우리나라는 세계 상위권의 패류생산국으로 2011년 패류생산량은 약 461천 톤으로 총 수산물 생산량의 14.3%를 차지하고 있으며 품종 별로는 굴이 306천 톤으로 66%, 바지락이 38천 톤으로 8.1%를 차지하고 있다(MOF, 2014).
패류는 생식, 가열조리 등 다양한 형태로 소비되는 국민기호 식품이지만, 생물학적 특성상 원료 수확해역이 연안에 위치하고 있고 여과섭식의 생리적 특성상 여러 가지 식품위생안전 위해에 노출될 가능성이 타 수산물에 비하여 상대적으로 높다(Hunter et al., 1999; Mallin et al., 2001). 특히 인구의 증가와 도시의 발달로 전염병이나 세균성 식중독 등 감염증을 유발하는 병원균이 함유되어 있을 확률이 높은 분변성 하수의 연안환경을 오염시키고 있으며 어장 및 양식장에도 심각한 피해를 야기시키고 있어(Metcalf, 1978; Miossec et al., 1988; Lipp et al., 2001) 생산단계에서부터의 철저한 위생관리가 요구되고 있다. 특히 굴의 경우는 가열 조리하지 않고 날것으로 섭취하는 경우가 많기 때문에 생산단계에서 오염되는 여러 가지 식품위생 위험요소로 인해 인체에 직접적인 위해를 미칠 수 있다(Sobsey and Jaykus, 1991).
수산물 섭취와 관련된 세균성 식중독의 가장 중요한 원인균은 장염비브리오균(Vibrio parahaemolyticus)으로 이 균은 전세계적으로 기수지역이나 연안의 해수와 수산물에서 널리 검출되는 해양상재세균이다( Ortigosa et al., 1994; Diggles et al., 2000; Urakawa et al., 2000). 이 세균의 식중독 관련성은 1950년 일본에서 최초로 밝혀졌고, 주로 하절기에 오염된 수산물의 생식이나 불충분한 가열 후 섭취 시에 발생하는 위장염의 주요원인이 되고 있으며 매년 하절기에는 이 균에 의한 식중독이 빈번히 발생하여 수산물 소비에도 큰 영향을 끼치고 있다. 우리나라의 경우 V. parahaemolyticus에 의한 식중독 발생 건수는 7월에서 11월까지가 80%이상을 차지하고 있다(KFDA, 2014). 따라서 이 균에 대한 효율적인 위생관리 및 식중독대책을 위해서 는 오염실태를 정확히 파악하고, 그에 대한 제어 등 적절한 대책의 강구가 절대적으로 필요하다.
한편 최근 임상 및 축산분야에서는 질병의 치료에 사용되는 각종 항균제에 내성을 나타내는 병원세균이 종종 분리되고 있으며, 수산분야에서도 어류양식장에서 장기간에 걸친 항균제의 사용으로 V. parahaemolyticus 등 병원성 세균군에서의 내성균의 발생이 우려되고 있다. 세균의 항균제 내성을 결정하는 인자들은 플라스미드 전이에 의해 쉽게 전달되는데 이러한 인자들이 사람에게 질병을 유발하는 세균에 전이될 수 있으며, 양식 생물이 이러한 매개체 역할을 할 수도 있다고 보고된 바 있다(Rhodes et al., 2000).
특히 항균제를 투여한 축산동물이나 사람의 분변으로부터 내성균이 자연환경으로 방출되기도 하는데, 사람이나 동물에게 투여한 항균제가 완전히 소화 흡수되지 않고 섭취된 항균제의 경우 약 70%정도가 성분이 변하지 않은 상태로 배설된다는 보고도 있다(Kümmerer, 2009).
따라서 본 연구에서는 우리나라 주요 연안 산 패류(굴, 바지락)을 대상으로 식중독 원인균인 V. parahaemolyticus 에 대한 오염실태 파악을 위하여 2011년 7월부터 11월까지 남해안 및 서해안 패류양식장을 대상으로 V. parahaemolyticus의 분포를 조사를 하였다. 또한 비브리오균의 항균제 내성패턴 연구의 기초자료로 활용하고자 분리된 균에 대한 항균제 내성 특성에 대해서 검토하였다.
