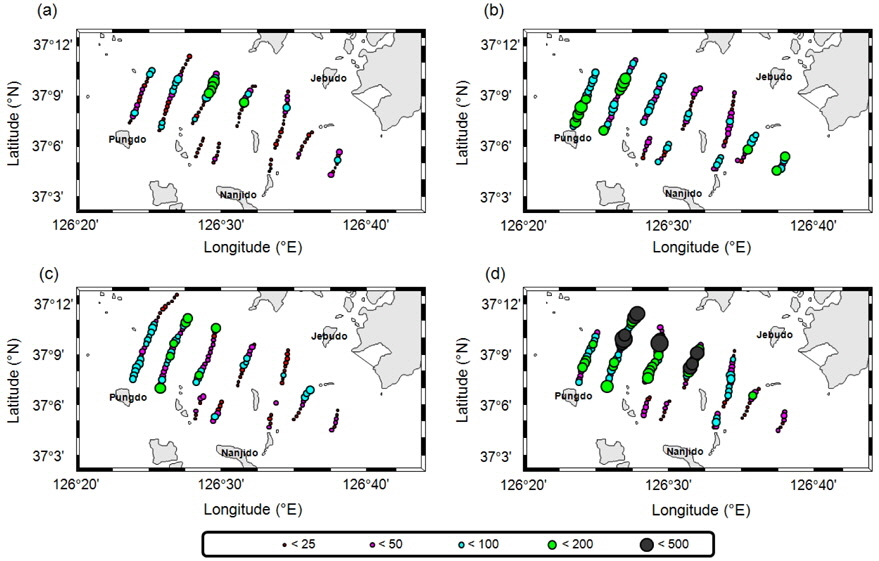
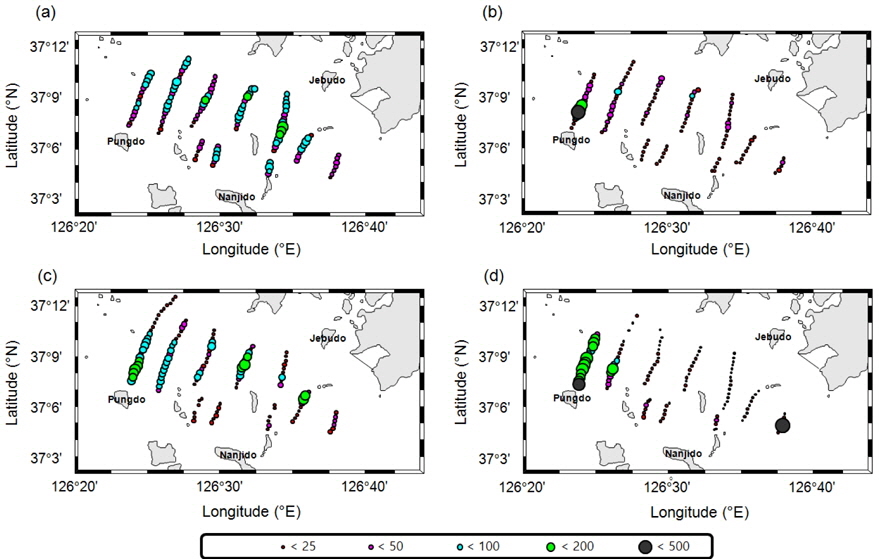
조사해역에서 획득한 음향 자료에 대해서 음향 산란강도 차이법(△MVBS)으로 치와 기타 어류 신호로 분리 후, 추출한 각각의
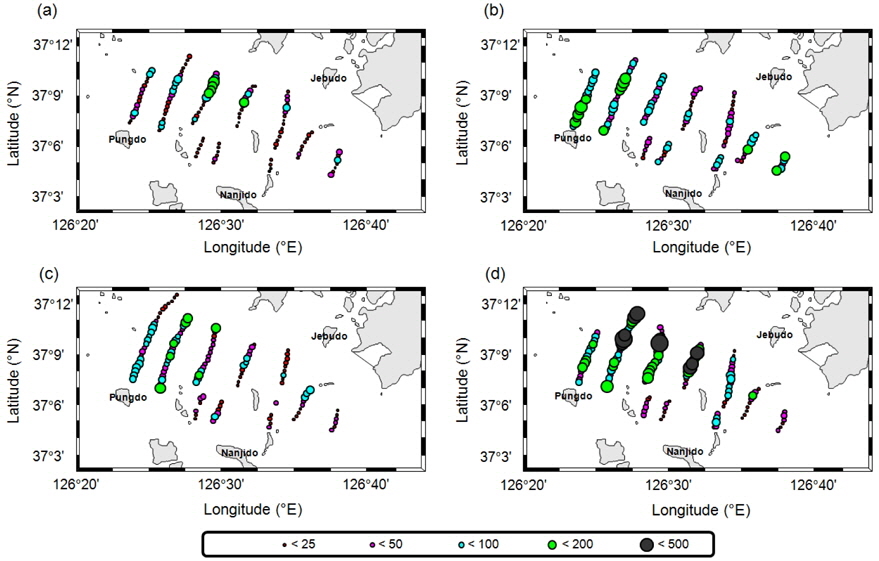
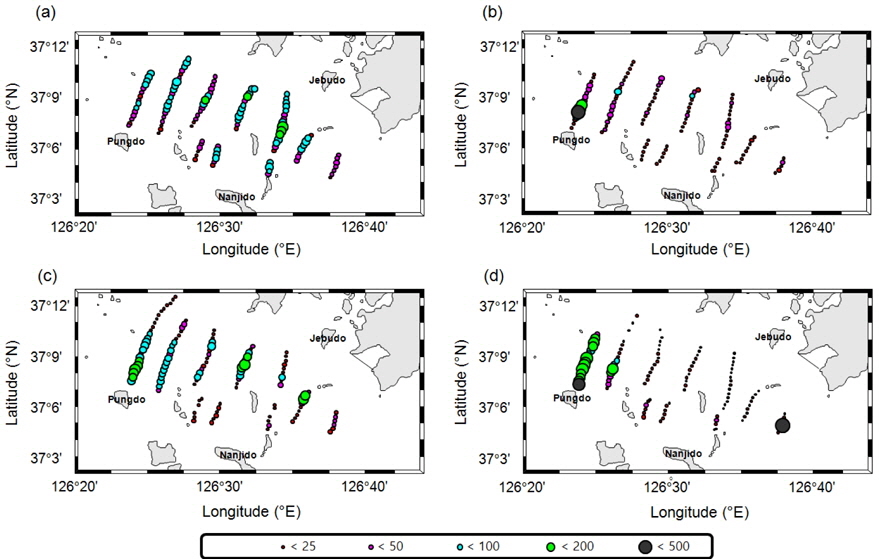
멸치를 제외한 기타 어류는 7월의 경우 전체적으로 전 정선에서 고른 분포를 보이며, 높은
음향 조사를 통해서 획득한 음향자료와 어구 어법을 통해서 채집된 어획자료를 이용하여 멸치와 기타 어류의 현존량을 추정하였다. 추출된
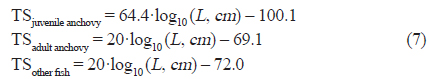
어획자료를 이용한 멸치와 기타 어류의 길이−무게 함수는 다음과 같이 유도 하였고(식 8), 기타 어종의 경우 멸치를 제외한 채집된 어종 중 생체량이 1 % 이상인 삼치, 밴댕이, 갈치에 대한 길이−무게 분포를 이용하였다(Table 2).