V. parahaemolyticus 분리를 위하여 우리나라 패류의 주 생산지인 남해안(통영·거제) 및 서해안(곰소만) 패류 양식장에서 각각 굴(Crassostrea gigas)과 바지락(Venerupis philippinarum)을 2011년 7월부터 11월까지 매월 채취하여 시료로 하였다. 패류 시료는 멸균된 용기에 채취한 후 10℃ 이하로 유지하면서 실험실로 운반하여 실험에 사용하였다. 시험에 사용된 시료는 굴과 바지락이 각각 40개였다. 시료 채취 시 해수의 수온과 염분은 YSI 556 multiprobe system (Yellow Springs, YSI Life Science, OH, USA)을 사용하여 현장에서 측정하였다.
V. parahaemolyticus 는 Bacteriological Analytical Manual(Elliot et al., 2005)에 준하여 3 tube MPN법으로 정량분석을 실시하였다. 각각 8개 채취지점에서 채취한 굴과 바지락 시료는 채취지점 별로 12개체 이상을 1회 시험에 사용하였으며, 패각을 제거한 패육과 패액 200 g에 phosphate buffered saline (PBS: 140 mM NaCl, 5 mM anhydrous Na2HPO4, and 1.5 mM KH2PO4 pH 7.4)을 2배 비율로 첨가하여 균질화 하였다. 그리고 80 mL의 PBS에 균질액 20 g을 첨가하여 10배 희석액을 만들어 시료로 하였다. 희석액을 이용하여 100배, 1,000배 등 단계별 희석용액을 만들어 사용하였다. 각 희석단계 별로 alkaline peptone water (pH 8.5±0.2, 2% NaCl 함유)가 들어있는 3개의 시험관에 접종하고 35±0.5℃에서 18-24시간 배양하였다. 배양한 후 Thiosulfate Citrate Bile Salt Agar (TCBS agar, Difco)에 획선 도말하여, 35℃에서 18-24시간 배양하였다. 전형적인 청록색 집락을 도말하여 Triple Sugar Iron Agar(Difco)에 접종하고 35℃에서 24시간 배양한 후, 전형적인 반응을 나타내는 균주를 대상으로 동정시험을 실시하였다. 동정시험에는 내염성 시험, 42℃ 발육시험, ONPG (O-nitrophenyl-β-D-galacto-pyranoside)시험 등 생화학시험이 이용되었으며 VITEK system (BioMerieux Vitek, France)으로 확정하였다. 정량분석의 결과는 최확수표를 이용하여 100 g 당 MPN으로 나타내었다.
동정된 V. parahaemolyticus 균주를 대상으로 식중독을 유발시키는 독소 유전자인 tdh, trh 유전자의 존재유무 확인은 PCR기법을 이용하였다. 독소 유전자 tdh, trh 검출을 위한 primer는 각각 VPD-1/VPD-2와 VPR-1/VPR-2 (Takara, Japan)를 사용하였다. 시험균주를 Tryptic Soy Broth (Difco, USA) 10 mL에 접종하여 37℃, 18-24시간 배양한 다음 10분간 heating block(EYELA, MG-1200, Japan)에서 가열한 후 균액 5 µL, deoxynucleoside triphosphate mixture (1 mM dATP, 1 mM dCTP, 1 mM dTTP, 1 mM dGTP) 2µL, 10×PCR 용액 5µL, primer(100 pmol/µL) 1 µL, Taq polymerase 1.25 U를 혼합한 후, 총 부피를 50 µL로 조정하고 Thermal Cycle (Takara, Japan)로 94℃에서 60초, 60℃에서 60초, 72℃에서 60초의 반응을 35회 반복하여 PCR을 실시하였다
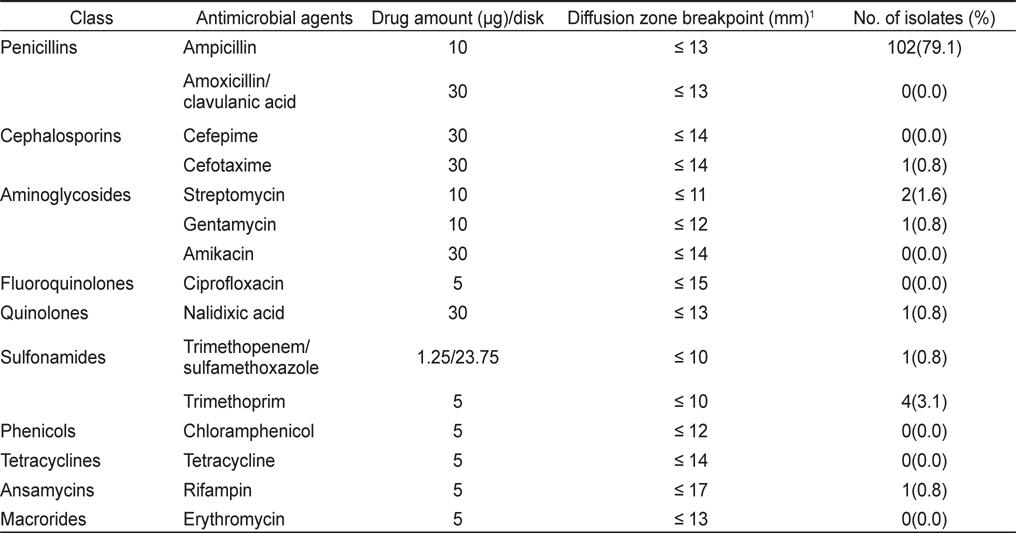
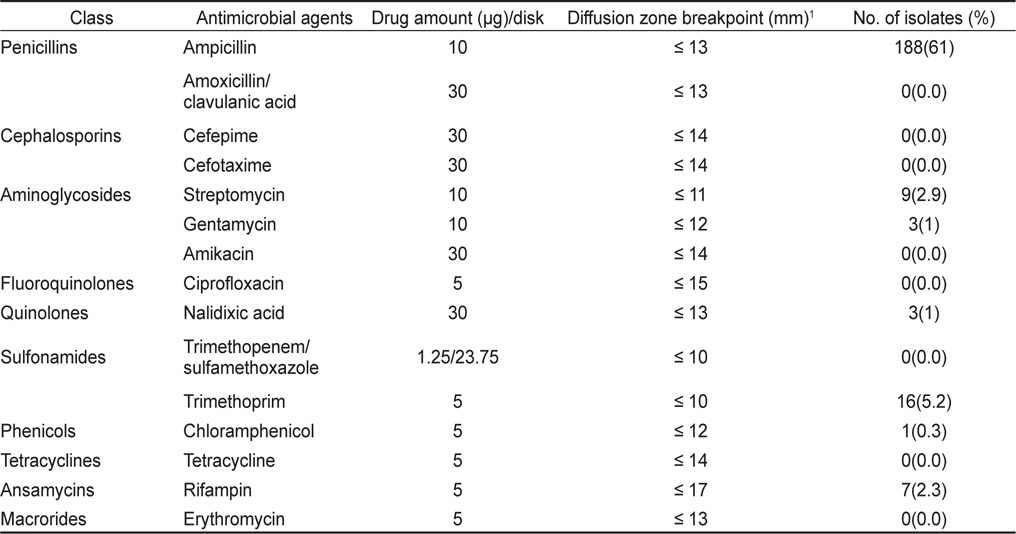
분리 동정된 각 V. parahaemolyticus 균주의 항균제 감수성은 Acar and Goldstein (1991)의 디스크 확산법을 이용하였으며 감수성 판정은 미국 NCCLS (National Committee for Clinical Laboratory Standards) 기준을 근거로 하였다. 분리된 각 균주를 Muller Hinton Broth (Merck, Germany)에 접종한 후 35℃에서 18-24시간 전 배양한 다음 균 배양액의 농도를 McFarland No. 0.5로 희석 조정하였다. 각 희석된 균 배양액을 미리 1% 농도가 되도록 NaCl을 첨가한 두께 4 mm의 Muller Hinton Agar (Merck, Germany) 평판에 도말한 후 실온에 5분간 방치하여 균액을 흡수시키고 항균제 디스크(φ 8 mm)를 균 접종 후 15분 이내에 고착시켰다. 시험 항균제는 Amikacin (30 µg), Ampicillin (10 µg), Amoxicillin/clavulanic acid (30 µg), Cefepime (30 µg), Cefotaxime (30 µg), Chloramphenicol (5 µg), Ciprofloxacin (5 µg), Erythromycin (5 µg), Gentamycin (10 µg), Nalidixic acid (30 µg), Rifampin (5 µg), Streptomycin(10 µg), Sulfamethoxazole/trimethoprim (25 µg), Tetracycline (5 µg), Ttrimethoprim (5 µg) 등 15종(BBL, USA)을 사용하였다. 항균제 디스크를 고착시킨 Muller Hinton Agar 평판은 35℃, 16-18시간 배양한 다음 균의 증식 저해대(inhibition zone)의 크기를 caliper로 측정하여 감수성 유무를 판별하였으며, 감수성 결과의 정도 관리를 위하여 Escherichia coli ATCC 25922를 대조 균주로 사용하여 각 항균제 디스크에 대한 역가를 확인하였다.