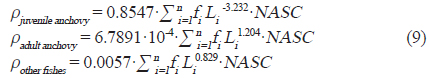
여기서
현존량 산출은 계산된 CF를 이용하여 단위 밀도(
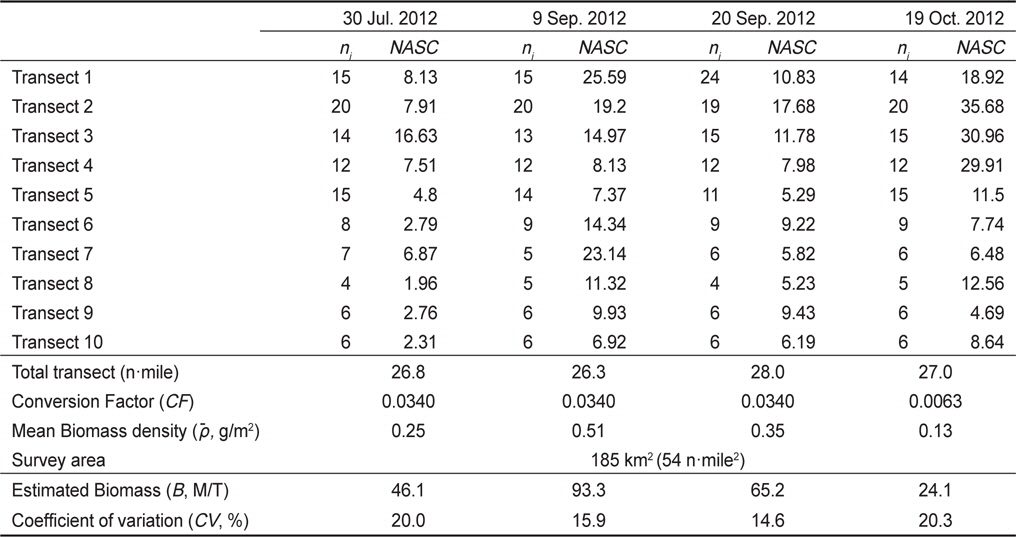
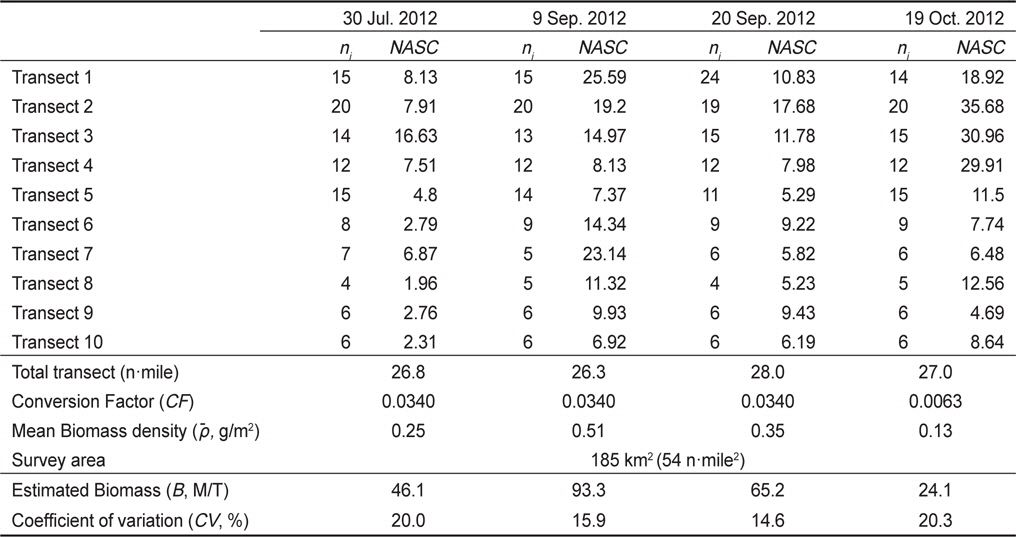
Standing stock estimation of Engraulis japonicus using acoustic survey in Asan Bay, Korea 2012. The ni is the number of 0.25 EDSU and the NASC is mean value of the nautical area scattering coefficient (m2/mile2) along i-th transect
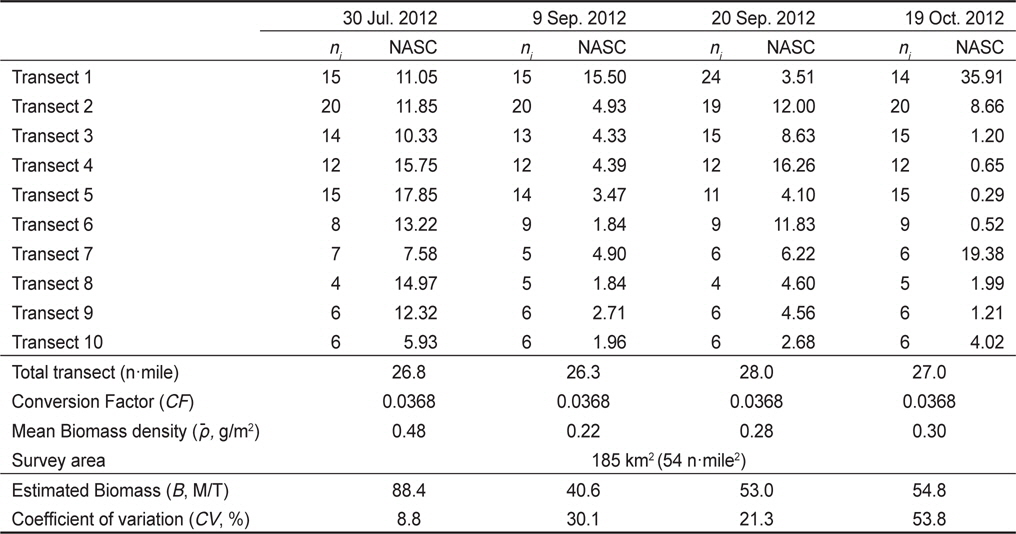
멸치를 제외한 기타 어종의 현장 밀도는 0.22−0.48 g/m2의 범위를 가지고 있으며, 시기 별로는 7월 0.48 g/m2, 8월 0.22 g/m2, 9월 0.28 g/m2, 10월 0.30 g/m2이었다(Table 4). 상대적으로 높은 밀도는 7월이고, 낮은 밀도는 8월이었다. 7월과 8월의 밀도 차이는 약 2.2 배였다. 평균 밀도로부터 기타 어종은 현존량은 7월 88.4톤(
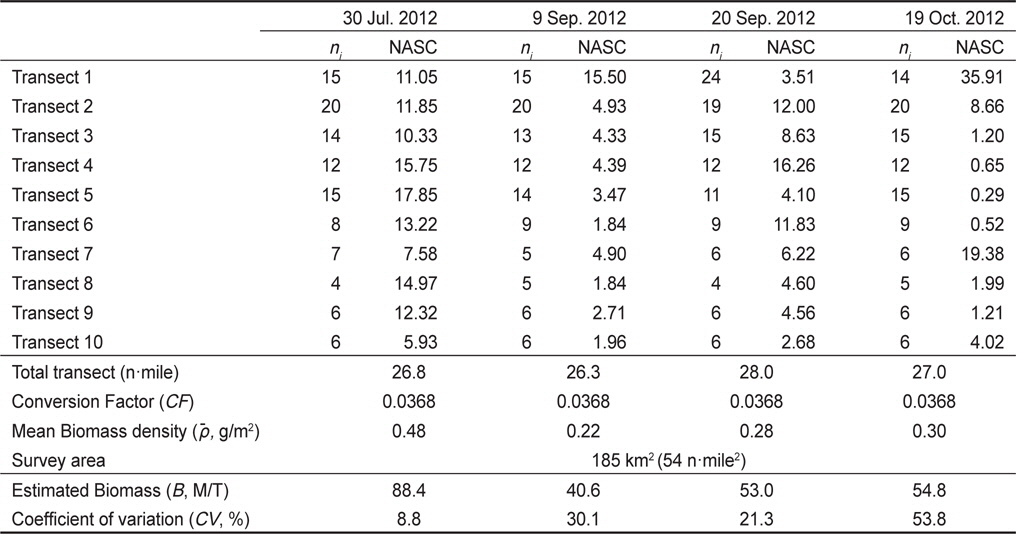
Standing stock estimation of fish without Engraulis japonicus using acoustic survey in Asan Bay, Korea 2012. The ni is the number of 0.25 EDSU and the NASC is mean value of the nautical area scattering coefficient (m2/mile2) along i-th transect
아산만 내의 멸치(
주간시기 하층부, 야간시기 상층부에 존재하는 일반적인 어류분포와 달리 하계시기에는 멸치 치어로 판단되는 음향 산란체가 주간시기 상층부터 해저면 부근까지 전층에 걸쳐 관측 되었다(Fig. 2). 