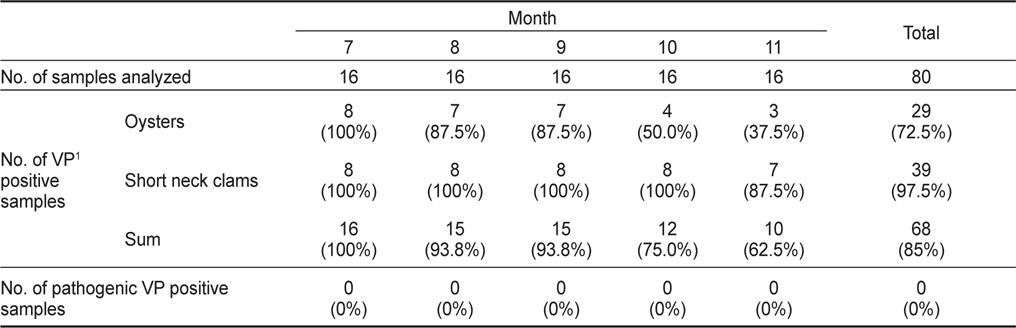
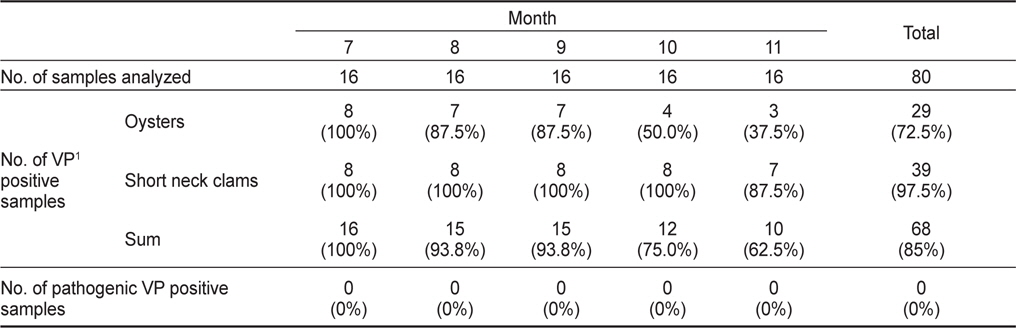
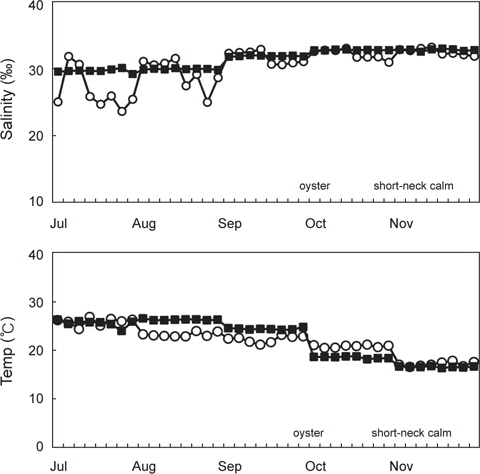
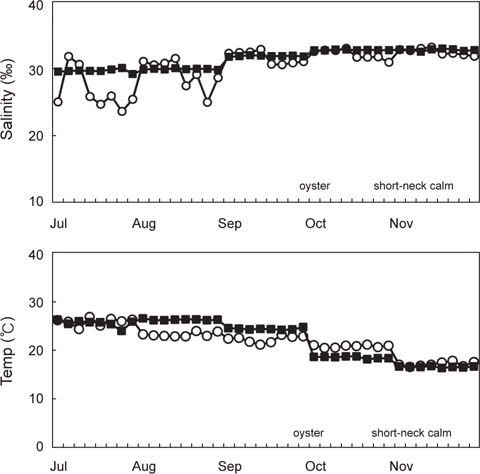
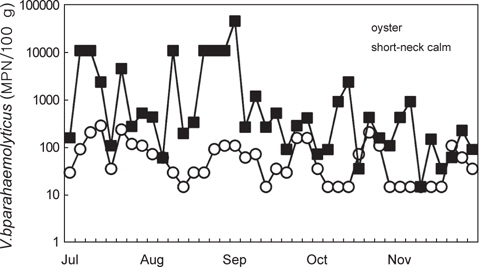
2011년 7월부터 11월까지 남해안 및 서해안 패류양식장에서 채취한 굴 40개 및 바지락 40개 총 80개 시료를 대상으로 V. parahaemolyticus 오염실태를 조사한 결과를 Table 1에 나타내었다. 조사 당시 수온의 범위는 16.4-26.9℃였으며, 염분의 범위는 23.6-33.1 psu이었다(Fig. 1). 굴과 바지락 채취해역의 수온 및 염분 차는 남서해안의 수리학적 특성과 해황에 기인하는 것으로 사료된다.