이러한 요인은 유영능력이 약한 멸치 치어 시기 연안 조석류에 의해 표층으로 부유한 것으로 판단되고, 산란 초기에는 표층에 머무르고, 성장에 따라 저층으로 이동하는 멸치의 생태학적 특징과도 일치하는 경향을 보였다(Cha et al., 2013).
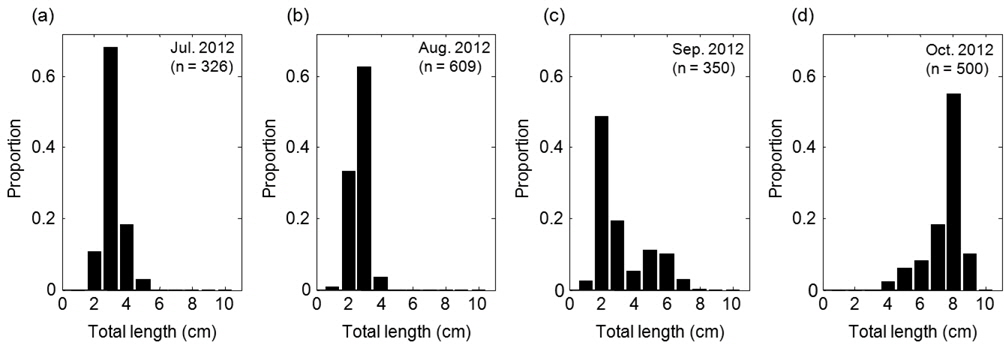
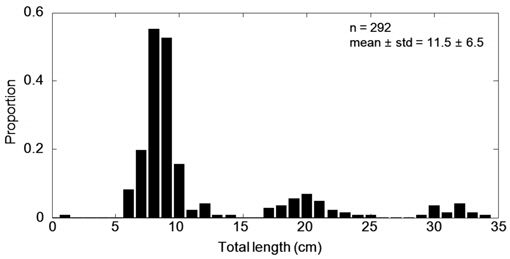
음향 조사를 통해서 측정된 멸치의 NASC는 10월>8월>9월>7월 순으 로 분포하였고, 가장 강한 음향 산란강도는 추계시기인 10월에 관측되었다(Fig. 5). 하지만 멸치 현존량 추정시 동일한 멸치 체장 조성이라면 현존량 또한 동일한 순서로 분포하겠지만, 7‒9월의 멸치는 치어 수준의 체장 분포를 보인 반면에 10월에 채집된 멸치는 성어 크기의 체장 분포를 보였다 (Fig. 3). 멸치 치어에 비해서 상대적으로 멸치 성어의 높은 음향 반사강도로 인하여 10월이 7−9월 비해서 낮은 멸치 밀도 및 현존량이 추정되었다(Table 3; KIOST, 2014).