80개의 시료 중 68개의 시료에서 V. parahaemolyticus가 검출되어 85%의 높은 검출율을 나타내었으며, 특히 7월에는 모든시료에서 검출되었다. 비브리오균은 보통 염분농도가 낮은 기수지역에서 검출율이 높다고 알려져 있으나(Annie and Marta, 2012), 굴 양식장이 위치한 남해안에서 7월부터 9월까지 나타난 염분농도 변화는 동 시기 발생한 대량강우로 인한 일시적인 환경변화로 V. parahaemolyticus 검출율에는 큰 영향을 미치지 않았다.
굴의 경우 총 40개 시료 중 29개 시료에서 검출되어 72.5%의 검출율을 나타내었다. 시기별로 보면 굴은 7월에는 모든 시료에서 검출되어 검출율이 가장 높았으며, 수온이 낮아짐에 따라 검출율이 낮아져 11월에는 37.5% (3/8)를 나타내었다. 반면 바지락은 40개 시료 중 39개의 시료에서 검출되어 97.5%의 높은 검출율을 나타내었다. 7월부터 10월까지 모든 시료에서 검출되었으며 11월에도 87.5% (7/8)의 검출율을 나타내어 굴보다 높은 검출율을 나타내었다.
이러한 경향은 남해안산 굴은 연승 수하식으로 양식되어 해수중에 존재하는 V. parahaemolyticus만을 체내에 농축하는 반면 서해안산 바지락은 각종 세균이 다량 존재하는 펄에서 바닥식으로 양식되고 있어 환경에서 유래하는 V. parahaemolyticus를 체내에 고농도로 농축할 확률이 높은데 기인하는 것으로 보인다. Ju (1983)에 의하면 제주, 거제, 남해, 욕지, 부산 및 마산 지역 해수 및 펄에 대한 V. parahaemolyticus 분포 조사결과, 해수시료에서 V. parahaemolyticus 검출율이 9.8% (36/367)인 반면 펄 시료에서는 18.8% (59/313)에 이른다고 보고한 바 있다.
이 연구 결과는 Chao et al. (2009)와 Das et al. (2009)가 중국 및 인도의 어패류에서 V. parahaemolyticus 검출율을 각각 47.2%와 55%라고 보고한 것 보다 높은 검출율을 보이고 있다. Kirs et al. (2011)의 조사에 의하면 총 58개의 굴 시료 중 55개의 시료에서 V. parahaemolyticus가 검출되어 94.8%의 높은 검출율을 나타내었다고 보고한 바 있어 본 연구결과와 유사한 결과를 나타내었다. 또한 미국의 Chesapeake Bay에서 채취한 굴 시료에서 100%의 검출율을 나타내었으며(Parveen et al., 2008), 멕시코의 걸프만의 시료에서도 100%의 검출율을 나타내어(Zimmerman et al., 2007) 본 결과와 유사한 결과를 나타내었다. 이러한 결과는 수온이 상승하는 하절기에는 전 세계적으로 V. parahaemolyticus 검출율이 높은 것을 시사하고 있으며 V. parahaemolyticus에 의한 식중독의 발생 시기와도 일치하고 있다.
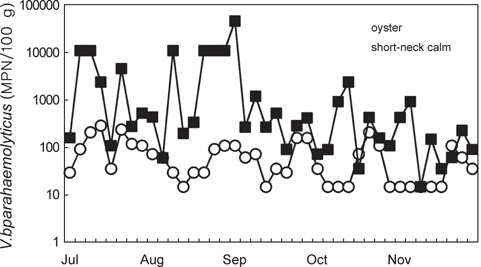
남해안에서 채취한 굴 중의 V. parahaemolyticus 균수는 <30-290 MPN/100 g였으며, 월별로는 7월과 8월에 높은 세균함량을 나타내었으며 11월에는 대부분의 시료에서 <30 MPN/100 g의 값을 나타내었다(Fig. 2). 굴의 V. parahaemolyticus 균수는 총 시료 40개 중 70% (28개 시료)가 <30-100 MPN/100 g의 범위를 나타내었으며, 101-1,000 MPN/100 g의 균수를 나타내는 시료는 30% (12개 시료)였다. Annie and Marta (2012)에 의한 Rhode Island의 굴과 해수에 대한 V. parahaemolyticus 조사결과, 7월에서 8월이 V. parahaemolyticus 에 의한 위험성이 가장 높은 시기라고 보고하고 있으며, Alabama 굴을 대상으로 시기별로 V. parahaemolyticus를 조사한 결과, 4월에서 11월에 100-1,000 CFU/100 g을 나타내었으며 12월에서 3월까지는 <100 CFU/100 g였다고 보고하고 있어 이 연구결과와 동일한 경향을 나타내었다(Angelo et al., 2003).
서해안 산 바지락 중의 경우 V. parahaemolyticus 균수는 <30-46,000 MPN/100 g이었으며, 7월부터 11월까지 채취한 총 40개의 시료 중 11월에 채취한 1개의 시료에서만 <30 MPN/100 g이었다. <30-100 MPN/100 g을 나타내는 시료가 총 40개 중 9개(22.5%), 101-1,000 MPN/100 g의 시료가 20개(50%), 1,001-10,000 MPN/100 g이 4개(10%), 10,000 MPN/100 g을 초과하는 시료는 7개(17.5%)이였다. 바지락 중 V. parahaemolyticus 균수는 101-1,000 MPN/100 g의 범위가 가장 많았으며, 10,000 MPN/100 g을 초과하여 검출되는 시료도 40개 시료 중 7개에 달하였다. 이중 57% (4개 시료)가 8월에 채취한 것으로 나타나 8월이 가장 많은 V. parahaemolyticus 식중독 환자가 발생한 것과 시기적으로 일치하는 경향을 보였다. 그러나 V. parahaemolyticus의 식중독을 일으키는 주요병원인자인 내열성 용혈독소(Thermostable direct hemolysin, TDH)와 내열성 용혈독소 유사독소(TDH-related hemolysin, TRH)의 유전자를 보유한 V. parahaemolyticus는 검출되지 않았다.