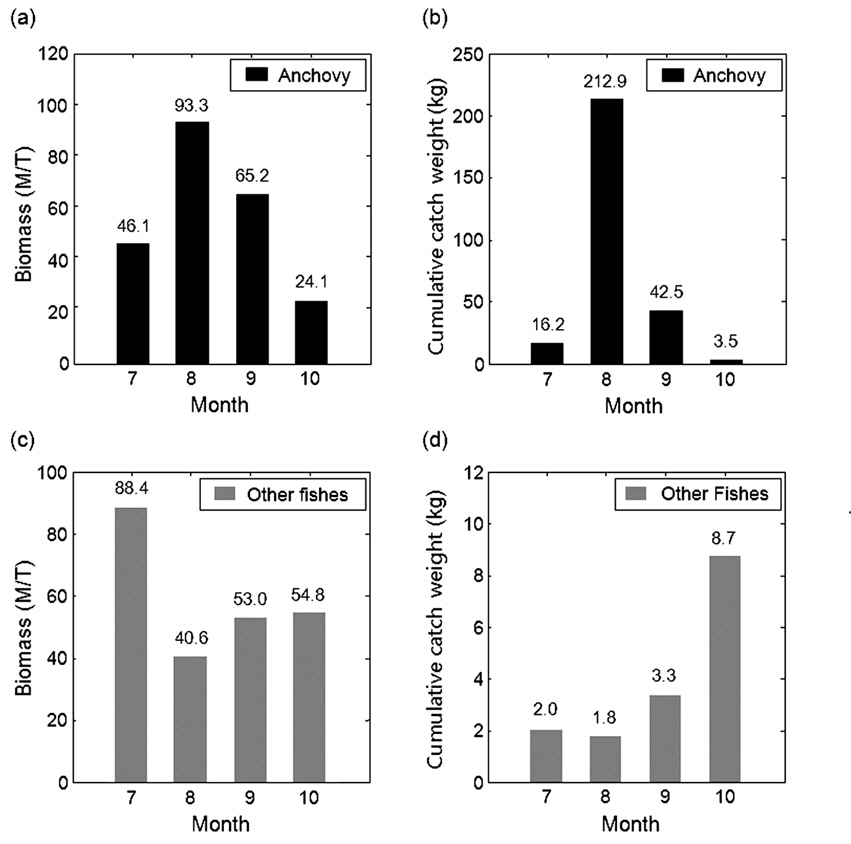
조사해역의 음향 자료를 통한 월별 현존량와 채집 자료를 통한 총 어획량의 무게로부터 멸치와 기타 어종의 하계 및 추계시기 분포 경향을 비교하였다(Fig. 7). 조사해역의 멸치 현존량은 8월이 최대, 10월이 최소였고, 채집 자료와 비교시 월별 경향이 동일하게 나타났다(Fig. 7a, 7b). 이는 유영능력이 약한 소형 멸치의 경우 채집 어구에 대한 회피성이 대형 어류에 비해 약해 포획될 가능성이 크기 때문에 음향 결과와의 편차가 적은 것으로 판단된다. 멸치에 대한 월별 분포 자료의 연관성을 알아보기 위하여 음향 밀도와 채집 멸치의 어획자료를 피어슨 상관계수(Pearson correlation coefficient)를 이용하여 상관분석을 실시하였다. 이때 비교군이 적어
멸치와 기타 어류를 다중 주파수를 이용한 신호 처리 기법(△MVBS)을 활용하여 현존량을 계산시 수층 내에 존재하는 동물플랑크톤과 반원니꼴뚜기(
본 연구에서는 아산만 인근해역에서 2012년 하계‒추계 시기 4회에 걸친 음향 조사를 통하여 주요 어류의 시·공간 분포 특성을 파악하였다. 기존의 어구 어법을 이용한 시·공간 분포 조사가 가지고 있는 어려움을 극복하기 위한 음향 조사 기법의 적용을 통해서 직접적인 분포를 확인하였고, 우점 어류에 대한 현존량 추정까지 가능함을 확인하였다. 본 접근 방법은 우리나라 연안 어류의 계절 더 나아가 연 변동성에 대한 다른 접근법을 제공 할 것이다.