이상의 결과 패류 중 V. parahaemolyticus는 수온이 상승하는 하절기에 주로 검출되었나 병원성 인자를 보유한 V. parahaemolyticus는 검출되지 않아 수확 후 온도-시간 관리와 적절한 가공처리를 통하여 위생학적 안전성을 확보할 수 있을 것으로 사료되었다.
남해안산 굴과 서해안산 바지락에서 분리된 V. parahaemolyticus 436균주(굴 129균주, 바지락 307균주)를 대상으로 15종의 항균제에 대하여 감수성 시험결과를 Table 2 및 Table 3에 각각 나타내었다.
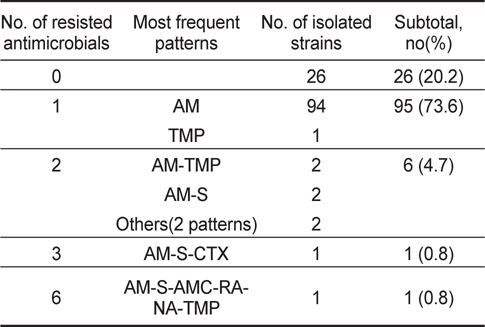
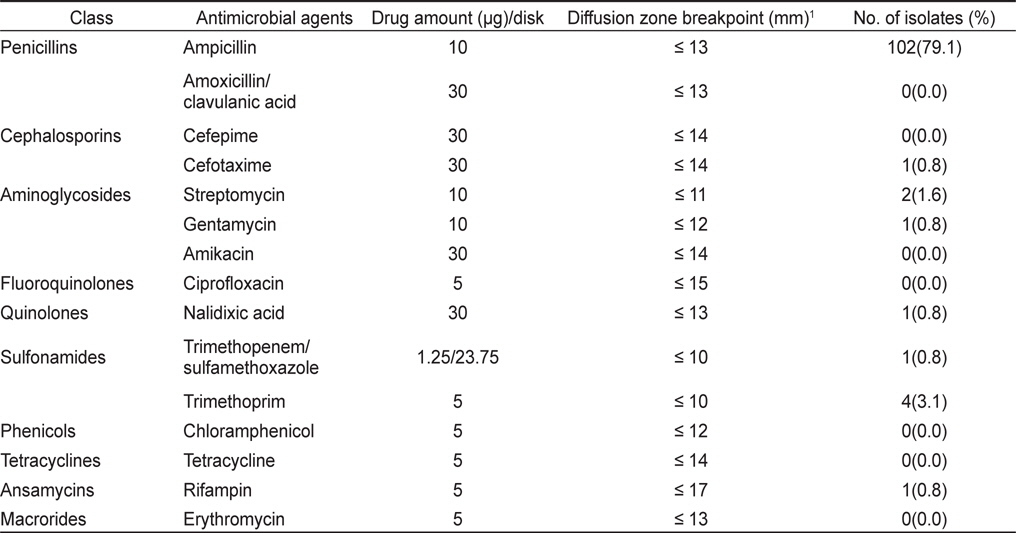
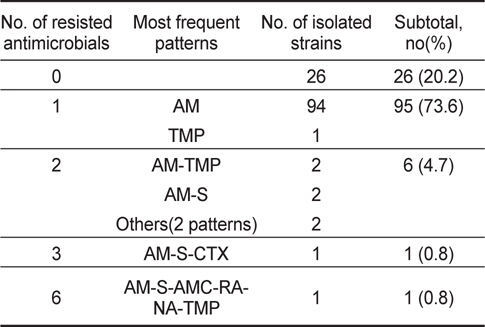
굴에서 분리된 V. parahaemolyticus 129균주 중 26균주(20.2%)가 항균제 15종에 감수성을 나타내었고, 나머지 103균주(79.8%)가 한가지 이상의 항균제에 대하여 내성을 나타내었다. 항균제 별 내성양상에 있어 제 1세대 항균제인 penicillin 계열 ampicillin에 대한 내성균 비율이 79%로 높게 나타내었으며 trimethoprim, streptomysin에 대해서는 각각 3.1% 및 1.6%의 내성균 비율을 보였다. 그 외의 항균제에 대해서는 내성균 비율이 1%미만으로 낮았다(Table 2).
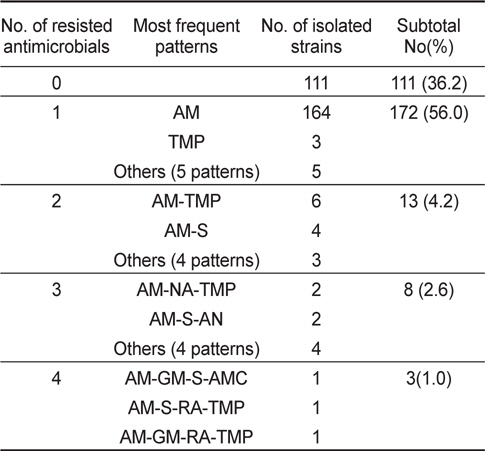
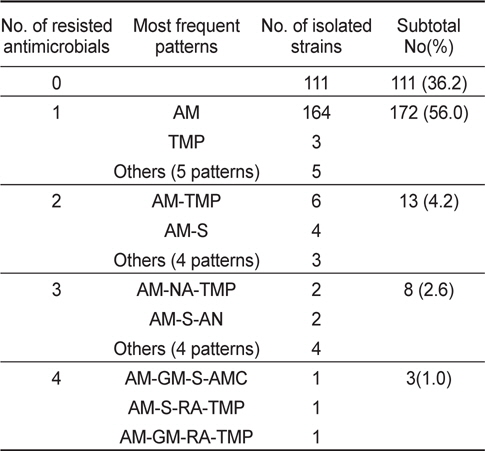
바지락에서 분리된 V. parahaemolyticus 307균주 중 111균주(36.2%)가 모든 항균제에 감수성을 나타내었고, 나머지 196균주(63.8%)가 한가지 이상의 항균제에 대하여 내성을 나타내었다. 바지락에서 분리된 V. parahaemolyticus 역시 ampicillin에 대하여 61.2%의 높은 내성균 비율을 보였다. 그 외에 trimethoprim, streptomysin, rifampin에 대해서 각각 5.2%, 2.9% 및 2.3%의 내성균 비율을 확인되었다(Table 3).
즉, 패류에서 분리된 V. parahaemolyticus는 ampicillin에 대해서 높은 내성을 나타내며 그 외의 항균제에 대해서는 대부분 감수성을 나타내어 내성패턴은 비교적 단순하였다. Oh et al. (2011)은 어류양식장에서 분리한 V. parahaemolyticus의 내성양상 조사결과, ampicillin에 57.8%, rifampin에 11.9%, trimethoprim에 6.4%, streptomysin에 8.7%의 내성균 비율이 확인 되었으며 그 외의 항생제에 대해서는 4%이하의 낮은 내성균 비율이 나타났다고 이 연구와 유사한 결과를 보고하였다.
많은 연구자들이 V. parahaemolyticus의 높은 ampicillin 내성에 관하여 보고하고 있다. Son et al. (2005)은 남해안 어류양식장의 어류와 해수에서 분리한 V. parahaemolyticus 균주 중 에서 97.9%가 ampicillin에 내성을 나타내었다고 보고한 바 있으며, 서해 연안해수(Lee et al., 2008)와 국내 유통 수산물(Lee et al., 2007)로부터 분리한 V. parahaemolyticus 균주 모두가 ampicillin에 대한 내성을 나타내었다고 보고된 바 있다. 또한 Oh et al. (2009)은 바다로 유입되는 기수지역에서 분리한 비브리오균에 대한 항생제 내성패턴 조사결과, V. parahaemolyticus 177균주 중 152균주(85.9%)가 ampicillin에 대하여 내성을 나타낸다고 보고하고 있다.
외국의 경우 인도네시아 Jakarta Bay에서 분리된 V. parahaemolyticus균주의 97%가 ampicillin에 내성을 나타내었다고 하며(Moritori et al., 1985), 이탈리아의 La Maddmlena, Tavolara Islands 그리고 Aleghero gulf 연안에서 분리된 Vibrio속 세균 중 80% 이상이 ampicillin에 내성을 나타내었다고 한다(Zanetti et al., 2001). 이와 같이 수산물뿐만 아니라 자연환경에서 분리된 V. parahaemolyticus을 비롯한 Vibrio속 세균이 ampicillin에 대하여 높은 내성을 가지는 것을 볼 때 ampicillin에 대한 Vibrio속 세균의 내성은 자연내성에 가까운 것으로 추정된다.
분리된 V. parahaemolyticus의 내성패턴을 Table 4와 Table 5에 나타내었다. 바지락에서 분리된 V. parahaemolyticus의 내성패턴이 22개 조합을 나타내어 굴에서 분리한 V. parahaemolyticus의 8개 조합보다 다소 복잡한 경향을 나타내었다. 그러나 대부분이 1개 항생제에 내성(굴-95균주; 73.6%, 바지락-172균주; 56.0%)을 나타내고 있고, 4가지 이상의 항생제에 대하여 내성을 나타내는 다제내성(Multiple antibiotic resistance)균은 굴에서 1균주(0.8%), 바지락에서 3균주(1.0%)로 매우 낮게 검출되었다. 뿐만 아니라 대부분이 ampicillin을 포함한 두 세가 지 항균제에만 내성을 보여 단순한 내성 경향을 가지고 있었다(Table 4, Table 5).
그러나 비브리오 균 중 일부는 내성인자들을 수계 침전물이나 주변 환경으로부터 획득할 수 있다는 보고(Neela et al., 2007)도 있으며, 주거지가 밀집하여 있는 지역에서 발생한 오수가 유입되는 지역에서 분리된 V. parahaemolyticus는 상대적으로 높은 내성균 검출 경향을 나타내었다는 보고도 있다(Oh et al., 2009). 따라서 다양한 내성인자를 보유한 세균들이 하천이나 마을 하수를 통하여 연안에 유입되어 해양에 존재하는 이종 또는 동종 세균으로 plasmid, transposon, integron 등과 같은 이동성 인자(mobile element)의 수평적 전이(horizontal transfer)를 통하여 내성유전자 풀이 형성될 가능성을 배제할 수 없다.
서식지가 연안에 위치하고 있으며 여과섭식의 생리적 특성을 가진 패류의 경우는 V. parahaemolyticus와 같은 식중독 원인균의 전달 매개체로서 나아가 해양세균과 육상세균 간 내성인자 전달 매개체로서도 위험 군에 속하는 수산물이기 때문에 생산단계와 수확 후 단계에서 적절한 위생학적 관리가 이루어져야 할 것으로 생각된다.